ELECTRON MICROSCOPY OF INFLUENZA VIRUS components can bevisualized (see Fig. 3, 4, and
5), so that considerable caution must be exercised in attempting to interpret structural changes in the nucleoprotein component during penetration into the cell. The diffuse external coat of the virus comprises the radial spikes observed in negatively stained preparations. The definition of these
pro-jections varies,depending on the thickness of the section and whether the particle is central to the plane of section. There is some variation in size ofthe viral particles as can be observed by com-paringFig. 1 and 2 with Fig. 3 and 4. It will be noted that small, discrete droplets occasionally are attached to the virus (Fig. 5) or the cyto-plasmicmembrane (Fig. 6 and 8).These may be theglobulesofprotein or lipoprotein commonly found in CAfluid.In Fig. 1,thevirus is in close proximity to the cell, whereas inFig.2 it appears attached to the bilamellar cell membrane, which seems tobeintact. In Fig. 3, thereseems tobea connecting bridge of material between the viral
envelopeand the cell membrane. InFig. 5, neither
the cell membrane nor the lower aspect ofthe
viral envelope canbedistinguished. It should be
emphasized at this
point
that the state of thecytoplasmic membrane at any given site is very difficult to determine with certainty because the surface of the cell followsanundulantcourse.As a general rule, however, the membrane seldom bends sharply so that there is a transition zone from areas in which the membrane is cross-sec-tioned and thus sharply definedtoareas where it lies obliquely and is thus illdefined
(see Fig.
2).
At sites ofpresumed
viralpenetration,
the cellmembrane wasrepeatedly foundtobe
indistinct,
andin suchinstances, the transition from
sharply
defined to diffuse structure was
usually
abrupt
(Fig. 5 and 12). In Fig. 6, theenvelope
of the virus appears tohavemerged
with thecell,
and the limiting membrane of the core isdisrupted,
suggesting that the
nucleoprotein
has obtained access tothecytoplasm
of the cell.These first six micrographs serve to illustrate the stages bywhich it is believed influenza virus attaches to the
cytoplasmic
membrane and the viralnucleic acidgainsentrytothe hostcell. After 15 min of incubationat 37C,
about85%l
of the virushad disappeared (whencompared
with the number ofparticlesseenat5min).
Thisproportion
increasedto95%c
after 30min, andatthe end of 1 hrvirtuallyall viralparticles
had vanished.Al-thoughthe
preparations
were warmedquickly
to 37C, therewas considerableasynchrony
evenby
the fifth minute; this became morestriking
with the passage of time.Accordingly,
reference will not be made inthe text toeach time interval indiscussing the
ensuing
micrographs,
which have been chosen toillustratedetails inthe process of viral penetration. (The reader is referred thelegends for information regarding the time in-tervals.)
Virus was often observed at some distance from the cell (Fig. 7). As mentioned in a preced-ing communication (5),it ispossible in thick sec-tions to see a layer,presumed to be mucoprotein, covering the surface of the entoderm. Since this layer is oflow density anddoes not pick up the uranyl or lead stains to any great extent, it is virtually invisible in the thin sections usually employed. The virus appears first to become at-tached to this layer and, in fixed specimens, seems to be suspended in space. Penetration to the sur-face of the cell proceeds rapidly. Initially, even though virus appears to be in contact with the cell, both layers of the cytoplasmic membrane remain clearly defined (Fig. 8). The site of con-tact between viral coat and cytoplasmic mem-brane broadens (Fig. 9), and fusion of the viral coatwith theexternal layerof the membrane may occur (Fig. 10). Next, there seems to be disinte-gration of the cytoplasmic membrane in proximity tothevirus (Fig. 11). With fusion of the virus to the cell and underlying dissolution of the
mem-brane, direct continuity between the viral core and the cytoplasm is established (Fig. 12).
Stages in rupture of the core with release of nucleoprotein were so rarely observed that even
after prolongedsearchonlyafewexamples could be found. It was concluded that entry of the nucleoprotein must occur rapidly and, hence, be rarely encountered in the usual type of prepara-tion. Assuming that the process of entry
re-semblesanenzymatic reaction, which is inhibited at4C, itseemed reasonabletosupposethat
cool-ingthemembranesfrom 37 to 4 Cmightslow up
the reaction and enable it to be visualized at various stages. Such proved to be the case, and micrographs wereobtainedwhich clearly seemed todemonstratethe process ofentry (Fig. 13-17).
Despitecareful study, the migration of individual nucleoprotein strands into the cytoplasm could not be followed, presumably because dispersion of the strands after their release from the viral coretakesplacequicklyeven atlow temperatures. The final stages indisappearance ofthe viruswere not easy to document because identification of
the viral particles became increasingly
difficult.
Figure 18 illustrates a typical example of what appearstobeapoorly definedviralparticle.Note thatthecytoplasmicmembrane is indistinct bothdirectly beneath and in the immediate vicinity of
theparticle.Astudyofserial sections showed that such particles were not eccentric to theplane of section since they were repeatedly found in only one section of a series.
In the case of herpes simplex virus, it was pointed out that occasionally the process
ap-peared to misfire, andthe
capsid
wasdivested of 927 VOL. 2, 1968on November 11, 2019 by guest
http://jvi.asm.org/
8
10
9
Vs.=
*..stt-s
A. S f4$,,, .._*$
*
I1I
1
2
½
[image:2.489.63.454.72.642.2]4', ...4X t g
FIG. 7. Virus adsorbedtoCAM for 1 hrat4 Candthenexposedto37 Cfor 45minfollowed
by
15minat4C. X 200,000.
FIG. 8-12. VirusadsorbedtoCAM for 1 hr at4 Candthen keptat37 C for 10min (Fig. 8-11) orfor 15
min (Fig.9 and 12). X 200,000.
928
I
4l
I
.!.Ir
b: ,r
'
on November 11, 2019 by guest
http://jvi.asm.org/
14
15'
16
.41~~~~4'
- . .
.~;'.:4t4~'~ ~
N~~~~~
S~~~~~~~~~~~~~~~~~~~~'
'-1"~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~g-4~~~~~~~~~~~~~~~~~~~~~~~~V
~~~~~~~~~4r~~~~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ M
FIo.
13 18.VirusadsorbedtoCAMfor 1 hr at 4 Cand then exposed to 37 Cfor 45min followedby 15 minat4C. X200,000.
on November 11, 2019 by guest
http://jvi.asm.org/
MORGAN AND ROSE
its envelope before a passageway into the cyto-plasm had been established (6). A similar event seems to occur with respect to influenza virus. Figures 19-23illustrate examples ofviral
disinte-gration under circumstances which would not appear conducive to penetration ofthe nucleo-protein into thecytoplasm. The linkage between
virusandcell, illustrated in Fig. 22, is reminiscent of theprotrusions associated withherpes simplex
virus (6).
Inrare instances, there appearedtobea break in the cytoplasmic membrane that permitted penetrationofaviral particlewith anintactcore and a discernible envelope. In Fig. 24, for ex-ample, the cytoplasmic membrane close to the
junction with anadjacent cell has parted, and a viral particle is observed within the cytoplasm.
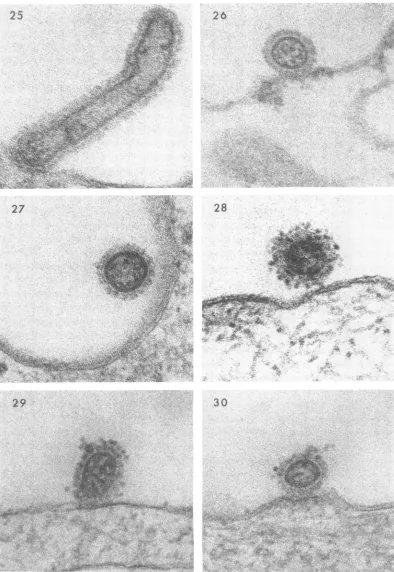
Another uncommon observation wasthe attach-ment of viral filaments to the cell surface (Fig. 25).Thescarcity of filamentous forms probably reflects their relative
instability.
Although virtually no virus remained on the
cell surfaceafter 1hrat37
C,
aninteresting excep-tion was the particle shown in Fig. 26. The cellhad undergone lysis so that very little remained
of thecytoplasm. Presumablythe irregular, rum-pled wall of the cell permitted attachment of the virus butpenetration could nottake place.
As noted earlier, the CAM exhibits relatively
littlephagocyticactivity, and,therefore, virus was rarely encountered within cytoplasmic vacuoles.
In most such instances, the virus
appeared
tobeintact (Fig. 27),which wasobtained froma
prep-aration incubated for 1 hr. The wall of this vacuole is poorly defined and probably oblique tothe plane ofsection. Viruswasneverobserved
at stages oftransit from avacuole into the
cyto-plasm. Itthus appeared, in contrast to the con-cept of penetration
by
phagocytosis,
that the virusactually may beprotectedand remainintactfor a considerable
length
oftime afterbeing
in-corporated in vacuoles.
Ina further effort to identify virus at terminal
stages ofentry,itwasdecidedtoadd ferritin-con-jugatedantibodytothepreparationwhen incuba-tionat 37 C was begun.It was hopedthat
virus,
suchastheparticle illustrated inFig.18, would be labeled and thus definitively identified. To our surprise, however, ferritin rarely attached to the virus once uncoating had begun. This suggested that during entry the antibody-combining sites atthesurface of the virus distaltothe cellbecome altered. The presence offerritin-conjugated
anti-bodyonthe surface of the virusdidnotinterfere
withattachment(Fig.28) but did prevent penetra-tion. Moderate numbers of particles
(consider-ably more than in preparations devoid of
anti-body) were adherent to the cytoplasmic
membrane after 1 hr ofincubation at 37 C. Note in Fig. 29 thatferritin is not evident between the cell surface and the virus particle, which probably means that tagging occurred after attachment had taken place. Nevertheless, the cell wall just beneath the virus, as well as the viral core, ap-pears to be intact. Ofparticularinterest was the observation that very few antibody molecules were necessary to prevent penetration. On the viral particle (Fig. 30), for example, only two ferritinparticles can be identified within the plane ofsection. Antibody did not prevent phagocyto-sis as illustrated by Fig. 31, which shows virus withinavacuole.
Oneconfusing type of structure often observed within the cytoplasm is shown in Fig. 32. At first, it was believed that intact virus had pene-trated the cytoplasm, where it wasuncoated with release of nucleoprotein (Fig. 33). Since such structures werealso present in uninfected cells, it soon became apparent that small phagocytic vesicleseither with (Fig. 34) or without (Fig. 35)
dense contentsformed at the free margins of the cell and migrated into the cytoplasm where, on occasion, they ruptured. The dark zone, so strongly suggestive of the viral coat in section, was commonly present on the surface of the vacuoleand apparently formed on the interior of the cytoplasmic membrane at the moment of budding inwards. From observations such as these, it can beinferredthatthethickening ofthe
cytoplasmic membranejust beneath the particle
(Fig. 36) indicates that phagocytosisof the virus was about to occur.
It seemed of interest to observe by negative stainingtheappearance ofvirusonthesurfaceof cells. Figure 37 shows virus after 1 minand Fig.
38 after 10 min ofincubation at 37C, following
adsorption at 4 C. It is evident in the latter
micrograph that the coat of the virus has been altered. Where the wall of the virus has not dis-integrated, the surface spikes are still visible, but they arepoorly defined and lack the orderly ar-rangement evident in the upper picture. Such changes are consistent with the suggestion made above that the antigenic sites of the viral coat at the timeof entry become alteredsothatantibody failstoattach. Theincreased density of the inte-rior of the virus (Fig. 38) maybe due to greater
permeability of thecoat tothe stain. DISCUSSION
Oneof themoststriking aspects of these obser-vations isthe rapidity with which uncoating and entryof the virus appearstooccur. Virtually all stages of the process were encountered within 5 min (Fig. 1-6), and there is reason to suppose
930 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
19
2
0
'''*
ri
en .:..
.sLs...
=;t s 8'SSfeS tNEi s i
s;n
.E>ot
R
r-n
fi X
=:
:>.
..!
iNN1 Ik.i..
* EN=. ;
:.s
,.',y
wit;.
orys22v+
*.
||
..xh
Sio
-e
.;-:;>t
tee
sPeSF
S;ri
;
\
:;
-F
*v
.
s
S
.t
s
.v
..Be%
;
^ Bj/_iE:e; .. .:^:r .. ..
<eg
... w. BISK.+'^t_, bi
.'a. eFS_ ^ ;hAiiti
,.
X
'
.'
-R.'&w
'
ti;
_' ", * 's
; '' z .' .4: ^S i.;_s _
2-3
S " 2 4 ., .' ..', '>%jA
iEV
'?'-
v-2
*.,
.t
t i X g S t R ;:
5111;
ail1E
I-*
i.;S
ite*iPi
FIG. 19-24. VirusadsorbedtoCAMfor 1 hrat4Cand then
kept
at37Cfor5min(Fig.
19-21)
orexposed
to37 Cfor 45 min(Fig.22-24)followed
by
15minat4C. X200,000.
-V
..
on November 11, 2019 by guest
http://jvi.asm.org/
26
I XP'..z"
28
;I
.a
4.
.I~~~~~~~~i..
30
FIG. 25. Virusadsorbedto CAMfor 1 hrat4C and then exposedto 37C for45minfollowed by 15min
at4C. X 200,000.
FIG. 26-27. Virus adsorbed to CAM for 1 hr at 4 C and then kept at 37 C for 1 hr. X200,000.
FIG. 28-30. Virus adsorbedtoCAM for 1 hrat4 C and then exposedto 37C for 1 hr. Ferritin-conjugated
antibodywasadded afteradsorptionat4 C. X200,000. 932
25
27
29
on November 11, 2019 by guest
http://jvi.asm.org/
E~~~
In~~
44
. F ; . -X < .. . e ;t Z'zr011
33...
34
.~.,.'Ai.4
antibody wa ade afe.dopto- t4C.2000
mm (Fig 36. 20,00
3
53
it-~t
;~~~~~~~~~~~~~~~~~~~~~~~~~
*e
FIG. 3 1. Virus adsorbedto CAM for 1 hrat 4 Candthenexposed to 37 Cfor I hr. Ferritin-conjugated antibodywasaddedafteradsorptionat4 C. X 200,000.
FIG. 32-36. Virus adsorbedtoCAMfor I hrat4C andthenkeptat37 Cfor 30 min (Fig.32-35) orfor 10 min(Fig. 36). X 200,000.
Aw
J711.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:7.489.34.431.50.629.2]MORGAN AND ROSE
37
38
* ' ;'c. NI'
"'S
FIG. 37-38. The surface ofanentodermalcell with attachedvirus.Negativestain.VirusadsorptiontoCAM wasfor1 hrat4C.Interval afteradsorptionwas at37 C for 1 min(Fig.37) orfor10min(Fig. 38). X 200,000.
934 J. VIROL.
4`.k
X&
on November 11, 2019 by guest
http://jvi.asm.org/
[image:8.489.60.454.35.607.2]ELECTRON MICROSCOPY OF INFLUENZA VIRUS that theuncoating of any givenparticle occurs in
evenless time. Ashasbeennoted, approximately 85%c of the virus had disappeared by 15 min. Considerable asynchrony was encountered so that a variety of stages were always observed in anypreparation. This mayreflect,in part atleast, thetime necessary for the virus to penetrate the surface coating of the cells (Fig. 7) and become closely attached to the cytoplasmic membrane (Fig. 8). In some cases, it would appear that a bond develops betweenthe envelopeofthevirus andthe outer layerofthe cell wall(Fig.9and10), thus providing a channel through which the nucleoprotein can enter the cytoplasm (Fig. 12). In other cases, the core is in close proximity to the cell (Fig. 6 and 11). The cytoplasmic mem-brane initially shows dissolutionat a small locus just beneath the virus (Fig. 11). This process
often extends so that at the moment ofentry a larger portion of the cell membrane becomes indistinct (Fig. 13-17). It can,ofcourse,be argued that in any given micrograph the membrane of
the cell may havebeencutobliquely and, forthis reason, is poorly defined. The micrographs cited
above, however, are representative of a large
number obtained over months of study, and it wouldseemunlikelythat ineachinstancethecell membrane inthe vicinity ofthe viruswasoblique totheplane of section. Theextensive dissolution of the cell membrane occasionally encountered could result from the attachment of a cluster of
viral particles (Fig. 37), only afew of which ap-pearwithinthe
plane
of section.Disintegration ofthecytoplasmic membranewas notaccompanied
by loss ofcytoplasmic components, and one can onlysurmise that attachmentofthe virus insome manner prevents outward movement of cell con-tents.Inaddition,repairofthecell membrane
ap-peared to be almost instantaneous after
disap-pearance of the virus. Cells, which must have received many viral particles, showed no visible alteration after the process of entry had been com-pleted. It isofinterest that therewasgreat varia-tion in the number of viral particles attached to anygivencell. Whethercells differ in the numbers which attach or intherapidity of entry, orboth, could not be determined, nor could the appear-anceof themitochondria, endoplasmic reticulum, ribosomes,ornucleus berelatedtotheamountof virus at the cell surface.
Ifthe stages ofpenetration are (i) attachment, (ii) dissolution of the viral coat, (iii) dissolution of thecytoplasmic membrane, (iv) ruptureof the core,and(v) entryofnucleoproteininto the
cyto-plasm,it isnotsurprising,consideringtherapidity
with which these events are occurring, that the precise sequence might occasionally go awry. Thus, dissolution of the viral coat and core might occur before firm attachment of the virus
or before an opening of the cytoplasmic mem-brane in close proximity to the virus is achieved, such as would seem to be the case in Fig. 19-23. Conversely, one can suppose that the cytoplasmic membrane could give way before dissolution of the viral coat, as may have occurred in Fig. 25. Examples of the former situation greatly out-numbered the latter.
Phagocytosis of virus was observed (Fig. 27),
but it was uncommon when compared to the large numbers of viral particles encountered in process of entry at the cell surface. It is note-worthy that very little phagocytosis of the virus had occurred by the tenth minute, at which time uncoatingand entry at the cell surface were most active. Moreover, stages in passage of the virus or of viral components through the wall of vacu-oles into thecytoplasm were notobserved.Indeed, the vacuoles actually appeared to protect virus from the events which had overtaken particles
attached to the cytoplasmic membrane. Such
observations leadustoconcludethat infection is initiated by entry at thecell surface rather than by phagocytosis.
It isfitting at this point to inquire why uncoat-ing and entry of the virus at the cell surface had previously escaped our attention, as wellas that
ofothers(1),who haveusedthethin-section tech-nique for studying virus replication. The answer undoubtedly lies in the rapidity with which the process occurs. If one examines cells that have
been incubated in the presence of virus, only a verysmallproportion of the particles is
encount-ered at adefinable stage of entry. In the present
study,twodifferent methodswereusedto
circum-vent this problem. First, some degree of
syn-chrony was obtained by
permitting
attachment to proceedinthecold and thenrapidly warmingthespecimens. Second,entrywasslowed downby
cooling themembranesafterincubation. Itwould not otherwise have been possible to dissect se-quential stages in the process.Inthe usualcourse ofexamination,after failingtoobserve entry,one is naturally inclined toexamine cells at progres-sively longer periods ofincubation. Since phago-cytosis ofvirus istaking placeat aslow but con-stant rate,and since, ashasbeen
mentioned,
the virus appears to remain within vacuoles for a considerable time, moreand more viralparticles
are observed within phagocytic vacuoles as the time of incubation islengthened.
Hence, it be-comes natural to conclude that it isby
this mechanism, rather than by therapid
and illusive processofuncoatingatthecellsurface,
that infec-tious nucleic acidobtains entrytothecytoplasm.
Itshould beemphasizedthatuncoating
of influ-enza virus atthe cell surface withpenetration
ofnucleoprotein is not a new concept. As
early
as1957, Hoyle and Finter (4), using virus labeled
VOL. 2. 1968 935
on November 11, 2019 by guest
http://jvi.asm.org/
MORGAN AND ROSE with radioactive isotopes, concluded "that on
entry into the cell the virus nucleoprotein is hydrolyzed with release of amino-acid, while the virus envelope protein and haemagglutinin re-main on the cell surface." More recently, Hoyle (3) again studied uncoating by examining in the electron microscope fragments of cytoplasm which had beenallowed to react with suspensions ofconcentrated influenza virus. He reported dis-integration of the virus lipoprotein layer, which "may result in rupture of the virus envelope with escape of the inner component." (Actually Fig. 4 in his paper resembles Fig. 38 of the present communication.) He makes it clear, however, that the eclipse phenomenon with disappearance of the virus does not occur in his system for it is "dependent on the active metabolic processes of the living cell." Presumably, he did observe at-tachment and theinitialstagesofviral uncoating,
although he was not able to follow events to completion.
Perhaps the mostperplexing problem which is raised by these observations is the precise mech-anism responsible for the uncoating process. If mediated by an enzyme, which would seem at first glance to be the most likely explanation, then the enzyme must be present in situ, for it couldhardlybesynthesizedinthe minute,or very fewminutes,necessaryforuncoatingof the virus to occur. But ifone assumes that some enzyme
actually attacks thecoatof thevirus, then how is it that veryfewantibody molecules, and these not
interposed between the virus and the cell, can prevent the action from occurring? Until
addi-tional information is obtained it would seem idle to speculate on these questions.
ACKNOWLEDGMENTS
WethankBaiba Mednis and FeM. Reyes for excel-lent technical assistance.
This investigation wasconducted under the spon-sorship of the Commission on Influenza, Armed ForcesEpidemiological Board, andwassupportedby the U.S. Army Medical Research and Development Command, Department of theArmy, under research contract no. DADA 17-67-C-7141, and by Public Health Service research grant AI-06814 from the NationalInstitute of Allergy andInfectious Diseases.
LITERATURE CITED
1. Dales, S., and P. W.Choppin. 1962. Attachment and penetration of influenza virus. Virology 18:489-493.
2. Duc-Nguyen, H., H. M. Rose, and C. Morgan. 1966. Anelectronmicroscopic study of changes at the surface of influenza-infected cells as revealedbyferritin-conjugatedantibodies. Virol-ogy 28:404-412.
3. Hoyle, L. 1962. The entry ofmyxoviruses into the cell. Cold Spring Harbor Symp. Quant. Biol. 27:113-121.
4. Hoyle, L., and N. B. Finter. 1957. Theuseof in-fluenza viruslabelledwithradiosulphurin studies ofearly stages of theinteractionof virus with the hostcell. J. Hyg.55:290-297.
5. Morgan, C., K. C. Hsu, and H. M. Rose. 1962. Structureanddevelopmentof virusesasobserved in the electron microscope. VII. Incomplete influenza virus. J. Exptl. Med. 116:553-564. 6. Morgan, C., H. M. Rose, and B. Mednis. 1968. Electron microscopy ofherpes simplex virus. I. Entry. J. Virol. 2:507-516.
936 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
JOURNAL OFVIROLOGY,Sept.1968,p. 937-943 Copyright(C 1968 AmericanSociety forMicrobiology
Vol.2, No.9
Prinitedin U.S.A.
Use
of
Zonal
Ultracentrifuge Systems
for
Biophysical
Studies
of
Rhinoviruses
JOHN L. GERIN,' WARD R. RICHTER,2 JAMES D. FENTERS,3 AND JACOB C. HOLPER
Departmentts of Ihifectious Diseaseantd Pathology,ScientificDivisioni, Abbott Laboratories, North Chlicago,Illinzois60064
Received forpublication29 May 1968
This paper reports the use of zonal ultracentrifuge techniques to conduct bio-physical studies of rhinoviruses grown with WI-38 cells. Good clean-out of
infec-tivityfrom rhinovirusharvestswasobtained with thecontinuous-flowB-VandB-IX
rotors. UseoftheB-Vrotor resulted in thesuccessful concentration of rhinovirus infectivity andantigenicity. Additional purification was achieved by the combined
use ofcontinuous-flow centrifugation and isopycnic banding procedures. Two
par-ticle sizes were found tobe associatedwith thevirus-infected cell harvests. The
in-fectious 22-nmparticlebanded indensity rangesof 1.38to 1.40g/cm3 in CsCland 1.26 to 1.27g/cm3 in potassium citrate. The 8.0 nm capsomere was composed of
2.0 nm subunits and banded with a density of protein at 1.28
g/cm3
in CsCl. Equivalent sedimentation coefficients of 155or185,dependingonparticledensityinsucrose,werecalculatedfromratezonalexperiments byuseof the B-IV zonalrotor. Rhinovirusesareassociated with upper
respira-tory illnesses in adults. They are classified as a subgroup of the picornaviruses and are
distin-guished from the enterovirus subgroup by their acidlability (16).
In view of the largenumber ofknown rhino-virus serotypes (11)andtheir poorgrowth intissue
culture,methods for theconcentration and
purifi-cation of rhinoviruses were considered for the purpose of
producing
concentrated polyvalent rhinovirus vaccines. Recentdevelopments
in zonal centrifugetechnology,
describedby
Ander-sonand his associatesatthe OakRidge
NationalLaboratories (1, 3), seemed applicable for this purpose. The use of these centrifuge systems for the biophysical characterization of rhinoviruses is reported in this paper.
METHODS AND MATERIALS
Rhinioviruses. The rhinovirus numbering system is that recommended by the Directors of the World Health Organization Respiratory and Enterovirus ReferenceCenter (15).Rhinovirus strains 1A, 1B, 2,
1Present address: Laboratory of Viral Diseases, National Institute ofAllergyandInfectious Diseases,
Building 7,Room 304,NationalInstitutes ofHealth, Bethesda, Md. 20014.
2Present address: University of Chicago, Depart-ment ofPathology, Chicago, 111. 60637.
3Present address: Illinois Institute of Technology, Research Institute,LifeSciences,ChemistryBuilding, 10 West 35thStreet,Chicago,111. 60616.
13, 14,and 17 were used in this study. Their history wasgiveninanearlierreport (10).
Tissue culture, virus growth, anid bioassays. The humanembryonic diploid WI-38 strainusedinthese studieswasobtained from Flow Laboratories, Rock-ville, Md.andfrom L. Hayflick oftheWistarInstitute inPhiladelphia,Pa. Themethodsusedforthepassage ofWIcellswere asdescribed by Hayflick and Moor-head(12). Conditions of cellgrowth weregiven inan earlier report (10).
WI-38 cell cultures were infected with approxi-mately 102TCID5 ofrhinovirus, and harvested when 80tolOO1 of the cells showed cytopathic effects(13). Harvestswere pooled and heldat -20 C. Infectivity titrations andserumneutralizationtestsweredonein tissue culture tubes of WI-38 cells and rolledat30 to 32 Caccordingtopublished procedures (10).
Preparationi ofguintea pig anitiserum7Z. Rhinovirus harvest or B-V concentrate was inijected into guinea pigsby either oftwoprotocols.Aftera preimmuniza-tion bleeding, agroup ofpigs was in-jected intraper-itoneally with2mlofaqueousvirustwiceaweek for 2weeks (protocol A), orintramuscularly withtwo 1-mlinjections ofaqueousvirus3 weeksapart(protocol B). All guinea pigs were bled 1 week after the last injection.
Con2tintuouis-flow
centtrifuigationt in the B- V rotor.Barringeretal. (3) havedescribedthe B-V rotor and its operation. Rhinovirus harvest material was pro-cessed in the B-Vrotor at 40,000rev/min at 5 Cat
various flow rates under aseptic conditions. After processing, the rotor was braked to a stop and the pelleted material was resuspended in the rotor con-tents. Pellet resuspension was aided by the use ofa
rubberpoliceman orby gentle shaking oftherotor at
on November 11, 2019 by guest
http://jvi.asm.org/
GERINET AL.
4 C overnight before unloading, or both. The sus-pendedpellet (130 ml) and the final rinse [20 ml of 0.01 M phosphate-buffered saline (PBS)] were then suctioned directlyintoa sterile vacuum bottle.
Combined continuous-flow centrifugation and iso-pycnicbanding inthe B-IXrotor. Andersonetal. (1) havedescribed the B-IX rotor anditstypical opera-tion. The B-IX rotor was loaded while spinning at 5,000rev/minwith eithera750-ml two-step (1.1and 1.4g/cm3) potassiumcitrate gradientor aprogrammed 600-ml linear (1.1 to 1.5 g/cm3) potassium citrate gradient followed by a 150-ml 1.5 g/cm3 potassium citrate cushion. The rotor was accelerated to speed and the harvest was processed through therotor at various flow rates. When the starting sample was expended, therotor was runfor 1 to 2 hrtoensure equilibriumandthenwasdeceleratedwithoutbraking to5,000rev/min. Thedensity gradientwasunloaded by pumping dense potassium citrate to the rotor peripheryand the resulting effluent was collected in thirty25-mi fractions.
Rateseparation in theB-IV rotor. The B-IVrotor (2) wasloaded at 5,000 rev/min withalinear 1,200-ml 10 to 55% (w/w) sucrosegradient followedbya 500-ml (w/w) sucrosecushion byuseofaBeckman (model 131) high capacity gradient pump. Sterile sucrose solutions were made by dissolving sucrose crystals in 0.01 M PBS (0.85%) and purified in our laboratory bypassageovergranularcharcoalcolumns. The rhinovirussample wasintroducedinto therotor in a25-ml volume followed by a 200-ml overlay of PBS,displacinganequivalentvolumeofcushion from the rotor periphery. The rotor was accelerated to
40,000rev/min,runfor 90 minat5C,anddecelerated without brakingto 5,000rev/min. The gradient was recovered from the rotorby displacement with
55%0
(w/w) sucrose solution and collected in 85 fractions of 20 ml each.Sedimentation coefficientswerecalculated(courtesy ofN. G. Anderson) bythe computerprogramatthe Oak Ridge National Laboratories. Equivalent sedi-mentation coefficients (S*; 4) were returned corre-spondingtothecenterof eachfraction forarangeof particle densities from 1.2to1.6g/cm3.
Isopycnic banding incesium chloride denisity gradi-ents. Rhinoviruses were banded in linear preformed cesium chloride gradients (1.1 to 1.6 g/cm3) in the SW25.1 rotor at25,000rev/minfor20 hr at 5C.By unloading thecentrifuge tubes from the bottom,the gradients wererecovered and collected in 2-ml frac-tions.
Denisity mneasurements. Potassium citrategradients wereeitherdetermined bythe useofstandard curves relating densityto refractive index or measured di-rectly withapycnometer.Refractive indiceswereread onthe Abbe 3Lrefractometer for the determination of cesium chloride densities and sucrose concentra-tions.
Protein. Samples ofrecovered gradient fractions were dialyzed extensively against PBS at 4 C and assayed for protein concentration by the method of Lowry et al. (17).
Electronz microscopy. Dialyzed samples of rhino-virus preparations were negatively stained with 2%
phosphotungstic acid (PTA) at pH 7.0 or with satu-ratedaqueousuranyl acetate by use of carbon coated collodion grids. Micrographs were taken with the RCA-EMU-3G microscope at initial magnifications of50,000 X or 100,000 X.
RESULTS
Concentration ofrhinovirusesbydirect pelleting in the B-Vrotor. Figure 1 shows the results of clean-out studies with rhinovirus IA in the B-V continuous-flow rotor. More than 95% of the infectivity wasremovedatflowrates up to 4liters perhr.Table 1 shows the concentration of rhino-virusinfectivity intheresuspended contents ofthe B-V rotor for a number ofseparateexperiments in which rhinovirus 1Aor 13harvests were used. Table 2 shows the concentration of rhinovirus antigenicity, asmeasured by guinea pigpotency tests, for several experiments inwhich either live orFormalin-inactivated harvests were used as starting material. Althoughcomplete recovery of infectivitywasnotalways obtained, good concen-tration ofantigenicity was achieved in all cases. An average 30-fold purification, on a protein basis, was also obtained by this direct pelleting procedure. We have concentrated up to 50 liters of membrane-filtered (0.45
Au,
Millipore Corp., Bedford, Mass.) rhinovirus harvest in a single B-V run under continuous operation.Examination of the lB-V concentrates by elec-tron microscopy revealed the presence of 22-nm virus like particles and numerous small annular
I-C.J L&. z U-cJ ,., I-. Lu 1-, 100 90 . 80 -70 -I
1 2 3 4 5 6 7 8
FLOW RATE(LITERS/HOUR)
FIG. 1. Threeexperiments with rhinovirus IA har-vestsshowingtheclean-out ofinfectivityat40,000revl min as afunction offlowrate intheB- Vrotor.
938 J. VIROL.
60,