0022-538X/85/020607-09$02.00/0
CopyrightC 1985,American Society forMicrobiology
Isolation
of Novel
Herpes Simplex
Virus
Type
1Derivatives
with
Tandem
Duplications
of DNA
Sequences
Encoding
Immediate-Early
mRNA-5 and
an
Origin of Replication
KENICHI UMENEt* AND LYNN W. ENQUISTt
Laboratory of Molecular Virology, National Cancer Institute, Bethesda, Maryland20205
Received 25 June 1984/Accepted 24 October 1984
Twonaturally occurring variations ofherpes simplex virus type 1 (Pattonstrain) with novel tandem DNA
sequenceduplications in the Scomponentwereisolated,andtheDNAwascharacterized. These variantswere identified among a number ofplaque isolates by theappearance ofnew restriction enzyme fragments that
hybridized withradiolabeled DNA from theBamHI Zfragment (map coordinates 0.936to0.949) locatedin the
unique S region. Oneisolate,SP26-3, carrieda3.1-kilobase-pair duplication defined by recombination between asiteinthe BamHIZfragmentandasiteneartheorigin of replicationin theinvertedrepeatsequenceof the S component carried by the EcoRI H fragment. The other isolate, SP22-4, carried a 3.5-kilobase-pair duplication defined byarecombinationeventbetweenatandemrepeatarrayintheBamHI Zfragment anda sitenearthe amino terminus of the Vmwl75genein theS-region inverted repeatsequencecontained in the
EcoRIKfragment. Both duplicatedsegmentscontained the entireimmediateearly mRNA-5coding regionas wellastheorigin of replication located in the invertedrepeatsequenceof the Scomponent.TheDNAsequence
ofeachduplication jointwasdetermined.
Thegenomeof herpes simplex virus (HSV) is remarkable
foritsunusual structure. The DNA itself isalinear, double-stranded molecule ofca. 160 kilobase pairs(kb), consisting oftwocovalently linked components, Land S, that consti-tuteca. 82and 18% of thegenome,respectively (Fig. 1A). A
shortsequence, a, isrepeated directly atthe termini of the HSVgenomeandis alsopresentin the inverse orientationat the L-Sjunction (15). Theasequences areimplicated in the
isomerizationof thegenome,wherebythe L and Ssegments
of the HSVgenomeinvert, yielding four structural isomers
(1, 9-11).
A different kind ofstructural rearrangement of the HSV
genome occursafterserialpassageof HSVathigh multiplic-ities ofinfection. The virus preparations soobtained contain defectiveinterferingparticles. The DNA isolated from these particles consists of multiple head-to-tail reiterations of specific HSVsequences(3, 4, 6). The class Idefective DNA
of HSV type 1 (HSV-1) strain Patton can be formed by a hypothetical recombinational event between the terminal S-regionasequenceandasite withinthe BamHI Zfragment (the B5B6 fragment of reference 2) (Fig. 1).
Inthis report,we present another structuralalteration of HSV-1 DNA that occurred in our laboratory stocks. We describe the isolation and analysis of two HSV-1 (Patton) derivatives withuniqueduplications in the S component.
MATERIALS ANDMETHODS
Cellsandviruses. Vero cellsweregrowninminimal essen-tial medium supplemented with 5% calfserum. Stocks of HSV-1 strain Patton were prepared as follows. Virus from single plaques was used to infect Vero cells in minimal
*Corresponding author.
t Present address:DepartmentofVirology, FacultyofMedicine, Kyushu University, Fukuoka 812, Japan.
tPresentaddress:E.I.du PontdeNemours &Co., Inc.,Central
Research andDevelopment, Experimental Station, Wilmington, DE 19898.
essentialmedium with 2% fetal bovineserum.The
multiplic-ity of infectionwas 0.01 PFUper cell.
Single-plaque isolates from HSV-1 strain Patton used in
this workare summarized in Table1.
HSV-1 DNA was prepared from viral particles obtained
afterglycerol gradient centrifugation as described by Den-niston etal. (2).
Plasmids and phage. Bacteriological methods were as
described by Umene and Enquist (17). The Escherichia coli host for theexperiments was LE392. Hybrid plasmids used inthis workaregiven in Table 1, andtheywere grownand
purified as described by Denniston et al. (2). A hybrid lambda phage, Dec24, carries the EcoRI H fragment of HSV-1 strain Patton, and its DNA was extracted as de-scribedby Umene and Enquist(17).
Gelelectrophoresis and Southern hybridization. Restriction endonucleaseswerepurchased from New England Biolabs, Inc.,Beverly, Mass.,orfromBethesda Research Laborato-ries, Inc., Rockville, Md., and digestion conditions were those recommended by the manufacturers. DNA markers,
agarose and acrylamide gel procedures, Southern transfer, andhybridization conditions have been presented by Den-niston etal. (2).
DNA sequencing. The nucleotide sequences were
deter-minedasdescribed by Maxam and Gilbert (8).
RESULTS
Isolation ofHSV-1 strain Patton derivativescarrying dupli-cations ofsequences involving the BamHI Z fragment. We have previously reported that the BamHI Z fragment (B5B6 fragment) of HSV-1 strain Patton varies in size among independent single-virusplaque isolates (17). This heteroge-neitywasdue toadifference intandemrepeat numberofa 15-base-pair (bp)sequence (Fig. 1B and C) (17a). We exam-ined thefragmentpolymorphism of the BamHI Z fragment in more detail, and DNA from a number of single-plaque isolates of HSV-1 strain Patton was digested with BamHI 607
on November 10, 2019 by guest
http://jvi.asm.org/
A)
a TRL
B) 0
.# L
UL
5
--K S -KW
IRL a IRs USTRsa
10 15
_I kilobase pairs
B8
Us
three early mRNAs
B6 M4M313tM,2t1p MI Hp
B7B6Sall B?4 RIB3
11 a 11 1
D)
tanderm 3 4 f-
-repeat s IEmRNA-5
0 0. I
L I I kilobase pairs
B4 5 Ss5 S RI B3
I! ' II'_ I. v 1 tt
or
EmRNA-5
FIG. 1. Maps of HSV-1 DNA. (A) Structure ofthe HSV-1genome. HSV-1 DNA consists oftwocovalentlylinkedcomponentsdesignated L andS. Eachcomponentconsists ofuniquesequences(ULandUs)bracketedbyinverted repeatsequences(TRL, IRL,IRS,andTRs).The shortasequenceis noted. The EcoRIH and Kfragments aredesignatedH andK, respectively. (B) Mapofthe Sregionof HSV-1.BamHl sites(Bi toB8), EcoRI sites(RI), andaSallsiteareindicated.The BamHI Zfragment (B5B6)is indicatedbyahatched box.Sall-B5 region cloned intopBR322(pBS127-2) is indicated. (C) Map of theBamHI Z(B5B6) fragment. MboII sites(MltoM4),HpaIIsites(Hp),andaSall
site(S1)areshown. Asequenceoftandem 15-bprepeatsisindicatedbyasolid box(17a).The 3' end of IEmRNA-5 is indicated(19).Similarly,
the 3' endsof threeearly mRNAs transcribed from the oppositestrandareindicated(17a). (D) Mapof theBamHI X(B3B4) fragment.The EcoRI site(RI) and five SmaI sites(S)areindicated. The90-bp originofreplication (16)is indicatedbyasolid box. The 5' end ofIEmRNA-5 is indicated(19).
and electrophoresed on an agarose gel. After transfer to a nitrocellulose filter, the DNA fragments were hybridized witha32P-labeled BamHI Z probe (the SalI-BamHI subfrag-mentcloned inpBS127-2) (Fig. 1B,Table 1). Unexpectedly, in someviral DNA preparations, weobserved not only the
BamHI Z fragment on the autoradiography but also three otherfragments (Fig. 2). One isolate, SP26, had inaddition
toBamHI Ztwo extraBamHI fragmentsof 2.9 and 5.9 kb that hybridized with pBS127-2 (Fig. 2, lane 1). Another
isolate, SP22,hada single extraBamHI fragment of 1.4 kb
(Fig. 2, lane 6).
These anomolous BamHIfragmentsweremaintained
dur-ing passage of each virus stock. For example, four single plaquesof SP26 and sixsingle plaquesof SP22wereisolated, and the DNA of the secondary single-plaque isolates was
propagated and analyzed. All four SP26 secondary single-plaque isolates retained the two extraBamHI fragmentsof 2.9 and 5.9 kb. One isolate, SP26-3, was analyzed further. TheanomolousBamHIfragment fromSP22wasless stable. Outof sixsecondarysingle-plaque isolates, onlyone(SP22-4) retained the 1.4-kbBamHI fragment.
CompositionandoriginofnewBamHIfragments.Thenew BamHIfragments of SP26-3 and SP22-4werefirst cloned in
pBR322. The hybrid plasmids carrying the 5.9-, 2.9-, and 1.4-kb novel BamHI fragments were named pUK171, pUK172, and pUK174, respectively (Table 1).
The origin of these novel BamHI fragments was
deter-mined by labeling each hybrid plasmid with 32P and then
using these probes in Southern blot hybridization experi-ments with viral and cloned HSV-1 DNA fragments.
Ini-tially, we assumed that the new BamHI fragments were
likely tobe derived from the Sregion becausethey
hybrid-ized to a BamHI Z fragment probe. This assumption was
subsequently provencorrect, andsome of thehybridization
data that substantiated thisaredescribedbelow.
When the BamHI Z probe (pBS127-2) was labeled and hybridizedtoEcoRI-digestedviralDNA isolated fromSP26-3 andSP22-4,weexpected the EcoRI Hfragment (BamHI-Z is
contained within EcoRI-H) to be the only EcoRI fragment
visible. Instead, theprobe hybridized toadistinctive set of
fragments (Fig. 3). For SP26-3 DNA, the majority of the
fragments were longer than the EcoRI H fragment ofour standard HSV-1 isolate (SP23). In addition, the EcoRI
digests of SP22-4, but not SP26-3, revealed a new 3.5-kb
fragmentvisible after ethidium bromidestainingthat
hybrid-ized with the BamHI Z probe.
The simplest explanation for the appearance of the new BamHI and EcoRIfragmentsis thattheyresult from tandem duplications ofBamHI Z DNA sequences (Fig. 4). These conclusions were substantiated by detailed mapping ofthe DNA sequences contained by plasmids pUK171, pUK172,
andpUK174,bothbydirect restrictionenzymemappingand
B3 RI
IV
IRS
C)
B2Bl
I
0
pBS127- 2
TRs a
kilobase pairs
Bs B4
rI%T-r,-Tr, 'I
- -- o
IrlllllllllAI VI mw..o
2
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.156.464.69.379.2]by hybridization experiments with bacteriophage lambda
hybrids carryingHSV-1 EcoRI-H (Dec24).
Structures ofHSV-1genomeswith novelduplications.When DNA from stocks of SP22-4 and SP26-3 was analyzed, it
becameclear that theduplicationsasdrawn inFig.4 existed
in several permutations. For example, stocks of SP22-4 predominantly contained genomes with the same 3.5-kb
duplicationonbothsides of the Sregion (Fig. SA). Thiswas
mosteasilyseenby cleavingwith enzymes thathaveunique
sites in Us and in theduplicated sequence(Fig. 5B and6).
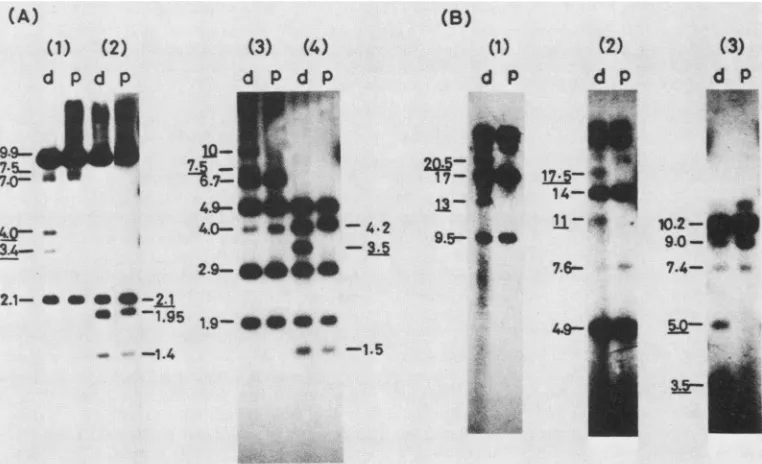
Forexample,XhoIcleavageof SP22-4 DNAgeneratednovel
fragments of4.0 and 3.4 kb that hybridized with a cloned
EcoRI Hprobe(Fig. 5B and 6). The 3.4-kbfragment derived from genomeswiththeduplicationontherightside of S, and the 4.0-kbfragment comesfrom genomes with the
duplica-tion on the left. Corroborating results were obtained with
otherenzymes(Fig.5Band 6).Itwaspossibletodeduce that
most ofthe genomes contained only duplications and not
triplications orhigher reiterations by cleaving SP22-4 DNA
with enzymes that do not cutintheduplicatedsequence. For example, when SP22-4 was cleaved withBglII and hybrid-ized with an EcoRI H probe, two novel fragments of 20.5 and 13 kb were observed; HindIII cleavage gave two frag-mentsof 11 and 17.5 kb(Fig.SBand 6). Fragments of these sizesare predicted only from duplicated sequences.
Ouranalysis of SP26-3 stocks indicates that the
duplica-tion exists in at least two permutations (Fig. 7A). One
population ofgenomes has the structure as diagrammed in
Fig, 4B. Itcontainedonecopyof theduplicationdefinedby the2.9-kb BamHI fragment(pUK172)on theright side ofS
plus one copy of the duplication defined by the 5.9-kb
BamHI fragment (pUK171) onthe left side ofS. The other
population contained twocopies oftheduplication defined
bythe 2.9-kb BamHIfragmentontherightsideofSplusone
copy of the same duplication adjacent to the duplication
TABLE 1. Virus,phage, and plasmids
Strain Description Reference
HSV-1strain Graham etal.
Patton (4)
SP22 Single-plaque isolate This report
SP23 Single-plaque isolate (used as a This report standard isolate)
SP24 Single-plaqueisolate Thisreport
SP25 Single-plaqueisolate Thisreport
SP26 Single-plaqueisolate This report
SP27 Single-plaque isolate This report
SP22-4 Single-plaque isolate from SP22 Thisreport
SP26-3 Single-plaque isolate from SP26 Thisreport
Phage
Dec24 Lambda hybrid phage carrying Umeneand
En-HSV-1 EcoRI H fragment quist(17)
Plasmids
pBS127-2 pBR322 derivative carrying Sall- Denniston et al.
BamHI(B5) fragmenton (2)
HSV-1EcoRI Hfragment
pUK171 pBR322 derivative carrying a Thisreport 5.9-kb newBamHI fragment
of SP26-3
pUK172 pBR322 derivative carrying a Thisreport 2.9-kb newBamHI fragment
ofSP26-3
pUK174 pBR322 derivative carrying a Thisreport 1.4-kb newBamHIfragment
ofSP22-4
l 2 3 4 5 6
5.9
2.9-
-aO * _ @*@
]BamHl
-Z
[image:3.612.359.522.76.256.2]* -1.4
FIG. 2. Southern hybridization profiles of BamHI digests of
single-plaque isolates of HSV-1 strain Patton, using 32P-labeled
pBS127-2as aprobe. Lanes: 1, SP26;2, SP27; 3, SP25; 4,SP24; 5, SP23; 6, SP22 (see Table 1). The sizes of the fragmentsareindicated in kb. TheBamHI Z(B6B5)fragmentwasdetectedin all isolatesas a fragment with heterogeneous length (1.8 to 2.0 kb) (17a).
Trhe
novel BamHI fragments in SP26 (lane 1) and SP22 (lane 6) are indicated.
defined by the 5.9-kb BamHI fragmenton the left side of S (Fig. 7A). Thiswas deducedbya digestion of SP26-3 DNA
with theenzymes describedinFig.7Band 8B. Forexample, in a Sacl plus EcoRI digestion, fourfragments of10.4, 7.7, 7.3, and 4.6 kb hybridizing to cloned EcoRI-H were ob-served (Fig. 8B, lane 3). These fragments can only be derived from the structures diagrammed in Fig. 7A, as the
predicted fragmentsfrom thestructure areshown inFig.7B.
FourextraEcoRI Hfragments were detected in the EcoRI digests of SP26-3 after hybridization with cloned EcoRI-H (Dec24) (Fig. 8A). The 18.5-kb EcoRI fragment can be derivedfrom the genome withonlyoneduplicationoneither sideof S(Fig. 7). The 25-kb EcoRI fragmentcanbe derived from thegenome with one duplication on one side of S plus
1 2 3 M
EcoRJ-H-
_
_
- 23.5
- 9.6
6.6
-4.45
[image:3.612.62.302.445.725.2]- 2.3 -1.95
FIG. 3. AnalysisofEcoRIfragmentsthathybridize to aBamHI Z probe. EcoRI-cut viral DNAs after electrophoresis on a 1%
agarose gelwere hybridized with32P-labeled pBS127-2DNAon a nitrocellulose filter. Lanes: 1, wild-type viral DNA (SP23); 2, SP26-3; 3, SP22-4; M, HindIll fragments of lambda phage DNA. The sizes of the fragments are indicated in kb. The EcoRI H
fragment is indicated.
on November 10, 2019 by guest
http://jvi.asm.org/
duplication
novelEcoRI fragment
8796 RI83 R183 8281
tandem t
repeats novel joint
83 95
pUK174
heterogeneous EcoRI-H fragment
duplication duplication
B) 8182 83RI
novel
joint
8e
as
pUKl71
~~P8I3 8281 07136 B584 ese4 BA1
11fmla" mlini
t novel joint
94 85
I I
pUKl72
FIG. 4. Structureof the duplications carried by SP22-4andSP26-3. (A)TheSP22-4 duplication. BamHI sites(Bi to.B8)and EcoRIsites (RI)areindicated. The solidboxesindicateinvertedrepeatsequencesof Scomponent.The boxeswithvertical linesindicate the 15-bp tandem repeatarrayfoundinBamHI-Z (Fig. 1C)(17a).Theduplicationisindicatedbyahorizontalarrowabovethe line. The novel EcoRI fragment isindicated. Thenovel BamHl fragment cloned inpUK174is shownbelow theline. Thenoveljoint is indicatedbyaverticalarrow.(B)The SP26-3 duplication. Thesymbolsarethe same asdescribedabove. Thetwo novel BamHIfragments cloned inpUK171 and pUK172are
indicatedbelowtheline.
two duplicationson the other side of S (Fig. 7). The other
EcoRI H fragments canbe explained asdrawn in Fig. 7.
FromSP26-3,twonovel BamHI fragmentswerecloned in
pUK171 and pUK172. The diagram of the SP26-3 duplica-tion in Fig. 7A adequately describes the twoBamHI frag-ments. The two novel BamHI fragments may derive from
the same duplication event (Fig. 9). The short duplicated segment of inverted repeatsequences of Scomponent
pro-videshomology for intramolecular inversiononthemolecule of line 1 of Fig. 9 (wavy line) as well as unequal intermo-lecularrecombination between the molecules of line 1 and line 2 ofFig. 9, either one of which will create the second novel BamHI fragment. The initial duplication could have created either the 2.9- or 5.9-kb BamHI fragment, and
subsequent recombination wouldyield the other. The initial recombinationaleventmight have been the essentialstepfor thegeneration of HSV-1genomewith theduplications. Once aduplicationwas made, it couldpropagate (Fig. 9).
Sequence analysisofthenoveljunctionsin thenewBamHI fragments. Thenucleotide sequenceof eachjunction in the newBamHI fragments wasdetermined and compared with corresponding known sequencesinthe HSV-1genome(Fig.
10). The nucleotide sequence across the novel joint of pUK171 and pUK172 was identical (Fig. 10). With the numbering system of Watson and Vande Woude (19), the EcoRI site in inverted repeat sequence of S component is used as the reference point. The noncoding strand with respect tothe immediate early mRNA-5 (IEmRNA-5)geneis presented. The novel joint found in the BamHI fragment cloned in pUK171 and pUK172 is defined by a fusion of
residue 79 (G) to residue 2891 (G) located in the MboII-2-MboII-3 fragment of BamHI-Z (Fig. 1C).
One segment of the noveljoint of pUK174 was defined
similarly, using the EcoRI site in invertedrepeatsequenceof
S component as the reference point. However, the other
segmentwasderivedfrom DNAencoding Vmwl75 (13). We
have used the numbering system of Murchie and McGeoch (13)todefine the site in Vmw175. Thenovel joint carried by pUK174 was formed by fusing 68 codons of the amino terminus of Vmwl75 (residue 1346, G) to the residue 2750 plus 20 of the second 15-bprepeatinthe BamHI Zfragment (Fig. 10D, E, and F).
DISCUSSION
We describe in thisreportthe isolation and characteriza-tion oftwo HSV-1 plaque isolates that contain DNA with novel duplications in the S region. Duplications of this general type have been described previously (5) among
recombinantsderived from markerrescueexperiments with Vmwl75temperature-sensitivemutations.Our observations relate tospontaneousduplications and the detailed analysis ofthe DNA structure.
The novel joint of the SP26-3 duplication fuses DNA between the IEmRNA-3 and IEmRNA-5 transcripts with
DNA in the BamHI Z fragment (Fig. 4B). The duplication jointislocatedbetweentwopromoters,one0.5 kb upstream
of the5'transcribedregion of the IEmRNA-5gene andone
0.2 kb upstream ofthe5'transcribedregionof the IEmRNA-3(Vmwl75)gene(13, 18, 19). This region alsoencompasses
anactivereplication origin (16). The DNA defining both the
A)
8on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.128.487.74.363.2]0 5
10
15
L I I Kb
e-EcDRI KS.EoRIH "
EcDoRl
K-,*(A)
S5,1 RI sc"Hki SsEH
B1S2 b3
sese RI 5i RI
SE SiCI k Rl ScSC
B7B6B5B4B3: B1
Rj
kx RI 9B1B2 B3BEt8B3
PJK174
B8 B7B6B5B4B3B5B4n II I 11 I 83 32Bn pUJK174
x
X -RI
Sc
Sc+
RI'-- 4.0
- 2.1
7.6
'- 4 3.5 10.3
Bg
20.5 ..H
K
411 17.5
[image:5.612.117.494.72.372.2]- - 5.0
FIG. 5. Analysisof the EcoRI Hregioninstocks ofSP22-4.(A)Restrictionmapsofunduplicatedviral DNA(top map)and thepredicted
structureofSP22-4DNA(bottom map).The BamHI sites(Bi toB8)areindicatedbelow each map.Saclsites(Sc),EcoRI sites(RI), Hindlll sites(H), XhoI sites (X), KpnIsites(K),andBgllI sites(Bg)areindicated. Theduplicated regionsareindicatedbyhatchedboxes,andextents
of duplications are noted by circles. Solid circles are the noveljunction. Open circles are the potential point involving the novel
recombination. The novel BamHIfragments clonedinpUK174areindicated.(B)Restrictionenzymecleavage productsthathybridizetoan
EcoRI H probe(Dec24)(Table1)aredelineated,and their sizes in kbareindicated,basedonthepredictedDNAstructure ofSP22-4shown
in(A).
(A)
(1) (2)
d P d P
(B)
(3) (4)
d P d P
(1)
d P
(2) (3)
4.2
-4
2-4Q
-
3.4-2.1-* _
:!:2f
1.95
1.a_
o MP v 4-1.4
d
Pd
P~w
-,14- _
li- 10.2
-
9.0-7.6-
7.4-V -1.5
FIG. 6. Southernhybridization profiles of restriction enzyme digestsofSP23 (parent;lane p) andSP22-4(duplication;lane d), usinga
32P-labeledEcoRI Hfragment (Dec24)as aprobe.The sizes of thefragmentsareshown inkb. The sizesofthefragmentswhichweredetected
onlyindigestsofSP22-4areunderlined.(A)Southernhybridization profilesofXhoI(lane1),XhoIand EcoRI(lane 2),Sacl(lane 3),andSacI and EcoRI(lane 4)digestsafterelectrophoresison a1%agarosegel. (B)Southernhybridization profilesofBglII (lane 1),HindlIl (lane2), andKpnI (lane 3) digests afterelectrophoresison a0.5%agarose gel.
(B)
34 - 4
2.1 '-4
3.5 1-4
13
3.5
'-v I II- 11 v -i
ri
I .
I I
I rzAo22zEzzzgr"- ri 11110,17wffirk
11 I I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.121.502.448.680.2]0 5 10 15 Kb -EcoRI-Kx EcoRI-H -*
EoRI-K-S,is
t s StI .a B7B6I II.
B3 B7B6B5B4:
Rj X.(
i84 B5 Bb pUKl71
RI
XK
KX KX RI B6 BONB 85B4
pUK172
K,^ KmX MRl
E85 E4B5 B8
I ie I
Bb B584 E B4 B5B4 :.-
i--i - 6.5
i -49.6
..-4 3.6
3.0 -i 3.6
- 4 4.6
i i 7.7
i i 4.7
, 7.8
17.6
, 4 4.6 3.1 i I 4 4. 6
6.5
-9.65 1
3.0 .-' 3.0 - - 3.0 7.3i . 4
10.4 F
7.1 i
-10.2 o
7.1
3.1 '-4
[image:6.612.143.471.73.456.2]3.1*- --4 3.1
FIG. 7. Analysisof the EcoRI Hregioninstocks of SP26-3.(A)Restrictionmapsofunduplicatedviral DNA(top map)andtwopredicted duplicationspresentinSP26-3DNA(middleand bottommaps).Thesymbolsaredescribed in thelegendtoFig.5.PvuIsites(P)andSallsites (S)areindicated. The novel BamHIfragmentscloned inpUK171andpUK172are represented. (B)Restrictionenzymecleavage products
thathybridizetoanEcoRI Hprobe (Dec24) (Table1)aredelineated,andtheir sizesin kbareindicated,basedonthepredictedDNAstructure
ofSP26-3 shown in(A).
originofreplicationand the transcribedregionof IEmRNA-5 isentirely includedin the duplication.
The noveljointof SP22-4 fuses DNAencodingtheamino terminus of Vmw175(68 codons;0.63 kb from the EcoRI site of the EcoRI Hfragment) tothe 15-bptandem repeatarray
located in the BamHI Z fragment (Fig. 4A) (13, 19). The
duplicatedsequenceof SP22-4 contains the total transcribed
region of IEmRNA-5 and a truncated transcribed region of IEmRNA-3. In addition, it also contains an apparently complete replication origin,asdefinedbyStowand McMon-agle (16).
Given the mechanisms togenerateduplications,onemust consider the capacity of the HSV DNA-packaging system. How much DNA can the capsid contain? An analysis of recombinants produced by rescue of HSV-1
temperature-sensitivemutantswith HSV-2DNAfragments indicated the
possibilityofaddingmorethan 8 kbtothe S component(5).
Our data indicate thateven more DNAcanbe added tothe
Scomponent. Wedetected fourextraEcoRI Hfragmentsin
SP26-3 viral DNA, the largest being28 kb (Fig. 8A). This indicates that HSV-1particles canbeformed withagenome
longer than wild type by at least 12.6 kb (28 to 15.4 kb;
normalEcoRI-H)ifotherpartsof the HSV-1genome arenot
deleted. There was no evidence of any larger EcoRI H
fragments. These apparently are not packaged or, if pack-aged, are not maintained as stableparticles. Perhaps larger duplications aremore readily lostduring replication due to homologous recombination events between duplicated
seg-ments. In fact, the molar quantity of the 28-kb EcoRI H fragment wasreduced, suggestingthat thegenome
contain-ingthe28-kb EcoRI Hfragment mightnothavebeen stable orthe DNAnot efficientlypackaged.
The DNA sequence of theregions involved in the
dupli-cations contains severalnoteworthy segments. One curious
observation isapotential DNAgyrasecleavage sequenceat the site of the SP26-3 duplication. DNA gyrase has been previously suggested to promote certain recombination
events in E. coli and eucaryotic cells (7). The consensus
sequence,5'-PyPu T GXPyX XPy-3' (Py,pyrimidine;Pu,
purine; X, random base), for an E. coli gyrase site was proposed by Morrison and Cozzarelli (12). The site of the
SP26-3 duplication in BamHI Z has the sequence
5'-TGCGGTCTT-3' (italicized Gisthe residue 2891/79in Fig. 10B),whichcorresponds to theconsensus sequenceexcept
(A)
=
(B)
RI J 18.5
I 21.6 ] 24.7 27.8
Sl+RI X+RI Sc+RI P+RI Bg+RI K +RI
R =7f4A7JM6YV70Jf-'
if 11 I I.
on November 10, 2019 by guest
http://jvi.asm.org/
(A)
(B)
(1)
(2)
(3)
(4)
(5)
(6)
P d d P d P d P d P d p d P
'1
Yk5'
- D6.04 7.1
4.2
04100
4.6-034_ *o 3.6-
044
3.0- *
3-1.915- w * 1
9-*-14.5-
=1.8- MD1.5
-,0.9~
0.8=
0.6-FIG. 8. Southern hybridization profiles ofrestriction c-nzymedigests ofSP23 (parent; lane p) and SP26-3 (duplication; lane d), usinga 32P-labeledEcoRIHfragment(Dec24) as a probe. The sizes of the fragments are showninkb. The sizes of the fragmentswhich were detected only in digests of SP26-3areunderlined. (A) Southern hybridizationprofiles ofEcoRIdigests afterelectrophoresison a0.5% agarose gel. (B)
Southernhybridization profiles of Sall andEcoRI (lane 1), XhoIandEcoRI(lane 2), Sacl andEcoRI (lane 3),PvuIandEcoRI (lane 4),BgIII
andEcoRI(lane 5), andKpnIandEcoRI(lane 6)digests after electrophoresis on a1%agarosegel.
for the third C residue. The significance of the strong boththeEcoRI-SmaI andMboII-2-MboII-3 fragmentsofthe
homology toan E. coligyrase cleavage site is notobvious. parentvirus(Fig.10Aand C). The sequence isnot present in
Analysis ofthe DNA sequences also implied that short the novel fragment ofthe SP26-3duplication (Fig. 10B).
DNA homologies may be involved (7). For example, the The viral DNA from stockscontainingthese duplications sequence5'-CGG-3' is found at the site of fusion of SP26-3 in contains nonduplicated DNA as well as additional tandem
Int
rd
molecular
pUK
721)
83
98
B786B51K
B5
RlB
12812
Intermolecular
3)
8485 8687I I
pLJ
Kl71FIG. 9. Modelforthederivation ofthe novelBamHIfragmentsinSP26-3. The symbols are described in the legend to Fig. 4. We have assumed that the initialduplicationcreated the2.9-kbnovelBamHIfragment (line1)defined by pUK172. The duplicated genome can undergo anintramolecular recombinationevent,usingTRshomology as indicated to generate the 5.9-kbBamHIfragment (line 3)defined by pUK171
(intramolecularpathway). Alternatively, the duplicatedgenome (line 1) could recombine with a normal genome in the inverted S permutation
(line2) to yield the same duplication (line 3) (intramolecular pathway).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.123.506.74.298.2] [image:7.612.107.512.435.680.2]I.pUK171, pUK172
A. RI-Smial 5--GTTGGTCGAACCCCCGGCCCCGCCCATCCGCGCC---3'
I .
63 79 96
novel joint
B. M2-Smal
5'--ACGCGCGTGTCGATGCGCCCCGCCCATCCGCGCC---3
1- i
2875 2891/79 96
C. M2-M3 5---AC G CGCGTGTCGATGCGGTCTT TT AG CGGAGCAG---3
2875 2891 2908
11. pUK174
repeat-1 repeat-2D. M1-M2 5---T
TG(CCACTCCCCACCCAC)(CCACTCCCC ACCCAC)---(
2750 2750.20
)CC AAA ---3
2772
repeat-i
novel4point
E. Ml-TaqI 5'--T
TG(CCACTCCCCACCCAC)tCA
GCGTCCTCGCCGGCG TCGGTGC---32750 2750-20 1346 1367
F. AluI-TaqI 5---CGCGGCGAGACGGCGTCCCC GGCGTCCTCGCCGGCGTCGGTGC---3'
[image:8.612.98.515.77.337.2]1325 1346 1367
FIG. 10. Nucleotidesequenceanalysis ofthenovel duplication junctionsfound in SP22-4 andSP26-3.LinesA, B,and C define the novel
junction inSP26-3 (the2.9-and 5.9-kbBamHIfragments cloned in pUK172 andpUK171,respectively).Thenumberingsystemisdescribed
inthetextandin references 13and19. (A) Aportion of theEcoRI-SmaIsequence(Fig. 1D) is shown. The 5' C residue of the Smal site of
theEcoRI-SmaIsegmentisatthe225th residue(19). TheG residueatnumber 79 is indicated.(B) Sequenceof the noveljunctiondefinedby
anMboII-2(M2)-Smal fragment. (C)Aportionof the MboII-2(M2)-MboII-3 (M3)sequencefrom BamHI-Z(Fig.1C) is indicated. The DNA sequencespresentedarethose of thenoncodingstrand of the IEmRNA-5gene asdefinedbyWatson and Vande Woude(19).LinesD, E,and Fdefine the noveljunctionin SP22-4(the 1.4-kb BamHIfragmentcloned inpUK174).Thenumberingsystemis described in thetextandby
Umeneetal.(17a)for(D)and Murchie andMcGeoch (13)for(F). (D)Aportionof the15-bptandemrepeatsequencein BamHI-Z(Fig. 1C) is shown. The Aresidueatnumber 2750 plus20is indicated. (E)Sequenceof the noveljunctionis indicated.(F)Aportionof thesequence ofanAluI-TaqI fragment fromwithin theB2B3BamHI fragment (Fig. 1B).
repeatsandpermutations of thebasic duplication.
Opportun-ities aboundfor recombination and unequal crossingoverto
generate avariety of novel S-region sequences in stocks of these viruses. One interesting effect of the duplication in SP26-3 (Fig. 4B) is that each inverted repeat region of S is
increased by3.1 kb (the size of the duplication) (Fig. 7 and
8A).Theramifications of thisonL-S inversionand defective
particle formation remaintobe seen.
The biological functions of the duplications observed in SP26-3 and SP22-4 are currently unknown. The selec-tive effects of multiple origins, additional promoters, and IEmRNA-5 segments must be examined. Although not ex-amined in detail, there are noobvious effects of the
dupli-cations on viruspropagation. It isnot clear howfrequently
these duplications occur in a given virus stock. Theywere isolated without selection after screening only 30 single-plaque isolates of strainPatton. Similarly, it is not obvious howtheseduplications relatetothose described by Knipeet
al. (5). Perhaps this region is recombinogenic due to the
presenceofreplication origins,activepromoters,andsetsof repetitive DNA sequences. The region is rich with such
detail. For example, the IEmRNA-5 region has been ana-lyzed in detail by Rixon and McGeoch (14). They have
shown that thereareactually three mRNAsspecifiedwithin this region that encode three partially overlapping genes (Vmwl2, Vmw2l, and Vmw33). These genes specify three mRNAs withdistinct 5' terminibuta common 3' terminus, thelongest ofwhich is IEmRNA-5. They speculatethat the Vmw2l protein interacts with DNA. Further study is
re-quired to determine whether these genes are expressed in the duplications. Furthermore, the relationship, if any, of
duplicationformation and thewell-knownL-Sisomerization
reaction and theformation of defective interfering particles
remains tobe established.
ACKNOWLEDGMENT
We thank R. Watson for invaluable advice. G. Vande Woudewas
the labchief.
Themajorpartof this workwasfunded by intramural research funds from the National Cancer Institute. A minorpart was sup-ported by grants from the Ministry of Education, Science and Culture of Japan. The costs for publication of this work were
supported byagrant-in-aid from the Fukuoka CancerSociety.
LITERATURE CITED
1. Davison, A. J., and N. M. Wilkie. 1981. Nucleotidesequencesof thejointbetween the L and Ssegmentsofherpessimplexvirus
types1and 2. J. Gen. Virol. 55:315-331.
2. Denniston,K.J.,M.J. Madden, L. W.Enquist,andG. Vande Woude. 1981. Characterization of coliphage lambda hybrids carrying DNA fragments from herpes simplex virus type 1 defectiveinterfering particles. Gene 15:365-378.
3. Frenkel, N. 1981. Defectiveinterfering herpesviruses,p.91-120. In A.J.Nahmias, W. R.Dowdle,and R. S. Schinazi(ed.),The human herpesviruses-an interdisciplinary prospective.
Elsevier/North-Holland Publishing Co.,New York.
4. Graham, B. J., Z. Bengali, and G. F. Vande Woude. 1978. Physicalmapof theoriginof defective DNAinherpes simplex
virustype1 DNA. J. Virol. 25:878-887. repeat-n
on November 10, 2019 by guest
http://jvi.asm.org/
5. Knipe, D. M., W. T.Ruyechan,B.Roizman,andI. W. Hallibur-ton. 1978. Moleculargenetics of herpes simplex virus: demon-stration of regions of obligatory and nonobligatory identity within diploid regions of the genome bysequencereplacement andinsertion. Proc.Natl. Acad.Sci. U.S.A. 75:3896-3900.
6. Locker, H., and N. Frenkel. 1979. Structure and origin of defective genomes contained in serially passaged herpes sim-plex virustype 1(Justin).J. Virol.29:1065-1077.
7. Marvo, S. L.,S.R.King, andS.R.Jaskunas.1983. Roleof short regions of homology in intermolecular illegitimate recombina-tionevents. Proc. Natl.Acad. Sci. U.S.A. 80:2452-2456. 8. Maxam, A. M., and W. Gilbert. 1980. Sequencing end-labeled
DNAwithbase-specific chemical cleavages. Methods Enzymol. 65:499-560.
9. Mocarski, E. S.,and B. Roizman. 1981. Site-specific inversion
sequence of the herpes simplex virus genome: domain and structural features.Proc.Natl. Acad. Sci. U.S.A. 78:7047-7051.
10. Mocarski, E. S., and B.Roizman.1982.Herpesvirus-dependent amplification and inversion of cell-associated viral thymidine kinasegeneflanked by viralasequencesand linkedto anorigin of viral DNA replication. Proc. Natl. Acad. Sci. U.S.A. 79:5626-5630.
11. Mocarski, E.S., and B. Roizman.1982. Structure and role ofthe
herpes simplex virusDNAtermini in inversion, circularization andgeneration of virion DNA. Cell 31:89-97.
12. Morrison, A., and N. R. Cozzarelli. 1981. Contacts between
DNAgyraseandits binding siteon DNA:features ofsymmetry
and asymmetry revealed by protection from nucleases. Proc.
Natl. Acad. Sci. U.S.A. 78:1416-1420.
13. Murchie, M.-J., and D. J. McGeoch. 1982. DNA sequence
analysis ofanimmediate-earlygeneregion of theherpes simplex virustype 1 genome (mapcoordinates 0.950to0.978). J.Gen.
Virol. 62:1-15.
14. Rixon, F. J., and D. J.McGeoch. 1984. A3'co-terminal family ofmRNAsfrom the herpes simplex virustype 1 short region:
twooverlapping readingframes encode unrelated polypeptides
one of which has a highly reiterated amino acid sequence.
Nucleic AcidsRes. 12:2473-2487.
15. Roizman, B. 1979. The structure and isomerization of herpes simplex virusgenomes. Cell16:481-494.
16. Stow, N. D., and E. C. McMonagle. 1983. Characterization of
theTRs/IRsoriginofDNAreplication of herpes simplex virus
type 1. Virology130:427-438.
17. Umene, K., and L. W. Enquist. 1981. A deletion analysis of lambda hybrid phage carrying theUsregion of herpes simplex virustype 1 (Patton). I. Isolation of deletionderivatives and identification of chi-likessequences. Gene 13:251-268. 17a.Umene, K., R. J. Watson, and L. W. Enquist. 1984. Tandem
repeated DNA inanintergenic region ofHerpessimplexvirus type1. (Patton). Gene30:33-39.
18. Watson, R. J., M. Sullivan, and G. F. Vande Woude. 1981. Structures oftwo spliced herpes simplex virustype 1 immedi-ate-early mRNA's whichmap atthejunctions of the unique and reiterated regions of the virus DNA S component. J. Virol. 37:431 444.
19. Watson, R. J., and G. F. Vande Woude.1982. DNAsequenceof
animmediate-early gene(IE mRNA-5)of herpes simplex virus
type I. Nucleic AcidsRes. 10:979-991.