Vol. 49, No. 2 JOURNALOFVIROLOGY, Feb. 1984,p. 479-489
0022-538X/84/020479-11$02.00/0
Copyright ©D 1984,American SocietyforMicrobiology
Two Types of Deletion Within Integrated Viral Sequences Mediate
Reversion of Simian Virus 40-Transformed Mouse Cells
KAZUO MARUYAMA* AND KINICHIRO ODA
Department ofTumor VirusResearch, Institlute of MedicalScience, University of Tokyo, 4-6-1, Shirokanedai, Minato-ku, Tokyo 108, Japan
Received 11 May 1983/Accepted 23 October 1983
Simian virus 40 (SV40) DNA insertions from SV40-transformed mouse cell line W-2K-11 and its
revertants M18, M31, and M42were cloned. W-2K-11 cells contain 1.5copiesof the SV40sequences ina
partially tandem duplicated form. The endpoints of the viral sequences at the virus-hostjunctions are located veryclosetothose reported by others, indicatingthat thereare somepreferred sites forintegration and rearrangement in SV40 sequences. One flanking cellular sequence is a long stretch of adenine and thyminewithrepeated AAAT, and the other isastretch ofguanineandcytosinewithrepeatedCCG. There
are patchy homologies between the flanking cellular sequences and the corresponding parental SV40
sequences. The sequencesaroundbothjunctions wereretained in all the revertants,whereas most ofthe
internal SV40sequencescoding for large T antigenweredeleted. Thecodingsequencesfor small T antigen areintact,and small Tantigenwasexpressedinall therevertants.Thefragments cloned from M18 and M42 wereidentical and 3.9 kilobases ofSV40sequences weredeleted. The parental SV40sequences aroundthe deletion site have sequences capable of forming a secondary structure which might reduce the effective
distance between thetworegions.TheSV40DNAretainedinM31iscolinear withSV40 virion DNA, anda unit lengthofSV40 DNAwasdeleted within the SV40sequences present inW-2K-11 cells. These results indicatedthattwo typesof deletion occurred during the reversion,onebetween
homologousl
sequences and theother between nonhomologous sequences.Ingeneral, theorganization ofacellular genome is stable and is conserved from
generation
togeneration.
However,the genomic structure has the potential to acquire new genetic information bygene rearrangements-an induction ofnewcovalent
linkage
betweendistant DNA sequences-such as amplification, deletion,transposition,
andacquisi-tion ofexogenous sequences. The interaction ofthe simian
virus 40 (SV40) genome with cellular genomes is a good
systemwithwhichtoanalyzegene rearrangement in
eucary-otic cells, since the covalent linkage between SV40 and
cellularDNAhasbeen foundintransformedcells(24)andin the evolutionary variants generated after serial passages of
SV40at high multiplicity of infection (15).
Characterization of
evolutionary
variantsby
restrictionenzyme analysis and electron microscopic heteroduplex mapping has suggested that recombination between SV40 and cellular DNA could occur at
multiple
sites (16,21).
Using Southernblotanalysis,
it has beendemonstrated that there is no specific site ofintegration
in both SV40 and cellular DNA in the transformed cells (6,13). By
means ofrecombinant DNA techniques, the fragments
containing
SV40 sequences from the transformed cells and those
con-taining the corresponding cellular sequences from
untrans-formed cellswere cloned to determine whether the
specific
sequences are involved in the
integrative
recombination.The results have also shown that there is no evidence of homology between viral and cellular DNAsattheintegration
sites (5, 23). However, Stringer reported an exception in
which there are five base pairs shared at the junction
between SV40 and rat DNAin the SVRE9 cell line (27).
Thegenomic structure seems tobe unstableatthe site of insertion of viral sequences (7, 12). We previously isolated
revertantsfromSV40-transformedmouseandratcellswhich
*Correspondingauthor.
contained multiple copies of viral DNA serially arranged within the length of about 30 kilobases (kb) with at least two
interveningcellular sequences between the viral sequences (19). Thereversion occurred at a fairly high frequency and all the revertants isolated had suffered deletions within the integrated viral sequences, even if the revertants retained theintactearly region and synthesized large T antigen (18). In the present paper, the molecular mechanism of gene rearrangements which mediates reversion of
SV40-trans-formedcellswasinvestigated by cloningandsequencing the DNA fragments containing the viral sequences from one
transformed mouse line and its large T antigen-negative revertants. The results revealed the following features. (i) The flanking cellular sequences in the transformed cells consist of either a long stretch of A+T or G+C sequences, and there are preferred viral sequences for integration and rearrangement. (ii) In
all
the revertants, both sides of virus-hostjunctions in the transformed cells were retained, butinternal viralsequences weredeleted. Deletion had occurred
either between homologous sequences or between
nonho-mologous sequences. (iii) Small T antigen is expressed in all therevertants.Thesefeaturesarediscussedin relation to the
mechanisms ofreversion in the transformed cells. MATERIALS ANDMETHODS
Cell lines. The transformed cell line W-2K-11 was
estab-lished by infection of C3H/He mouse kidney cell line C3H2K-C4 with virions of SV40 strain777.RevertantsM18, M31, andM42 wereisolated from W-2K-11 cells by negative selection with bromodeoxyuridine as described previously
(19).
Blot hybridization. High-molecular-weight cellular DNA
andrecombinant plasmid DNAwere cleaved tocompletion with various restriction enzymes and electrophoresed on agaroseslabgels.After alkali denaturation, DNAwas
trans-ferred to a nitrocellulose filter or nylon membrane by the 479
on November 10, 2019 by guest
http://jvi.asm.org/
method of Southern (25). The fragments containing SV40 DNA sequences were detected by hybridization to 32p_
labeled SV40 DNA prepared by nick translation (22),
fol-lowedby autoradiography as described previously (19). DNAcloning.High-molecular-weight DNAfrom W-2K-11 cells was cleaved tocomnpletionwith EcoRI. DNAs from the revertants were also cleaved tocompletion with EcoRI and
separatedby electrophoresison anagarosegel.Theslicesof agarose containing the fragment-bearing SV40 sequences were cut out, and DNA was eluted by using glass powder
(31). Phage vector XgtWESXB (28) was propagated by the PDS method describedby Blattneretal. (4), usingthe supF
strain of Escherichia coli Ymel. XgtWESXB DNA was
cleaved with EcoRI, and the cohesive endswere annealed. The large terminal fragments (X arms) were separated by
centrifugation through a 10 to 40% sucrose gradient as
described by Maniatis et al. (17). The annealed arms were
ligated with the EcoRI fragments of cellular DNA and
packaged invitro (3). Recombinantphages containingSV40 sequenceswere screenedby plaque hybridization (2). DNA
fragments containing SV40DNA sequences in recombinant
phageswere subcloned into aplasmid vector(pBR322) and
characterized by restriction mapping. These recombinant
plasmids were designated as pW2K, pM18, pM31, and
pM42.
DNAsequencing. DNA sequences were determinedbythe
method of Maxam and Gilbert (20). SV40 nucleotide
se-quences were numbered according to the SV numbering
00 CQ
T-E
2"1- '..
6.4w_-W
5.2o-
^_
i5.4
4.04-2.8b
kb
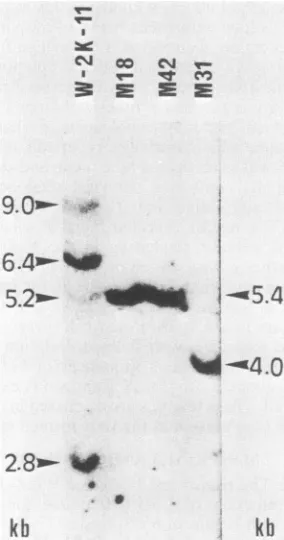
kbFIG. 1. AnalysisofSV40DNAsequencesin W-2K-11 cells and
in therevertants M18, M31,and M42. DNA(20,ug)extracted from
transformed cell line W-2K-11 orfromone of its revertants, M18, M31,orM42,wasdigestedwithEcoRl,electrophoresedona1.2%
agarosegel, transferredtoa nitrocellulosefilter, andhybridized to 32P-labeled SV40 DNA. The lengths of the fragments containing SV40sequences were estimatedfrom thepositions of size marker DNAfragments whichwere runinparallel.
system(30).Theright-handvirus-hostjunctionatnucleotide 4311 in W-2K-11 cells (Fig. 3) was sequenced with plasmid pW2K2.8, which contained the 2.8-kb EcoRI fragment de-tected by Southern blot hybridization of W-2K-11 DNA, from the SV40 Hinfl site at nucleotide 4376 and from a cellularEcoRI site which was about 120 base pairs (bp) from thejunction. Thejunction at nucleotide 4311 in revertant M18 was sequenced withpM18 from a cellular EcoRI site. The left-handjunction at nucleotide 1664 in revertant M18
was sequenced from the SV40 Hindlll site at nucleotide 1493. Thejunctionat nucleotide 1664 in revertant M42 was
sequenced with pM42 from a cellular Hinfl site located within 30 bp of the junction. The virus-virus junction at nucleotides 161 and 1521 in revertant M18 (Fig. 3) was
sequencedfrom the SV40HindlIl site at nucleotide 1493.
Immunoprecipitation.The W-2K-11 cell line and its
rever-tantsgrowntosemiconfluence werelabeled with
[35S]meth-ionine for 2 h. The cell extracts wereprepared by lysingthe cells inlysisbuffercontaining25 mMTris-hydrochloride(pH
8.0), 150 mM NaCl, 0.5% Nonidet P-40, and 150 p.g of
phenylmethylsulfonyl
fluoride per ml. Normal hamsterse-rum was added to the extracts, and the mixtures were
incubated at
0°C
for 30 min. Then, a 10% suspension of Formalin-fixedStaphylococcus
aureusCowan I wasadded,and the mixture was agitated at 4°C for 30 min. After
centrifugation,
the supernatantswerecombined with eitheranti-Thamsterserum ornormalhamsterserumand
incubat-edat
4°C overnight.
Themixturesweresimilarly
treated withbacteria,
andthe pelletswerewashed threetimes withRipabuffer
containing
50 mMTris-hydrochloride
(pH 7.5), 150mM
NaCl,
10% TritonX-100,
0.1% sodiumdodecyl
sulfate,and 150
F±g
ofphenylmethylsulfonyl
fluoride and 10 mg ofdeoxycholate
per ml andsuspended
insample
buffer. Aftercentrifugation,
the supernatants wereelectrophoresed
on asodium
dodecyl
sulfate-14%polyacrylamide
gel. The gelwasdried andfluorographed.
RESULTS
Cloning
of the viralinsertionsinW-2K-11 and the revertantcelllines. To
analyze
the detailed structures ofSV40DNAintegrated
intothe cellularchromosomes intransformedcellline W-2K-11 and inits revertantcell lines M18, M31, and
M42,
we cloned the viral insertions together with flankingcellularsequences fromeach of these celllines by use of A
phage
vectors. As described in a previous paper (19), apossible
arrangement of viral DNA sequences in these celllineswas constructedbased onthe resultsof Southernblot
analysis.
Inthetransformed cell lineW-2K-11,
two genomeequivalents
ofSV40
sequences areserially arranged withinalength
of about 30kb,
with two intervening cellularse-quences. In therevertantcelllines M18, M31,andM42, less than one full copy of the SV40 sequence is retained in a
single
site. A Southern blotanalysis of the DNAfragmentsgenerated by
digestion
of cellularDNAfrom these lineswithrestrictionendonucleaseEcoRIisshownin Fig. 1.
W-2K-11 DNA
yielded
four bands withlengthsofapproxi-mately 9.0,
6.4,5.2,
and2.8kb. To clarifythearrangementof these four
fragments
in thechromosome,
W-2K-11 DNAwas
partially digested
with EcoRI and cloned in X phageCharon4A. From4
p.g
ofpartially digested
W-2K-11 DNA, wecouldconstruct aDNAlibraryof 1.6 x106
recombinantphages.
Afteramplification by
the method of Maniatisetal.(17),
a total of 5 x 105 recombinant phage plaques werescreenedforSV40sequences and three cloneswereisolated.
After several
purification
steps, restriction endonucleasedigestion
ofthese recombinantphages generated
manyon November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.117.259.385.655.2]DELETION OF INTEGRATED SV40
SEQUENCES
481norfragments whichhybridizedtoSV40DNA,
indicating
an occurrence of high frequency recombination within the ACharon 4A vector. Such recombinations were
reported
byClaytonand Rigby (7); recombinantphages carryingtandem
duplications tendedtobe unstable. Inour case,homologous
sequences cloned in the phage vector were prone to be
deleted byrecombinations.
To minimize these homologous
recombinations,
wedi-gestedW-2K-11 DNAtocompletion withEcoRI and cloned in a XgtWESXB phage vector. Fourteen clones containing SV40 sequences were isolated by screening 1.5 x 106 recombinant phages; 10 of these clones contained a 5.2-kb fragmentand 2containeda2.8-kb fragment, but the remain-ing 2clonescorrespondedtononeof four fragments detected by Southern blot analysis. These twoclones seemed to be derived by rearrangement within phage DNA. To clone the
6.4-kb fragment detected by Southern blot analysis, we
digested W-2K-11 DNA to completion with EcoRI, and the DNA fragments witha lengthof about 6.4kb were purified and cloned in XgtWESXB. Three clones containing SV40
sequences were isolated by screening 8 x 105 recombinant
phages.All of these clones contained a6.4-kbfragment. As
mentionedbelow, part of the 6.4- and 2.8-kb fragmentsare
retainedin therevertants,but theSV40sequencesinthe 9.2-and 5.2-kb fragments are completely excised in the rever-tants. Inthisrespect, the9.0-kb fragment wasnot essential
in the present study and was notcloned.
The revertants M18, M31, and M42 generated only one bandeach of about 5.4, 4.0, and 5.4 kb,respectively (Fig. 1).
Cellular DNAs from eachrevertantweredigestedto comple-tion with EcoRI, and the fragments corresponding to these
lengths werepurified. Tencloneswereisolated by screening 6 x 105 recombinant phages constructed from M18. Three and seven clones each were isolated by screening 4 x 105
recombinant phages constructed from M31 and M42. The
sizesofclonedEcoRIfragmentscontainingSV40sequences
were identical to that of the fragment detected by the blot
analysis. These EcoRI fragments were subcloned in the plasmid vectorpBR322 and analyzed further.
trhe
recombi-nantplasmidscontaining thesequencesfrom W-2K-11 weredesignated pW2K6.4, pW2K5.2, and pW2K2.8. The num-bersrepresentthelengths of the cloned fragments in kb. The recombinant plasmids containing the sequences from the revertants were designatedpM18, pM32, andpM42.
Structure of three cloned fragments from W-2K-11 cells.
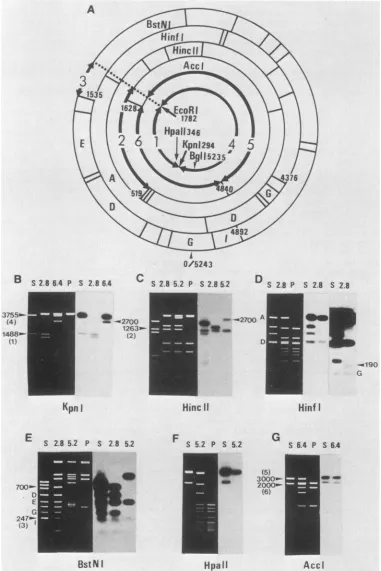
The physical maps of the DNA fragments cloned from transformedcell line W-2K-11 weredeterminedby Southern blothybridization analysisand partial restriction endonucle-asedigestion ofend-labeledfragments. Figure 2 showssome
of these analyses. EcoRI plus KpnI digestion of pW2K2.8 (Fig. 2B) producedtwofragments whichhybridized toSV40 DNA. One of them conligrated with the 1,488-bp fragment corresponding to viral DNA between the KpnI site at nucleotide 294 and the EcoRI site at nucleotide 1782 (Fig.
2A). EcoRI plus HincII digestion (Fig. 2C) produced the 1,263-bpfragment correspondingtoviralDNA between the
HincII site at nucleotide 519 and the EcoRI site (Fig. 2A). Theseresults indicated that the SV40insertion in pW2K2.8
extended counterclockwise from the viral EcoRI site at
nucleotide 1782. Digestion with EcoRI and Hinfl (Fig. 2D)
produced the viral Hinfl fragments A, D, G and one more fragment of about 190 bp which hybridized to SV40 DNA,
indicating that the SV40 insertion in pW2K2.8 was colinear withSV40 virionDNA,andthatthe end of the viral fragment was located about 190 bpapart from the viral Hinfl site at
either nucleotide 4459 or nucleotide 4376. Digestion with
EcoRI and BstNI (Fig. 2E) produced the viral BstNI frag-ments D, E,G, and Iand the247-bpfragmentcorresponding toviral DNAbetween the BstNI site atnucleotide 1535 and theEcoRI site(Fig. 2A) and one morefragment of about 700 bpwhich hybridized toSV40
DNA,
indicating that the other end of thefragment was located about 700 bp apart from the viral BstNI site at nucleotide 4892. These results indicated that the viralinsertion extended at most toi90bpbeyond the Hinfl site at nucleotide 4376.EcoRI plus HincII digestion ofpW2K5.2 (Fig. 2C) pro-duced the 1,263-bp fragment corresponding to the viral
EcoRI-HincII
fragment (Fig. 2A) and the 2.7-kb fragmentwith weak radioactivity, indicating that the SV40 DNA insertion in pW2KS.2 extended counterclockwise from the viral EcoRI site but not much beyond the HincIl site at nucleotide 519. Digestion with EcoRI and HpaII produced only onefragtnent which hybridized toSV40 DNA (Fig. 2F), indicating that the SV40 insertion did not contain the HpaII site ofSV40DNA. These results indicated that the endpoint of SV40 insertion in pW2K5.2 was located between the
HincII site atnucleotide 519 andtheHpaII site at nucleotide 346.
EcoRI andKpnI digestionofpW2K6.4 (Fig. 2B) produced the 3,755-bp fragmentcorresponding to viral DNA between the KpnI site at nucleotide 294 and the EcoRI site at nucleotide 1782 (Fig. 2A) and the 2.7-kb
fragrhent
which hybridized to SV40 DNA. EcoRI plus AccI digestion ofpW2K6.4 (Fig. 2G) produced about 3,000- and 2,000-bp fragments corresponding to viral DNA fragments between the EcoRI and AccI sites and between two AccI sites, respectively. Note that there are twoAccI recognition sites in SV40 strain 777 DNA, the one at nucleotide 1628 and the other atabout nucleotide 4840. These results indicated that the SV40 insertion in pW2K6.4 was colinear with SV40
virion DNA and extended clockwise from the EcoRI site to
beyond the AccI site at nucleotide
i628.
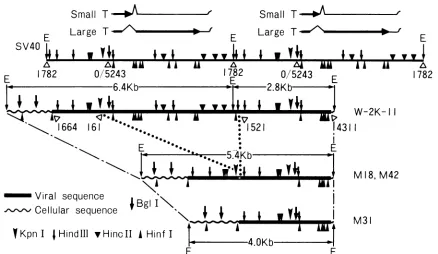
Structures of the cloned DNAs deduced from these ex-periments are schematically presented in Fig. 3 and can be summarized as follows.
SV40 sequences in pW2K2.8 extend counterclockwise
from the viral EcoRI site to the virus-host junction located close to the SV40 Hinfl site at nucleotide 4376, so that the 2.8-kb fragment contains only about 100 bp of flanking cellular sequences. SV40 sequences in pW2K5.2 extend
counterclockwisefrom the viralEcoRI site to thevirus-host junction located between the SV40HinclI site atnucleotide 519 and the
HpaII
site at nucleotide 346, so that the 5.2-kb fragment contains only about 1.3 kb ofSV40sequences and about 3.9 kb offlanking cellularsequences. SV40sequences in pW2K6.4 extend clockwise from the viral EcoRI site at nucleotide 1782 to the virus-hostjunction located close to the SV40AccI
site at nucleotide 1628, so that the 6.4-kb fragment contains almost a unit length of SV40 DNA se-quences and 1.3 kb of flanking cellular sequences. Because of afailure incloning them from the partially digested W-2K-11 DNA fragments, the arrangement of these three frag-ments in a chromosome is unclear. However, as described below, the 6.4- and 2.8-kbfragments seem to be adjacent to each other.Structure of cloned fragments from revertants M18, M31, andM42.Figure 4 shows some of the restriction endonucle-ase analysis ofpM18, pM31, and pM42, and their physical mapsconstructed from these results are schematically repre-sented in Fig. 5.KpnIdigestion ofpM18and pM42 (Fig. 4A) produced 8.35- and 1.35-kb fragments hybridizing to SV40
DNA,indicating that there are two KpnI sites separated by VOL. 49, 1984
on November 10, 2019 by guest
http://jvi.asm.org/
B
S2.86.4
P S2.864
C55_- t2Q
4) _ b 4 2700
1263-888- (2)
K1)
Kpn
I
E
S 2.8 5.2 P S 2.8 5.2
S2.8 5.2 P S 2.8 5.2
a-270 a
Hinc
11
F S 5.2 P S 5.2
eD
D S 2.8 P S 2.8 S 2.8
0
e
al_
D "M _
Hinf
I
G
S 6.4 P S 6.4
(5)
3000" 2000"
(6)
BstNI
Hpall
Accl
FIG. 2. Restrictionenzymemapping of clonedDNAfrom W-2K-11 cells. (A) RestrictionmapofSV40DNA.Restriction fragmentsand nucleotide numbersareindicated. Thefragments are: 1, EcoRI-KpnIfragment (1,488 bp);2, EcoRI-HincIIfragment (1,263bp); 3,
EcoRl-BstNI fragment (247 bp);4, EcoRI-KpnI fragment (3,755 bp); 5, EcoRI-AccI fragment (3,000bp); 6, AccI-AccI fragment (2,000 bp). (B)
Restrictionendonuclease digestion patterns and Southern blot analysis of the recombinant plasmids pW2K2.8, pW2K5.2, and pW2K6.4. DNAs were digested with EcoRI plus the indicated restriction endonuclease. The digests were electrophoresed, stained with ethidium
bromide(whiteonblack panels),transferredtonylonmembrane, and hybridized with32P-labeled SV40DNA(blackonwhite panels).The
DNAsusedwereSV40(S),pW2K2.8(2.8),pW2K5.2 (5.2), pW2K6.4(6.4),and pBR322 (P). The lengthsof the fragments in bpareindicated.
371
14l
--190 G
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.117.498.76.647.2]DELETION OF INTEGRATED SV40 SEQUENCES 483
Small T
Large
T
\
I
_WYA
Jm
I
LA
-6.4Kb-4
WA-E
IVw I i
-rOSh
1- .J.LC U
2.8Kb
t
'I
I
Nt
_~ Viral
sequenceCellular sequence
E
pW2K6.4
pW2K5.2
pW2K2.8
;Bgl
I
Kpn
IIHindIll
vHincll
AHinfIFIG. 3. Structures of three cloned fragments from W-2K-11 cells. The restrictionmapofSV40DNAlinearizedatthesingleEcoRlsiteand thecoding regions for large andsmall Tantigensaswellasthedirection of the viralearly transcriptsareindicatedatthetopof thefigure. Car-etsymbolsrepresentthesites of splicing and the thin,wavylinesatthe 3'end of thetranscriptsrepresentpolyadenylate. Physicalmapsofthe
clonedfragmentswerededucedfromsomeofthe datashown inFig.2.Allmaps aredrawn such that thedirectionofearlySV40 transcription is from lefttoright. Onthe basis ofthis conversion, werefer inthetext tothe leftandright endsof theclonedfragments. E indicatesthe EcoRI site.
1.35kb. KpnI digestionofpM31producedonlyonefragment
of 8.35 kb, indicating that there is only one KpnI site in
pM31.EcoRIplus KpnI digestionof these three recombinant
plasmids(Fig. 4B) produced threefragmentsof4.3,2.7,and 1.35 kb. The 2.7- and 1.35-kbfragments hybridizedtoSV40
DNA sequences, and the 4.3-kb fragment corresponded to the unit length of pBR322. In pM18 and pM42, the total
lengths of 2.7 and 1.35 kb are smaller than that of the inserted EcoRIfragment (5.4 kb) by 1.35kb, suggestingthat the 1.35-kbfragment wasadoublet. These resultsindicated that theorder of thesefragmentsinthe clonedDNA inpM18
and pM42 is 2.7-kb EcoRIlKpnI fragment-1.35-kb KpnIlKpnI fragment-1.35-kb KpnIlEcoRI fragment. If the
1.35-kb KpnIlEcoRI fragment has no SV40sequences, the relative radioactive intensity of the two bands in Fig. 4A
mustbeequaltothatofthetwobandsinFig.4B.Theresults
obtained showed that this was not the case, indicatingthat the outside 1.35-kb fragment contained SV40 sequences. If
SV40 DNA insertion is not interrupted by cellular
se-quences, two KpnI sites separated by 1.35 kb are in SV40
sequences, and the SV40 insertions inpM18 and pM42 are
not colinear with SV40 virion DNA, which has only one KpnI site. As suggested previously (19), SV40 DNA inser-tions in M18 and M42 appeared to be identical. We
con-firmed this assumption bydigestion of pM18 and pM42 with EcoRI and either Hinfl or Hindlll. The electrophoretic
patterns of DNA fragments generated were identical (Fig.
4C).
Toanalyze the SV40 sequences present in pM18, pM31, andpM42 in detail, DNAsweredigested with BstNI, which
cuts SV40 and pBR322 DNAat 16 and 6 sites, respectively.
The cloned DNAs in pM18 and pM42were inserted in the reverse orientation to identify thefragments containing the end of the cloned DNA and theadjacent pBR322sequences.
All the plasmids produced the viral BstNIfragments D, E, G, andI (Fig. 4D). The intensityofthesefragments stained
with ethidiumbromide suggested thatfragments D and E in pM18 and pM42 were doublets or triplets, whereas these
fragments in pM31 appeared to be singlets. The result indicated that the SV40 insertion in pM31 is colinear with SV40 virion DNAandthat the 1.35-kbsequencecontaining the BstNI fragments D and E is duplicated in pM18 and pM42.Since, pM18differsfrompM42 onlyintheorientation
of theinsertion, twofragmentsnotcommontoboth ofthem
mustbethefragmentscontainingthe endof theclonedDNA
andtheadjacent pBR322 sequences.Suchfragmentsarethe 2.0- and0.83-kb fragmentsin pM18and the 2.4-kbfragment
in pM42 (Fig. 4D). Because the total lengths of these
E
14I
E
Iw
4SV40
E
v v
a
;J,mj. v lw v vvA &AA& v v Ava Al
VOL. 49. 1984
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.137.479.78.417.2]A
1831
42 183142
8.35m-a
1.35w- _
K(pnl
B
18
31
42 18
31
42
2.7m- 4 4.
1.35m
EoR
5I
_EcoRI
+
I(pnl
E
S 18 31 42 2.8 P
C
D
S 18 31 42 2.8 P
18 42 18 42
A B
C
D E F G
H
H
inf
I,Hind
IIIEcoRI
F S 18 31 42 2.8 6.4 P
BstNI
G
18 31 42 6.4 18 31 42 6.4
6 ~~~~~~~~3.3
5-B -ml.2 A 2.O-
--C
-_O.8
BD C
D
F
_E
F~~~~~~~~~F_ _ _~~~~~~~
[image:6.612.67.555.78.467.2]Hinf
I
Hindill
EcoRI
+Bgl
I
FIG. 4. Restriction enzyme mapping ofcloned DNA from revertants M18, M31, and M42. DNAs were digested with the restriction
endonuclease indicated below the panel. The digests were electrophoresed, stained with ethidium bromide (white on black panels),
transferredtonylon membrane,andhybridized with32P-labeled SV40DNA(blackonwhite panels inA, B, andG). TheDNAsineach panel are:pM18 (lane 18), pM31 (lane31),pM42 (lane 42), pW2K2.8 (lane2.8), pW2K6.4 (lane6.4),SV40(laneS),and pBR322 (laneP).The lengths
ofthefragments areindicated in kb.
fragments must be equal, pM42 must have a fragment of about 0.4 kb which may comigrate with the viral BstNI fragment E.
The two fragments of 2.0 and 0.83 kb present in pM18 were also present in pM31 (Fig. 4D). The same relations were also seen by digestionofpM18 and pM31 with either Hinfl(Fig. 4E)orHindIII(Fig. 4F). These results indicated that both ends of the cloned EcoRI fragments in pM18and pM31 were identical. Figure 4D, E, and F also shows that one of the fragments containing the end of the cloned fragmentswasalsopresent inpW2K2.8, indicatingthat one endof the cloned DNA inpW2K2.8wasidenticaltothoseof
pM18andpM31. The 2.7-kbEcoRI-KpnI fragment
contain-ing the other end of the cloned DNAs was present in
pW2K6.4, pM18, pM31, and pM42 (Fig. 2B and 4B). The same relations were seen by digestion ofpW2K6.4, pM18, pM31,andpM42 (Fig. 4G)withEcoRI andBglI,inwhichthe BglI fragment containing the viral-hostjunction is 2.0 kb in
pW2K6.4andpM31and3.35 kb inpM18andpM42, owingto the1.35-kb duplication. These datasuggestedthat the other
end ofthe cloned fragment wasalso identical in pW2K6.4, pMl8, pM31, and pM42.
The structures of the cloned DNAs in pM18, pM31, and
pM42arepresented schematicallyinFig. 5togetherwiththe
structureof thejointbetween theSV40sequenceswithin the
6.4- and 2.8-kbfragments. These structurescan be summa-rizedasfollows. SV40 sequencesinthe 5.4-kbfragmentsof M18 and M42 extend clockwise from a virus-hostjunction
located close to the SV40 Hinfl site at nucleotide 4376 to another virus-hostjunction located close to the SV40 AccI siteatnucleotide 1628 throughavirus-virusjunction. There is about 1.35 kb of duplication approximately between nucleotides 150 and 1500 of the SV40 DNA in thefragments.
TheSV40sequencein thecloned 4.0-kbfragmentinpM31is colinear with SV40 virionDNA; however, bothends of the clonedfragmentand the virus-hostjunctionsare identicalto those of the 5.4-kb fragments in pM18 andpM42. Further-more, the left ends of the fragments in these plasmids are identical tothe leftend of the 6.4-kbfragment inpW2K6.4,
and the right ends of the fragments in these plasmids are
-. 2.4
2.0
--0.83
on November 10, 2019 by guest
http://jvi.asm.org/
DELETION OF INTEGRATED SV40 SEQUENCES 485
E
SV40
A
1782
'N4
\A'
161664Ns
Small T
Large
T
Iw it IvI
AA
0/5243
gf
AKh-1782 -E
,
TVv41
<<<A. AT
161
II0..
*I0.
E
*
00-I%
Viral sequence
4Bgl I\
^
Cellular
sequence
Small
T
qrvor T _
E
LUI-WV
4
i
I
w
A
I
mm-1-5
:1521
..
5.4Kb-I
wY1:0
I
0/5243 E
-2.8Kb
t
,,,,
W-2K-14311
E
3,y4'
- .-. m A
I wY'
YKpn
I
I
HindIII
wHincII
AHinf I
A4.0Kb so
E
E
M18,
M42
[image:7.612.92.530.71.325.2]M31
FIG. 5. Structuresof clonedfragmentsfromrevertantsM18, M31,and M42and thatofcorresponding fragmentsfrom W-2K-11 cells.The restrictionmapof SV40 DNA linearizedattheEcoRlsite is shownatthetop.Twocopiesarearrangedinhead-to-tail tandemarrays.The
con-versions arethesameasin Fig.3. The numbersbelow the line indicatethenucleotide numbersaccording toSV numberingsystem (30). Physicalmapsofthecloned fragmentswerededuced fromsomeof the data shown inFig.4.Structures of thefragmentsclonedfrom M18 and
M42areidenticaltothose described in the text.Theclockwise directionisindicatedasrighttoleft in thetext. identicaltotherightend ofthe 2.8-kbfragmentinpW2K2.8.
These results indicate that the 6.4- and 2.8-kb fragments clonedfrom W-2K-11 cellsare adjacenttoeach other; i.e.,
the viral EcoRI sites of both fragments originated from identical EcoRI sites located inside the SV40 DNA. Thus,
W-2K-11cells contain about 1.5copiesofapartiallytandem arrayofSV40DNA andcontainanintact early transcription unit. This structure in W-2K-11 DNA explains the precise
excision ofinfectious SV40afterfusionwith simiancellsas reported previously(19).All therevertantsdonotcontainan
intact early transcription unitand cannotcode for the intact largeT antigen.
DNA sequences ofthe junctions. The SV40 sequences in revertants M18, M31, and M42 were generated by a dele-tion(s) within the integrated SV40 sequences in the trans-formedcell line W-2K-11. Toanalyzethepatternsof
integra-tion and deletion of viral sequences, the base sequences
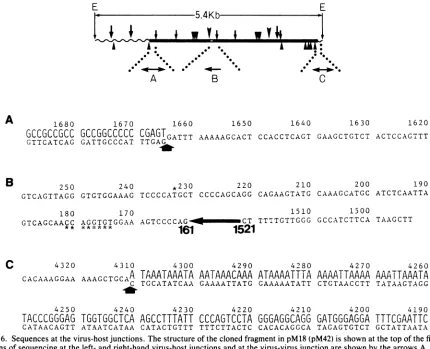
were determined at two virus-host junctions which are present in all of these cell lines and at one virus-virus junctionwhich ispresentintherevertantcelllines M18and M42. The top panel of Fig. 6 shows the positions and
directions in which thesequences weredetermined.The 200-bpHindIII-Hinflfragment (Fig. 6A)containing the left-hand junctionwasobtainedfrompM18andpM42, and the 180-bp
EcoRI-Hinfl fragment (Fig. 6C) containing the right-hand junctionwasobtainedfrompM18andpW2K2.8. The 160-bp KpnI-HindIII fragment (Fig. 6B) containing the virus-virus junction was obtained from pM18. These fragments were end labeled, and their sequences were determined by the method of Maxam and Gilbert(20).
At the left-hand junction, SV40 sequences stretch to nucleotide 1664 and then nonviral sequences begin. The flankingcellularsequences arecomposed almost completely
of guanines and cytosines, and they are organized in an unusual repeating sequence of CCG. At the right-hand
junction, SV40 sequences stretch to nucleotide 4311 and then nonviralsequencesbegin. Contrarytothesequencesof
theleft-handjunction,the first 53 nucleotides in the flanking
cellular sequencesare composed of adenines and thymines except for one cytosine, and they are also organized in a
strikingly unusual repeating sequence of AAAT. As shown
in Fig. 6C, there is patchy homology between the cellular flanking sequences and the corresponding parental SV40
sequences which were displaced by recombination. The
sequencesatthe left- andright-hand junctionswereidentical
inpM18andpM42and inpM18andpW2K2.8, respectively.
The physical maps of the cloned DNA in pW2K6.4, pW2K2.8, pM18, pM42, and pM31 were also identical around these junctions as shown above. These results
strongly suggest that the sequences at both the left- and
right-hand junctionsinW-2K-11cellswereretained in allthe revertants tothe nucleotide sequence level.
DNAsequencesalso indicated that theSV40insertions in all the revertants contain the coding region forviral capsid proteins VP2 and VP3 and, unexpectedly, for small T
antigen.Theright-hand junctionatnucleotide 4311 islocated downstream of the small Tantigen coding regionandthe 5' and 3' splice sites butupstreamof the mRNA polyadenyla-tion signal. Since the right-hand flanking sequences are
composed of exclusively A+T sequences, there are five
consensus polyadenylation signalsof AATAAA. In spite of their normal morphology, these three revertant cell lines seemed to synthesize SV40 small T antigen by usingthese cellular polyadenylation signals. Thisassumption was con-firmed by immunoprecipitation analysis of the cell extracts prepared from the revertants. As shown in Fig. 7, small T
antigen wasdetected in all the revertants, butintactlarge T
antigen was not. The results indicate that SV40 small T
antigen is notinvolved in the maintenance of transformed state.
E
1782
'\IL,
u
Jr
.
;
I
I
wV
0-11
I
",-r
v . .T.
VOL.49, 1984
J.-lr iJ
on November 10, 2019 by guest
http://jvi.asm.org/
E
5.4Kb
v~~
.
0
- 00~~~~~0 *
0 00 0
A
1680
GCCGCCGCC
GTTCATCAG
1670
GCCGGCCCCC
GATTGCCCAT
B
1660
CGAGTGATTT
wV,
I
* 0.
. *
c
1650 1640 1630 1620
AAAAAGCACT CCACCTCAGT GAAGCTGTCT ACTCCAGTTT
250 240 *230 220 210 200 190
GTCAGTTAGG GTGTGGAAAG TCCCCATGCT CCCCAGCAGG CAGAAGTATG CAAAGCATGC ATCTCAATTA
180 170 1510 1500
GTCAGCAAiC AGGTGTGGAA AGTCCCCAG CT TTTTGTTGGG GCCATCTTCA TAAGCTT
* ******
161
1521
C 4320
CACAAAGGAA
4250
TACCCGGGAG
CATAACAGTT
4310 AAAGCTGCAA
4240
TGGTGGCTCA
ATAATCATAA
4300
TAAATAAATA
TGCATATCAA
4230
AGCCTTTATT
CATACTGTTT
4290
AATAAACAAA
GAAAATTATG
4220
CCCAGTCCTA
TTTCTTACTC
4280
ATAAAATTTA
GAAAAATATT
4210
GGGAGGCAGG
CACACAGGCA
4270
AAAATTAAAA
CTGTAACCTT
GATGGGAGGA
TAGAGTGTCT
4260
AAATTAAATA
TATAAGTAGG
4190
TTTCGAATTC
[image:8.612.89.519.76.425.2]GCTATTAATA
FIG. 6. Sequencesatthevirus-host junctions.Thestructureof the cloned fragmentin pM18 (pM42) is shownatthetopof the figure.The
directionsofsequencingattheleft- and right-hand virus-hostjunctions andatthevirus-virusjunctionareshown by thearrowsA,C, andB. (A)Thearrowindicates theleftjunction. Thetopline ofdoublet region, showninlargeletters, is the flanking cellularsequences,and the
bottom line shown in smalllettersrepresents thedisplacedviralsequences.(B) Thesequencesaround thevirus-virus junction inpM18are
shown. Thearrowindicatesthesequencesdeleted. The nucleotides underscored withasterisksaredeleted, and the nucleotideoverscored
withan asterisk at233 is substituted ascompared with the publishedsequence of strain776 (30)as described in thetext. (C) Thearrow
indicates theright junction. Thetopline ofdoublet region is the flanking cellularsequences,and the bottom linerepresentsthedisplaced viral
sequences.
The sequences around the virus-virusjunctions in pM18 indicate that the nucleotideat 161joinstothe nucleotideat 1521, deletingan internal 3.9 kb of viralsequencesinwhich
the intact early transcription unit is located. The parental SV40 sequences around the deletion site 161/1521 indicate that there is almost no homology between the two regions taking part in thejoining, but there is a possible secondary structurewhichmightreduce the effective distance between nucleotides 161 and 1521 (see below). There is a second deletion (eight bases between nucleotides 174and 181) and
one substitution(G-* Tatnucleotide 233)nearthedeletion
site as compared with published sequences of SV40 DNA
strain 776. As described above, W-2K-11 cellswere estab-lished by infection of mouse kidney cell line with SV40 virions strain 777. To distinguish whether these differences
had been generated concomitant with the large deletion or originated from the difference insequences between strains
776 and 777, both SV40 DNAs were electrophoresed after digestion with BstNI, whose recognition sequences are locatedonlyatthe deletion andsubstitution sites. The result indicatedthatthere isan eight-base deletion instrain 777as compared with the published DNA sequence of strain 776, but there is nobase substitution at nucleotide 233.
DISCUSSION
In the present paper, the mechanism ofgene rearrange-mentwith respect to viral sequences in SV40-transformed
mouse cells was studied by cloning the DNA fragments containingviralsequencesfromthetransformed cellline
W-2K-11 and its revertants M18, M31, and M42. In W-2K-11 cells, about two genome equivalents per cell were serially arrangedwithin about 30 kb, with at leasttwo intervening
cellular sequences between the viral sequences (19). To determine the relative positions of these viral sequences, largeDNA fragments generated by partial EcoRI digestion were cloned into phage vector Charon 4A. However, the
cloningof thefragments correspondingtothose detectedby
blot analysis was unsuccessful due to recombination be-tweenthehomologous sequencesduring propagationof the
recombinant phages. The unstable feature of recombinant phages carrying tandem duplication was also reported by Clayton andRigby (7).
The cellular DNA fragments were therefore cloned after completedigestionwithEcoRIto minimizethehomologous recombinations within X phages. The physical maps ofthe
fragments cloned from W-2K-11 cells and the revertants, however,revealed thattwoof the EcoRI fragments
contain-A
-B
I
on November 10, 2019 by guest
http://jvi.asm.org/
DELETION OF INTEGRATED SV40 SEQUENCES 487
M42
M31
M18 M 5 W2K
C4
[image:9.612.61.299.76.263.2]T N T
N T N
T
N T N
T N
FIG. 7. Immunoprecipitation analysis ofSV40antigens synthe-sized in revertants. The cell extracts labeled with[35S]methionine
were precipitated with either anti-T hamster serum(T) ornormal
hamsterserum(N)asdescribedin thetext.Theantigenseluted from
the immunoprecipitates were analyzed by electrophoresis on a sodium dodecylsulfate-14% polyacrylamide gel, followedby fluo-rography.Theblack and whitearrowsindicate thepositionsoflarge and small Tantigens, respectively. Molecularweightswere estimat-ed from thepositionsof thefollowingmarkerproteins:bovineserum albumin, 68,000 (68K); ovalbumin,45K;chymotrypsinogenA, 25K; cytochrome c, 12.5K. Abbreviations: C4, normal parental cell line C3H2K-C4; W2K, W-2K-11 cells; M5, large T antigen-positive
revertant;M18, M31, M42, largeTantigen-negativerevertants.
ing the virus sequences (6.4 and 2.8 kb) are adjacent, comprising 1.5 copies of the SV40 sequences in a partially tandem duplicated form (Fig. 5). The remaining two frag-ments (9.0 and 5.2 kb) containing much shorter SV40
se-quences may be located close tothe 1.5-copy fragment.
Thesequence dataattheSV40-host junctionsinanumber of different cell lines (5, 26, 27) and in SV40 evolutionary
variants (9-12) have suggested that there is no specific integration site in either cellularor virus DNA, but there is
some preference for A- and T-rich sequences in the SV40
DNAatthejunctionandpatchy homology between the viral and cellular sequences. In W-2K-11 cells, the one flanking
sequence is along stretch ofadenines and thyminesand the other is a stretch of guanines and cytosines, and these
flanking sequenceshave characteristicrepeatsof AAATand
CCG, respectively. Similarfeatureswere reported in
SV40-transformedratcell lines in which theflankingsequencesare composed ofasimple sequenceof CArepeatorastretch of adenines and guanines (26). These simple and specific
se-quences are not present at every flanking cellular sequence
atSV40-hostjunctionssofar sequenced; however,the high frequencyofappearanceseemstosuggestsomecontribution of these sequences to integrative recombination or merely reflect thefrequent occurrenceof these simple sequencesin mammalian cells. The cluster of A+T might facilitate the formation ofthe initial recombination owingtoinstability of
hydrogenbonds between adenine and thymine assuggested byGutai andNathans (11).
Itwassuggestedthat the virus-hostjunctionsappeartobe clustered in the 5' half of the late transcriptionunit between nucleotides 300 and 1100 (7). In W-2K-11 cells, the SV40
nucleotide numbersat the left andrightvirus-hostjunctions are 1664 and 4311, and those numbers at the virus-virus junctionsinrevertantsM18 and M42are1521 and 161. These
four sites at the
junction
all relate to thejunctions
in thetransformed cells and the evolutionary variants
previously
reported by otherinvestigators. The left virus-hostjunction
at nucleotide 1664 is located close to the right virus-hostjunction
atnucleotide 1668 intheSV40-transformed
ratcells14B(5).Theright junctionatnucleotide4311 islocated close to the virus-virusjunction at nucleotide 4306 in the
evolu-tionary
variant evlll4(9).Thevirus-virusjunction
atnucle-otide 161 is located close to the virus-virus
junction
atnucleotide 157 in evlll7 (9),and the
junction
atnucleotide1521is locatedclosetothevirus-host
junction
atnucleotide1518 inthetransformedratcells SVRE17(26).The distance between these related
junctions
isonlythree to five bases.*These
relations mightbeanaccident orreflect theselectiveadvantage inan
integration
atthelateregion
tomaintainthetransformed state or selective
replicative advantage
on thevariants,-but these reasons alone cannot account for the above similarities. These data suggest that thereare
specific
sites inSV40DNA sequencestofacilitate
integrative
recom-bination and rearrangement. The viral sequences in thepreferred
recombination sitesmight
be useful fortheintro-duction ofexogenous DNAinto the cellularchromosomes. It should be noted that the viral sequences
preferentially
used for
integrative
recombination with cellular sequencesare also used for the formation of virus-virus
junctions.
These results suggest that virus
integration
into cellular sequencesand theformationof virus-virusjunction
might
be mediated identically or that virus-hostjunctions
do notreflect the initial integration site but reflect the secondary
rearrangements after the initial
integration
event orboth.The
parental
SV40sequencesdisplaced
around thevirus-virus junction in M18 and M42 share little homology but
have sequences
capable
offorming
asecondary
structurewhich might reduce the effective distance between
nucleo-tides 161and 1521. Afew
potential secondary
structures canbe constructed betweentwo50-nucleotidesequencesaround
nucleotides
161 and 1521. The structure shown inFig.
8 isthe most stable structure deduced from calculations
by
the method of Tinoco et al. (29). The relation between thedeletionat,the repeated sequencesand the
secondary
struc-tures has been inferred in E. coli (1) and in the human I-globingenefamily (8)in which"slippedmispairing" during
DNA
replication might
be involved in the deletion. Asshown in
Fig.
8, thereis a short repeat nearthesecondary
structure. If DNAreplication
was started fromtheflanking
cellular sequences or from the SV40
replication origin,
thesingle-stranded
intermediate could form thissecondary
structure.
Figure
9summarizes thestructuresoftheSV40 sequencespresentin thetransformedandrevertantcells. Thereare two typesofdeletionduringthereversion ofW-2K-11cells. One
deletion is seen in M31 in which the SV40 insertion is
colinear with
SV40
virion DNA as a result of a deletionbetween homologous sequenceswithintheduplicated SV40
DNA
region.
This result suggests thatevenin nonpermissivecells a
precise
excision of SV40 DNA occurs without "apermissive factor(s)" supplied by
fusion withpermissive
monkey
cells. Thepermissive
factors may berequired only
for the
replication
of the excised DNA and not for theexcisionof viral DNA from the cellular genomes. Laniaetal.
suggested
similar,possibilities
inpolyomavirus-transformed cells (14). The other deletion is seen in M18 and M42 in which the SV40 DNA insertion is not colinear with SV40virion DNAasaresultof deletion between
nonhomologous
regions
(nucleotides 161 and 1521) within the SV40se-quences.
VOL.49, 1984
on November 10, 2019 by guest
http://jvi.asm.org/
A
5' 4 3'
M18
-CTTATGAAGATGGCCCCAACAAAAAGCTGGGGACTTTCCTTGCTGACTAA-1521
-CTTATGAAGATGGCCCCAACAAAAAGAAAAGGAAGTTGTCCAGGGGCAGC-SV40
* * ***-TTGCATACTTCTGCCTGCTGGGGAGCCTGGGGACTTTCCTTGCTGACTAA-161
B
1544 GOC 144
A*T
COG
G*C
G
*C
C*"G
G
TA-T
1521
C535\Q.GQISS
161AGAAAAGGAAGTTGVT
'
\GGAGCCTIGGGG
IACTT
L.
~/
FIG. 8. Nucleotide sequences andapossible secondarystructurearound thevirus-virusjunction inrevertantM18. (A)SV40sequences aroundthejunction(nucleotides1521/161)arecomparedwith those intworegionstakingpartin thejoining.Thejunctionisindicatedbythe arrow, and sequences common to thetwo regionsareindicatedby asterisks. (B) Potential secondary structuresaround thejunctionwere screenedin the sequences.Themoststablesecondarystructurededuced from calculationbythemethodofTinocoetal.(29) is shown.The shortrepeatsequencesareindicatedby the box.
'431
M18 M42 W-2K-II M31
FIG. 9. Schematicrepresentationof viral sequencesintegratedin W-2K-11cells and therevertants.The viral sequencesintegratedintoa cellularchromosomeareshownbyclosed bars. The dotted lines represent the
flanking
cellular sequences. InW-2K-11cells, about 1.5copiesof viral sequence are integrated in tandem array with partiallyduplicated sequences. Thedeletion of viral sequences had occurredintwo ways; one occurred between homologous sequences, displacing a unit length ofSV40 DNA (M31), and the other occurred between nonhomologous sequences, displacing an internal 3.9 kb of viral sequence (M18, M42). The virus-virus junction (nucleotide numbers
161-*1521) is shownbysmall arrows.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.612.123.487.74.423.2] [image:10.612.69.551.514.683.2]DELETION OF INTEGRATED SV40 SEQUENCES 489
The SV40sequencesretained in M18and M42 are
identi-caldespite the factthattheywereisolatedindependentlyand
have different morphology. Two possible reasons for the occurrence of revertants which have an identical deletion
pattern can be considered. (i) In parental W-2K-11 cells, there may be aprototype of the revertants which has some advantageofexistence,e.g., ahigh efficiencyofplating. (ii) The deletion between nucleotides 161 and 1521 occurs preferentially by forming the secondary structure as stated
above.
Nucleotide sequences of SV40 DNAand flankingcellular
sequences in revertants M18, M31, and M42 predicted the expressionof small Tantigen byusing cellular polyadenyla-tion signals (AATAAA) in the flanking sequences. In these
revertants, the SV40
insertion
stretches to nucleotide 4311andcontains the smallTantigen coding regionandthe 5' and
3' splice sites for 19S mRNA but lacks its polyadenylation signal. However, there are five consensus polyadenylation
signals (AATAAA) within the adjacent cellular sequences. The 19S mRNAfor small Tantigenmustbe polyadenylated
byusingoneofthesesignals.All therevertantsisolated from W-2K-11 cells expressed small T antigen, suggesting a
possiblerole for small Tantigenin the selectiveadvantageof
these revertants. These revertants might help clarify the
functions ofSV40smallT antigen.
ACKNOWLEDGMENTS
We thank Hideo Ideda, Yukio Mishima, Hiroyuki Kato, and Hiroko Sato forexpertadviceon invitropackaging, colony hybrid-ization, base sequencing, and small T antigen assay, respectively. We also thank Nobuo Yamaguchi and Sumio Sugano for valuable discussions.
This workwas supportedby grants from the Ministry of Educa-tion, Science,andCultureof Japan and by grants from the Mitsubi-shi Foundation.
LITERATURE CITED
1. Albertini, A. M., M. Hofer, M. P.Calos, and J. H.Miller.1982. On theformation of spontaneousdeletions: the importanceof short sequencehomologies in the generation of large deletions. Cell29:319-328.
2. Benton, W. D., and R. Davis. 1977. Screening Xgtrecombinant clonesby hybridizationtosingle plaques. Science196:180-182. 3. Blattner,F.R.,A. E.Blechl, K. D. Thomson, H. E. Faber, J. E. Richards, J.L. Slightom,P. W.Tucker, and D. Smithies. 1978. Cloning human fetal y globin and mouse a-typeglobin DNA: preparation and screening of shotgun collections. Science 202:1279-1284.
4. Blattner,F.R.,B.G. Williams, A. E. Blechl, K. D. Thompson,
H.E. Feber, L. A. Furlong, D.J. Grunwald, D.0. Kieffer, D. D. Moore, J.W. Schumm, E. L. Sheldon, and 0. Smithies. 1977. Charon phages: saferderivatives ofbacteriophagelambda forDNAcloning. Science 196:161-169.
5. Botchan,M., J. Stringer,T. Mitchison, and J. Sambrook. 1980. Integrationand excisionofSV40 DNAfromthe chromosome of
atransformedcell. Cell20:143-152.
6. Botchan, M., W. Topp, and J. Sambrook. 1976. The
arrange-ment ofsimianvirus 40 sequencesin the DNA of transformed cells. Cell9:269-287.
7. Clayton,C.E.,and P.W.J. Rigby. 1981.Cloningand character-ization of the integratedviral DNAfrom three lines of SV40-transformedmousecells. Cell 25:547-559.
8. Efstratiadis, A.,J. W. Posakony,T. Maniatis, R. M. Lawn, C. O'Connell, R. A. Spritz, J.K. DeRiel, B.G. Forget, S. M. Weissman, J.L. Slightom, A. E. Blechl, 0. Smithies, F. E. Baralle,C. C.Shoulders, and N.J. Proudfoot. 1980. The
struc-ture and evolution of the human ,B-globin gene family. Cell 21:653-668.
9. Gutai, M. W. 1981. Recombination in SV40-infected cells: nucleotide sequencesatviral-viral recombinantjointsin
natural-ly arising variants. Virology 109:344-352.
10. Gutai, M. W., and D. Nathans. 1978. Evolutionary variants of simian virus 40:nucleotide sequence of a conservedSV40 DNA segment containing the origin of viral DNA replication as an invertedrepetition. J. Mol. Biol. 126:259-274.
11. Gutai, M. W., and D. Nathans. 1978. Evolutionary variants of simian virus 40: cellular DNA sequence and sequences at recombinant joints of substituted variants. J. Mol. Biol. 126:275-288.
12. Hiscott, J. B., D. Murphy, and V. Defendi. 1981. Instability of integrated viral DNA in mousecells transformedby simian virus 40. Proc. Natl. Acad. Sci. U.S.A. 78:1736-1740.
13. Ketner, G., and T. J. Kelly. 1976. Integrated simian virus 40 sequences in transformed cell DNA: analysis using restriction endonucleases. Proc. Natl. Acad. Sci. U.S.A. 73:1102-1106. 14. Lania, L., S. Boast, and M. Fried. 1982. Excision of polyoma
virus genomes from chromosomal DNA by homologous recom-bination. Nature(London)295:349-350.
15. Lavi, S., apd E. Winocour. 1972. Acquisition of sequences homologous to host deoxyribonucleic acid by closed circular simian virus 40deoxyribonucleic acids. J. Virol. 9:309-316. 16. Lee, T. N. H., W. W. Brockman, and D. Nathans. 1975.
Evolutionary variants of simian virus 40: cloned substituted variants containingmultiple initiation sitesforDNA replication. Virology 66:53-69.
17. Maniatis, T., R. C. Hardison,E.Lacy,J. Lauer, C.O'Connell,
D.Quon, G. K. Sim, and A. Efstratiadis. 1978. The isolation of structural genesfrom libraries of eucaryotic DNA. Cell 15:687-701.
18. Maruyama, K., T. Furuta, K. Ueda, and K. Oda. 1982. Proper-ties of T antigen-positive phenotypic revertants isolated from SV40-transformed rat and mouse cells. Microbiol. Immunol. 26:31-40.
19. Maruyama, K., T. Hiwasa, and K. Oda. 1981. Characterization of flat revertantcells isolated from simian virus40-transformed mouse and rat cells which contain multiple copies of viral genomes. J. Virol. 37:1028-1043.
20. Maxam, A. M., and A. Gilbert. 1977. A new method for sequencingDNA. Proc. Natl. Acad. Sci. U.S.A. 74:560-564. 21. McCutchan, T., M. Singer, andM. Rosenberg. 1979. Structure
ofsimianvirus 40recombinantsthatcontainboth host and viral DNA sequences. J. Biol. Chem. 254:3592-3597.
22. Rigby, P. W. J., M. Dieckmann, C. Rhodes, and P. Berg. 1977. Labeling deoxyribonucleic acidtohigh specificactivity in vitro by nick translation with DNA polymerase I. J. Mol. Biol. 113:237-252.
23. Sambrook,J., R. Green, J.Stringer, T. Mitchson, S. L. Hu, and M. Botchan. 1980. Analysis ofthe sites of integration of viral DNA sequences in rat cells transformed by adenovirus 2 or
SV40.Cold SpringHarborSymp. Quant. Biol. 44:569-584. 24. Sambrook,J., H. Westphal, P. R.Srinivasan,and R. Dulbecco.
1968. The integrated state of viral DNA in SV40-transformed cells. Proc. Natl. Acad. Sci. U.S.A. 60:1288-1295.
25. Southern,E.1975. Detection ofspecificsequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98:503-515.
26. Stringer,J.R.1981.Integratedsimian virus 40 DNA: nucleotide sequencesatcell-virus recombinantjunctions.J. Virol. 38:671-679.
27. Stringer, J. R. 1982. DNA sequence homology and chromo-somal deletion at a site of SV40 DNA integration. Nature (London)296:363-366.
28. Tiemeier, D.,L.Enquist,and P. Leder.1976.Improved deriva-tive ofa phage X EK2 vector forcloning recombinant DNA.
Nature(London) 263:526-527.
29. Tinoco, I.,0. C. Uhlenbeck,and M. D. Levine.1971. Estimation ofsecondary structure in ribonucleic acids. Nature (London) 230:362-367.
30. Tooze, J. 1980. Themolecular biologyoftumorviruses,2nd ed. ColdSpring HarborLaboratory, Cold Spring Harbor,N.Y. 31. Vogelstein, B., and D. Gillespie. 1979.Preparative andanalytical
purification of DNA from agarose. Proc. Natl. Acad. Sci. U.S.A. 76:615-619.
VOL. 49,1984