0022-538X/87/082428-10$02.00/0
CopyrightX3 1987,American Society forMicrobiology
The
65,000-Mr DNA-Binding and Virion trans-Inducing Proteins of
Herpes Simplex Virus Type 1
H. S.
MARSDEN,'*
M. E. M. CAMPBELL,' L. HAARR,2 M. C. FRAME,' D. S. PARRIS,3 M. MURPHY,' R. G. HOPE,' M. T. MULLER,4 ANDC. M. PRESTON'Medical Research Council, Instituteof Virology, GlasgowGIl SJR, Scotland'; Department of Biochemistry, University of Bergen, Bergen 5014, Norway2;andDepartment ofMedicalMicrobiologyandImmunology3andDepartment of
Microbiology,4 Ohio State University, Columbus, Ohio 43210
Received 17 February 1987/Accepted27April 1987
The possible identity ofthe herpes simplex virus type 1 (HSV-1) 65K (65,000-Mr) virion protein which stimulates transcription from immediate-early genes with the HSV-1 65KDNA-binding protein was investi-gated. Thetwoproteinswerefound tobe distinctby the threeseparate criteria ofimmunological reactivity, tryptic peptide fingerprinting, andmobilityin two-dimensional gels. UsingHSV-1/HSV-2intertypic
recombi-nantsandaserotype-specific antiserum, welocated thegeneencodingthe 65KDNA-binding proteinbetween coordinates 0.574 and 0.682 onthe HSV-1 genome. The proteinis posttranslationally modified by phospho-rylation. In crudeextractsof HSV-1-infected cells the 65K trans-inducing proteindid notdetectablybindto double-stranded calfthymusDNA under the conditions ofourassay.
Expression ofherpes simplex virus (HSV) genes is
tem-porally regulatedandis consideredto occurinat least three
phases termed immediate-early (IE), early, and lateor(x,
P,
and -y (13, 27, 28, 31, 50). IE genes are expressed in the
absence ofde novo protein synthesis (27) but their
expres-sion is stimulated by a component of the virus particle (5, 42).Thistrans-inducing factor was identified aspolypeptide Vmw65 (11) andis designated 65KTIF in the present study.
The mechanismbywhich 65KTIF stimulates transcription
fromIE genesis notunderstood; however, onepossibilityis that it interacts with some IE-gene-regulatory DNA se-quence. Interestingly, a responder element located several hundred bases upstream of the IE mRNA 5' terminus has
beenidentified andcorresponds tothe consensus sequence
TAATGARATTC (R is purine) (10, 35, 44, 53).
Previously, we identified a major DNA-binding protein in HSV type 1 (HSV-1)-infected cells with an apparent Mr of ca. 62,000 (6). This DNA-binding protein and 65KTIF have the same electrophoretic mobilities in sodium dodecyl
sul-fate-5to12.5polyacrylamidegels, and so we designated the
DNA-binding protein
65KDBP.
It seemedimportant toestab-lish whether
65KDBp
and 65KTIF were one and the sameprotein and whether 65KTIF can bind to DNA. This study
addresses these questions. We characterized the two pro-teins and show that
65KDBp
and 65KTIF are distinct and that65KTIFdoes not detectably bind to DNA under the
condi-tionsofourassay.
MATERIALS AND METHODS
Cells. BHK21clone 13 cells (34) were used throughout. Viruses. HSV-1 strain 17syn+ (9) and HSV-2 strain HG52 (51) were used in this study. The isolation of HSV-1/HSV-2
intertypic recombinants and the determination of their
genome structures have been previously reported (12, 15, 37, 46, 54).
Radioactive labeling. Confluent monolayers in
50-mm-diameterdishes or roller bottles were infected at a
multiplic-ityofinfection of 5 to 20 PFU of HSV-1 per cell. After 1 h,
*Corresponding author.
unadsorbed viruswas removed and the infected cell
mono-layer was radiolabeled. [35S]methionine (specific activity, >1,000 Ci/mmol; Amersham International) was used at a
concentration of 20 p.Ci/ml for most experiments or 280
,uCi/ml fortwo-dimensional (2D) gel electrophoresis;
label-ingwasperformedin Eaglemediumcontainingone-fifth the
normal concentration of methionine and 2% calf serum. Label wasadded 5 hafter the end of theabsorptionperiod, and cultures were harvested after an additional 30 min (pulse) or 18 h. To chase theradioactivity in pulse-labeled cells, monolayers were washed withcomplete medium and incubatedforanadditional6h inthat medium. [3H]mannose
(specific activity, 10to20Ci/mmol; Amersham)wasusedat
a concentration of 100 ,uCi/ml in Eagle medium
supple-mented with 2% calf serum. (The ethanol in which the mannose was suppliedwas removed before use.)Label was added 5 h after the end ofabsorption, and cultures were harvested about 18 h later. To labelwith
[32p]p,,
uninfectedcell monolayers were grown for 3 h in Eagle medium
containing 10 plM
Pi
(one-tenth the normal concentration)and serum which hadpreviously been dialyzed against 0.9%
(wt/vol) NaCl. After virus absorption, infected cells were
washed with and labeled in the phosphate-reduced medium with 140 ,uCi of carrier-free
[32P]P,
(Amersham) for 22 h.Invitro protein synthesis. RNAsamples were translatedin
a fractionated rabbit reticulocyte system treated with
micrococcal nuclease (41, 43).
Hybrid arrest ofin vitro protein synthesis. Hybrid arrest wasperformed as described previously (36, 45) with plasmid
pMC6, which contains 2.4 kilobase pairs of the HSV-1
sequencerepresentingmostof the genome region specifying the 1.9-kilobasemRNA that encodes65KTIF (11).
Purification ofDNA-binding proteins. Ahigh-salt extract of 109BHKcells infected with 12.5 PFU of HSV-1 per cell was preparedasdescribed (14). To reduce the salt concentration, theextract (210 ml) was then dialyzed at 4°C against three changes (2 liters each) of B2 buffer (50 mM NaCl, 20 mM Tris hydrochloride [pH 8.2], 1 mM EDTA, 1 mM 2-mercaptoethanol, 10% glycerol). The extract was dialyzed foratotalof 20 h. Thedialysate was centrifuged at 10,000 x
gfor1h toremovetheslightprecipitate which formed, and
2428
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 65K DNA-BINDING AND trans-INDUCING PROTEINS 2429
DNA-cellulose
eluted with O-05-201M NaCI in B2 buffer
QO to
s:o LO
.26 6 6O
O)
0w-w
-____ _
0.
CL
g
-155K -1130K gB
-87K
-65K
65K- % V
-43K
FIG. 1. Purification of65KDBPby DNA-cellulose chromatography. Ahigh-salt extract ofBHKcells infected with HSV-1 strain 17 and labeled with [35S]methionine wasdialyzed againstB2buffercontaining 0.05 MNaCl and thenapplied toaDNA-cellulose column. Bound proteins were eluted with a linear NaCl gradient (from 0.05 to 0.90 M) and then a step to2 MNaCl. Fractions were collectedand analyzed by SDS-PAGE. (Left panel) Autoradiograph of two gels. The left shows polypeptides eluted by the salt gradient and the first fraction of the
2 MNaCl step (as indicated above the tracks), and the right gel shows polypeptides inthesubsequent fractions of the 2 M NaCl step.Asample of the extract loaded on the column(input) was also analyzed on each of the gels. (Right panel)Polypeptides inthethird and fourthtracks of theright-hand gel (of the left panel) stained with Coomassie brilliant blue.
the supernatant was loaded onto a double-stranded
DNA-cellulose column (1)which hadbeen equilibrated first with
B2 buffer, then with B2 buffer containing 2 M NaCl, and
finally with B2buffer again. DNA-cellulose was madewith
calf thymus DNA(D-1501;Sigma Chemical Co.)and
consti-tuted a 100-ml bed volume packed in a 26-mm-diameter
column. Aflow rateof1 ml/minwas usedthroughout, and
10-ml fractions were collected. Afterthe inputwas loaded,
the columnwaswashed with 350 ml ofB2buffer(about twice
the volume necessary to removeunboundproteins). Bound
proteins were eluted first with a 200-ml saltgradient in B2
buffer,
starting at 0.05 M NaCl and rising to 0.9 M NaClfollowed by a 2.0 M NaCl step in B2 buffer. All fractions
were analyzed by sodium dodecylsulfate-polyacrylamidegel electrophoresis (SDS-PAGE).
In experiments designedto test whether65KTIFbinds to
calfthymus DNA,theprocedurewasscaled down to the use
ofan extract of2 x 107 infectedcells, and theextract(inB2
buffercontaining0.05 M
NaCl
asabove) wasloadedonto a1-mlDNA-cellulose column which had been equilibratedas
described above (14). Bound proteins were eluted
sequen-tially with B2 buffer containing 0.15, 0.6,and 2.0 MNaCl.
Purification of virions. Cell-released virions labeled with
[35S]methionine
wereprepared fromBHKcells infectedwith5 PFU ofHSV-1 percell as described previously (18, 49).
For two-dimensional (2D) gel electrophoresis, the virions
werepurified furtherby centrifugation through
54%
Percoll(in0.025 M sucrose,0.01% bovineserumalbumin,and 5 mM
Trishydrochloride [pH7.5])for30min at 74,000x g(35,000
rpm; Beckman 50Ti rotor). Thevirus-containing band was
collected, diluted in 1 mM EDTA-1 mM Trishydrochloride
(pH 7.5)to atotal volume of4.1ml, andlayeredonto a1-ml
60%
(wt/vol)
sucrose cushion in 10 mM Trishydrochloride
(pH 7.5), andcentrifugationwasperformedat84,000x gfor
90min(30,000rpm;SorvallAH650rotor). Purified viruswas
collected from the
interphase.
The whole procedure wascarried out at4°C.
NP-40 extraction of virions. Virus particles were
sus-pended in 50 mM NaCl-10 mM Tris
hydrochloride (pH
7.5)-i
mM EDTAat1.6 x1010
particles
per mlandadjusted
to a concentration of0.03% in Nonidet P-40 (NP-40). The
mixture was allowed to stand for 1 h at
4°C
and thencentrifuged
at 100,000 x g for 1 h at5°C.
The supernatantwas stored at -70°C.
Immunoprecipitation. Extracts for
immunoprecipitation
were precleared by centrifugation at 11,600 x gfor 5 min
(13,000 rpm; MSE
Microfuge).
A50-,lportion
ofprecleared
virionextract orpartially purified 65KDBPwasincubated for
3 h at4°Cwith(i)10,u ofLP1monoclonal
antibody
(39)plus
10
RI
of rabbit anti-mouseimmunoglobulin
G(Cedarlane
Laboratories),
(ii)
10RI
ofnonimmune ascitesfluid,
or(iii)
10RI
of 13809 or 13810 rabbit serum.Antibody-polypeptide
complexes were
precipitated by
binding
toprotein
A-Sepharose
(1hat4°C)
and washed several times with0.6 MLiCl
containing
0.1 M Tris hydrochloride (pH 8.0) and 1%2-mercaptoethanol,
andboundprotein
waseluted with0.125M Tris
hydrochloride
(pH 6.8)containing
2%SDS,
2%glycerol, and5% 2-mercaptoethanol.
Gel electrophoresis.
Samples
wereprepared,
andSDS-PAGE was carried out by using the buffer system of
VOL.61,1987
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.570.39.528.60.322.2]Man Met
WIC Ext IP Con
pgB-w_
-D
gD-FIG. 2. Immunoprecipitation of proteinsfromanextract(Ext)of [35S]methionine-labeled HSV-1-infected BHKcells with rabbit
an-tiserum 13810(IP)ornonimmune rabbitserum (Con). Asampleof
HSV-1-infected BHK cells labeled with [3H]mannose (Man WIC, i.e.,mannose,whole infectedcell)wasalsoanalyzedtoidentifythe
position to which HSV glycoproteins had migrated. Polypeptides
wereseparated by SDS-PAGE.
Laemmli(33) and either 5to12.5%gradient gelscross-linked with1/20 (wt/wt) N,N'-methylenebisacrylamide or7.5%gels
cross-linked with 1/40 (wt/wt) N,N'-methylene bisacryl-amide. 2D nonequilibrium pH gradient gel electrophoresis
was performed essentially as described by O'Farrell et al. (40) with minor modifications (23). The second dimension
was a9% polyacrylamide gel cross-linked with 1/40(wt/wt) N,N'-diallyltartardiamide.
Treatment with alkaline phosphatase. Bacterial alkaline phosphatase (EC 3.1.3.1; Sigma) was diluted in waterand incubated ata final concentration of15 U/ml for 30 min at
37°C withproteins extracted asfor 2Dgel electrophoresis. Autoradiographyand fluorography. Slabgelsfrom 2D gel electrophoresiswerefixed, infusedwith2,5-diphenyloxazole asdescribed by Bonner and Laskey (8), dried, and exposed
toKodakXAR-5 filmat -70°C. Other gelsweretreated with
En3Hance (New England Nuclear Corp.), dried, and
ex-posed to KodakX-Omat XS1 film at -70°C.
Western blotting (immunoblotting). Proteins were
elec-trophoretically separated by SDS-PAGE and transferred to
nitrocellulose strips (15 by 0.6 cm) witha Trans-Blot
appa-ratus(Bio-Rad Laboratories,Inc.).Antigensimmobilizedon
thenitrocellulose and reacting with antiserumweredetected with 1251I-labeled protein A iodinated by the method of Hunter and Greenwood (29)as described by Towbin et al. (52) with minor modifications (23).
Tryptic peptidemapping.Both the 2MNaClfraction from the DNA-cellulose column containing partially purified 65KDBpanda0.03% NP-40 virion extractcontaining65KTIF
[image:3.612.121.241.71.353.2]1 2 3 4 5
FIG. 3. Specificityof monoclonal antibodies MA1044 and LP1 for65KTIF. Totalcytoplasmic RNA extracted from BHK cells at 7 h afterinfection with 20 PFUof HSV-1wastranslatedin vitro(TOT, track1). Other aliquotsof the RNAwerefirsthybridizedtoplasmid pMC6(HYB, tracks 4 and5)ortopMC6and thehybrid denatured (HYB/DENAT, tracks2 and 3) beforein vitro translation. Mono-clonalantibodiesMA1044 and LP1wereusedtoimmunoprecipitate the in vitro translationproducts (tracks2to 5). Polypeptideswere
resolvedby SDS-PAGE.
VIFRION
Ext Cofi IP
LP1 13810
1 2 3 4
DNA BP
Ext Con IP
LPI 13810
5 6 7 8
qB-
___k-65K-- _,.
gD-: _*
-65K
dyefront-
-FIG. 4. Antigenic distinction between 65KDBp and 65KTIF.
[35S]methionine-labeled NP-40extractsof virionsand DNA-binding proteins (DNA BP) inthe 2 M NaCl eluate shown inFig. 1 were
immunoprecipitated (IP) with monoclonal antibody LP1 orrabbit
antiserum 13810. Nonimmune ascites fluid was used as a control
(Con). Proteinswereanalyzed by SDS-PAGE.
HYB / DENAT
1-- o
O1
I T-n-I
-155K
gB
-65 K
HYB
° J
I"-..4
VI 63-VI
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.367.508.72.366.2] [image:3.612.333.546.462.666.2]HSV-1 65K DNA-BINDING AND trans-INDUCING PROTEINS 2431 65K
DBP '-a
MIX
7. .4
e4 4_ 2
1
3
4- 5,.6,,
7-,¢10
-a
'WI
xb
.:
i o _
a,i
65K TIF
8,9
7- -f,10
9r" h
9- f 0 I
.
k
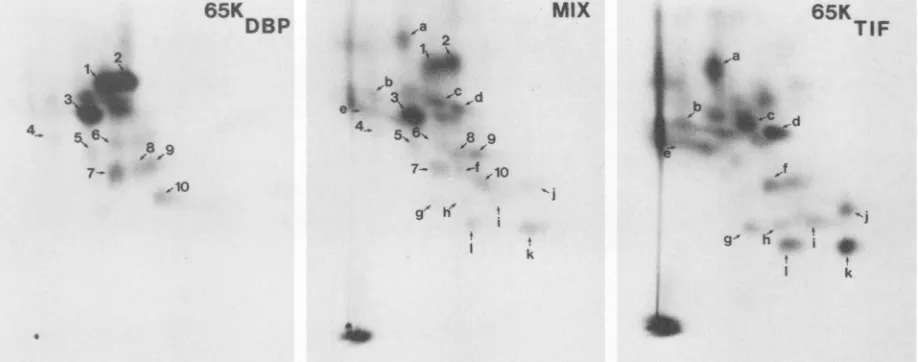
FIG. 5. Fluorographs showing 2D tryptic peptide maps of 65KDBP (left panel), 65KTIF (right panel), and a mix of65KDBPand65KTIF (middle panel). Extracts of DNA-binding proteins and virions prepared as shown in Fig. 4 were individually separated on SDS-10% polyacrylamide gels, and the relevant proteins were excised. Tryptic peptides were prepared from each protein and applied to thin layers of cellulose: the point of application (origin) is shown by the black dot at the bottom left of each panel. Peptides were separated first by electrophoresis (vertical direction of the fluorograph) and then by ascending chromatography (horizontal direction of the fluorograph). Some tryptic peptides have been designated with a number or a letter to facilitate comparison.
were subjected to SDS-10% PAGE. The gels were dried
immediately after electrophoresis. Four spotsof radioactive
ink were placed at the corners of the gel to facilitate
alignment with the autoradiographs. Slices ofthe gel
con-taining the desired polypeptides were cut out, and the
polypeptides wereeluted by the method of Anderson et al.
(2). Theywere desalted by passage through Sephadex G25,
andtheSDSwasremovedbythemethod ofHenderson etal.
(25). Thepelletobtained from thisprocedure was suspended
in 100
RI
ofa 1% solution ofammonium bicarbonatecon-taining 10 ,ugof
L-1-tosylamide-2-phenylethyl
chloromethylketone-treated trypsin. After16h at37°C,anadditional1,ug
of treated trypsin was added and incubation was continued
for 4 h. The peptides were
lyophilized,
oxidized withperformic
acid (26), andthendiluted 50-fold withwaterandlyophilized.
Peptides inpH 2.1electrophoresisbuffer (aceticacid-formic acid-water, 8:2:90) were applied to a spot 4 cm
from each oftwoadjacent edges of cellulose chromatogram
sheets (20by20 cm; no.13255; Eastman Chemical Products,
Inc.). After electrophoresis for 45 min at 600 V, the
chromatogram
wasdried inacurrentof coldair andpeptideswere separated in the seconddimension byascending
chro-matography in
water-butan-1-ol-pyridine-acetic
acid(24:30:20:6). Dried chromatograms were sprayed with
En3Hance
and exposedat -70°C toKodak XS1 film.
RESULTS
Purification of65KDBP. DNA-binding proteins fromBHK
cells infected with HSV-1
(strain
17) wereprepared
andanalyzed
by
SDS-PAGE.Figure
1shows theproteins
intheextractwhich wasloadedontothe column
(input)
and in therelevant fractions eluted from the column
by
increasing
concentrations of NaCl.
65KDBp
eluted across the wholerange ofsaltconcentrationswithtwopeaks,onearound 0.7 MNaCland one in the 2MNaCl step. The
65KDBP
eluting
inthe 2MNaClstephad beensubstantially purified
asjudged
by both autoradiography (Fig. 1,
left)
and Coomassiebril-liant blue
staining
(Fig.
1,right);
thefraction didcontain,
inaddition to 65KDBP, small amounts of the
major
capsid
protein (155K), glycoprotein B or the major DNA-binding protein (130K), and some lower-molecular-weight compo-nents.
Preparation of an
anti-65KDBp
serum. The 2 M NaCl fraction containing65KDBP
was emulsified with an equal volume of Freund complete adjuvant. Rabbits were immu-nized with threeintramuscular injections at 10-day intervals. Each injection contained about 20,ug of65KDBP,
asjudged by the intensity of Coomassie brilliant blue staining of theelectrophoretically separated protein compared to that of
known amounts of
P-galactosidase,
ovalbumin, and bovine serum albumin. Ten days after the third injection serumsamples were collected. To test the specificity ofthe two
antisera(designated 13809 and 13810),immunoprecipitations
were performed with an extract of infected BHK cells
labeled with [35S]methionine. The immunoprecipitateswere
analyzed by SDS-PAGE (Fig. 2). Infectedcellslabeledwith
[3H]mannosewere alsorunonthe samegelasthe markers.
The 13810 serumselectively precipitated from the extract a number of proteins, including a 65K protein and proteins
comigratingwithgBandpgB.The serummay also
immuno-precipitate the major capsid protein (155K); however, the
presence ofa minor comigrating band inthe control immu-noprecipitation raises doubts on this point. A similar
speci-ficity was seenwith serum 13809(datanotshown).
Immunological evidence that 65KDBP and 65KTIF are
dis-tinct. Previousexperiments (11), which established thatthe virion factorresponsiblefortrans-activationofIE genes was
65KTIF,
used the technique ofhybrid arrest of translationand usedmonoclonal antibodyMA1044. This
antibody
spe-cifically precipitated two
polypeptides,
V163 and VI61,which were recognized as the in vitro translation products
corresponding to
65KTIF.
As demonstrated by theexperi-mentshown in Fig. 3, in which plasmidpMC6wasused to arrest
specifically
the translation of65KTIF,
monoclonalantibodyLP1(39)alsogivesaspecificreaction with65KTIF.
Intheexperimentsdescribed belowweused LP1toidentify
65KTIFv
The antigenic relatedness of
65KDBp
and65KTIF
wasinvestigated
by
immunoprecipitation experiments
withVOL.61, 1987
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.78.537.71.252.2]INFECTED
CELLS
A
a4-403....
*-CDNA-- BINDjN
POLY P EPTI DES
8
VI R I NS C
a
4.M~
ci e
A
-C
OUb
A qD
A
....-34
34.
A>-l;:B
FIG. 6. Differential mobilityon 2Dgels of65KDBp and65KTIF. DNA-binding proteinsand extracts of BHKcells infected with HSV-1 strain 17 andpurifiedvirions wereresolvedby2Dgelelectrophoresisandvisualizedby autoradiography. Here and in the 2Dgelsshownin other figures, ampholines (pH 3.5 to 10) were used for electrophoresis in the first dimension and 9% gels cross-linked with
N,N'-diallyltartardiamidefor the second dimension. Sampleswere runindividually (A, B, andC)andaredisplayedasindividual autoradiographs
and as the superposition ofautoradiographs (A+B and A+C). Spots 34 and 403 were previously described in the numbering scheme of
Marsdenetal. (36). SpotsA(actin)anda,b,c, d, e, and fwereusedtoaligntheautoradiographs. Only the relevantportionof thegelsis shown and includes polypeptides with apparent molecular weights of between about 43,000 and 85,000 and relative mobilities in the
nonequilibriumpH gradientgel (36) betweenabout 0.1 and 0.5.
monoclonal antibody LP1 and rabbit antiserum 13810 (Fig.
4). The source of65KTIF was an NP-40 extract of virions
(producedasdescribed in Materials andMethods)knownto
contain theactivity stimulatingIE transcription (Fig. 4,lane
1), whereas the source of 65KDBP was the DNA-binding proteins in the 2 M NaCl eluate shown inFig. 1(Fig.4,lane
5).Antibody LP1precipitateda65Kproteinfrom thevirion
extract (lane 3) butnot the DNA-binding proteins (lane 7),
whereasantiserum 13810precipitateda65Kproteinfrom the
DNA-binding proteins (lane 8) but not from the virion
extract(lane 4). Detergent treatmentwith NP-40at
concen-trations ofup to 3% did not extract from virions any 65K
proteinwhichwas precipitable by antibody 13810 (datanot
shown).Noproteinsweredetected inimmunoprecipitations
with control nonimmune sera (lanes 2 and 6). This
experi-mentdemonstrated that the 65Kprotein in thevirionextract
is distinct from that in the purified DNA-binding proteins.
ThepresenceofgBintheimmunoprecipitateof the virion
extract with antiserum 13810 (Fig. 4, lane 4) wasexpected,
since theserum contains antibodies specificforgB (Fig. 2). However,the presenceofgBin the immunoprecipitatewith LP1 (Fig. 4, lane 3) was unexpected. This observation has been consistentlymade and suggestseitherthatthere might
beaphysicalassociation between65KTIFandgB orthat the
epitope recognized by
LP1 is common to both65KTIF
andgB.
Biochemical evidence that
6SKDBp
and 65KTIFare distin'ct. Two biochemical tests,tryptic
peptide fingerprinting
andmobility
on2Dgels,
wereusedtoinvestigate
therelatednessof65KDBp and 65KTIF. For
tryptic
peptide
fingerprinting
itwasnecessary to
purify
theproteins.
Thiswas doneby
firstpreparing
extracts of virions andDNA-binding
proteins
asdescribed above. These extractswere
separately subjected
to SDS-PAGE, and then the 65K bands were excised and
fingerprinted. Figure
5 shows the pattern obtained for65KDBP (left
panel),
65KTIF(right panel),
and a mixture of65KDBp and 65KTIF
peptides
(middlepanel).
To facilitatecomparison,
10 65KDBPpeptides
(1 to 10) and 12 65KTIFpeptides
(a to 1) were identified in both the patterns of theindividual
proteins
and the mixture. Thisanalysis
of the patterns revealed that they were different and thus demon-strated that the twoproteins
are distinct.To compare the mobilities of65KDBP and 65KTIF by 2D
nonequilibrium pH gradient gel
electrophoresis
(40),infect-ed-cell extracts were made
by
the method of Haarr and Marsden(22),
whereasDNA-binding
protein
and virion extracts wereprepared
as described above. The extractswere
subjected
to 2Delectrophoresis,
and the relevant.- f
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.126.498.70.385.2]HSV-1 65K DNA-BINDING AND trans-INDUCING PROTEINS 2433
-_ _~
0
17 m m 52=3
65K- -:-:..
d?1 - -S Iq
Nx -
-17 cc _- 52 'm cx
_~U
1.1
17 a1 I 52 (ai
cR
4
52 2
RE6 Bx6tl7-1)
Bxl(28-1-1) R12-5
1exilr
(1 ) Bxl(21)Dxl(34-1) Dxl1(48)
FIG. 7. Physical mapping of65KDBp. BHK cells wereinfected with HSV-1 (17), HSV-2 (52), and the intertypic recombinants RE6,
Bx6(17-1), Bxl(28-1-1), R12-5, 17+xllr, Bx1(24), Dxl(34-1), Dx1(48), Fx9(5-8), Bx5(7-2), and Dx1(34-2). Proteins were
dena-tured and analyzed by electrophoresis in 5 to 12.5% gradient
SDS-polyacrylamide gels. Separated polypeptides were transferred
electrophoreticallytonitrocellulosemembranes and incubated with
antiserum 13810. Bound antiserumwas visualized by125I-proteinA andautoradiography.
regionsof theautoradiographs obtained are shown in Fig. 6.
Panel A shows the infected-cell extract, in which gD and spots 34 and 403 are indicated. The nomenclature used is that ofMarsden et al. (36), and spots A (actin) and a to f serve to align the autoradiographs. Panels B and C show
65KDBp
and 65KTIF (arrowheads), respectively.Super-position of the autoradiographs of the infected-cell and
DNA-binding protein extracts (A+B) shows that 65KDBP
corresponds tospot403, whereassuperposition ofthe
auto-radiographs ofthe infected-cell and virion extracts (A+C)
shows that 65KTIF corresponds to spot 34. This result also
demonstrates that65KDBPand 65KTIFaredistinct.
Physical mapping of65KDBP. We nextattempted to locate
theregion ofthe HSV-1genome whichencodes 65KDBP. A
setofHSV-1/HSV-2 intertypic recombinants could be used
since the 13810 antiserum was type specific for 65KDBP.
BHK cells were infected with these recombinants and the
parental strains 17 (HSV-1) and 52 (HG52, HSV-2). The
infected-cell proteinswereseparatedby SDS-PAGE,blotted
onto nitrocellulose membranes, andprobed with antiserum
13810, andbound antibodywas visualizedwith
125I-protein
A. Figure 7 shows that the antiserum reacted with
HSV-1-(e.g., lane1) but notHSV-2-infectedcells (e.g.,lane4)and
that it reacted with cells infected with recombinants RE6,
Bx6(17-1), BX1(28-1-1), 17+x11r(1),
Bxl(24),
Fx9(5-8),Bx5(7-2) and Dx1(34-2) but not R12-5, Dx1(34-1), or
Dxl(48).
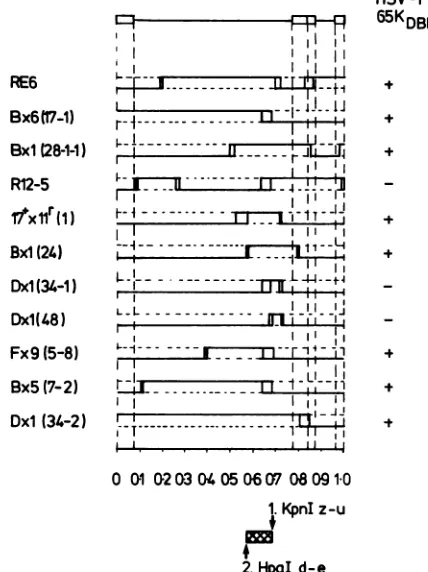
Correlation ofthese data with the genomestructureof therecombinants(Fig. 8) givesamaplocation for65KDBP between approximate coordinates 0.574 and 0.682 with all
dataconsistent. These coordinates aredelimited onthe left
by the HSV-2 HpaI d-e restriction enzyme site and onthe
right by theHSV-1
KpnI
z-urestriction enzyme site.Posttranslationalprocessingof65KDBP.The data
presented
in Fig. 6identified65KDBPas spot 403definedpreviously
ina 2Dgel analysis ofHSV-1-induced
polypeptides (36)
andclassifiedas a
processed
polypeptide.
Wethereforedecidedr-1-~~~~~~
I
i~~~~i
IIA
rn~~~OI~~~
4.42f
L-L
l. ljill
lLLll!l
Fx9(5-8)
Bx5(7-2)
Dxl (34-2) r -'-.
',. . s.i.
HSV-1
65KDBP
+
t
001 2W 3040506 07 08 091-0 1.KpnIz-u
+
2.HpaI d-e
FIG. 8. Summary ofthe genome structuresof the eleven
recom-binants used for the experiment shown in Fig. 7. The genome arrangementof HSVDNAisillustrated in theprototype configura-tion at the top of the figure, showing the long and short repeat sequences andthe long andshort unique regions. Vertical dotted lines correspondtothelimits of thelong and shortrepeat sequences. Those sequences of the recombinant derived fromthe type 1 and
type 2 parent arerepresented by a thick continuous line superim-posed respectively on the upper and lowerof the two horizontal
dottedlines. Crossoverregionsareindicated byone or twovertical lines between the thickcontinuous horizontal lines. The distance betweentwovertical lines indicates theremaining region of
uncer-tainty for thatcrossover event. When theregion ofuncertaintywas
small,the crossover appears asasingle vertical line.Thescaleatthe
bottom isexpressedasfractions of the genomelength. The
induc-tion orlack of induction of65KDBP by each recombinant is indicated
on the right-hand side of the figure (+ or -, respectively). The deduced map location of65KDBP is shown by the cross-hatched
area,anddelimiting restriction sites in thegenomeinHSV-1(I)and HSV-2
(4)
areKpnIz-u andHpaId-e, respectively.to investigate theprocessing eventsinvolved in itsgenesis.
Infected cells were pulse-labeled for 30 min at 5 h after
absorption with
[35S]methionine
andeither harvestedimme-diatelyorchased in unlabeled medium for 6 h. Anextractof
the infected cells was made, and proteins from
aliquots
of theextract wereimmunoprecipitated
with antiserum 13809. The extractsandimmunoprecipitates
wereanalyzed
by
2Dgel electrophoresis, and relevant regions ofthe
autoradio-graphsareshown in the left fourpanels of
Fig.
9. The data show that spots39, 40, 41, 42,and315(previously
identifiedas short-lived-precursor or transient-intermediate
polypep-tides[36]) werelabeled
during
thepulse
and thatduring
the chase the more basic of these spotsdisappeared
and the moreacidic 403 spotappeared.
We next investigated whether
65KDBP
wasphosphory-lated. Infected cells were labeled with [32p]p;, and extracts VOL.61, 1987
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.69.301.72.268.2] [image:6.612.330.544.76.362.2]CE LL
EXTRACT
3 14 3 -A2*2X
P U L S E
4 03 34
Amil.l..:~~~~~...
d~~~~~~~~~~~~~~~~
--..
:=:
:X
IMMUNE
PR E
CI PI
TAT
ION
315 39- 42
4,i
P ULS E
TREATMENT
WITH
PHOS
P HATA SE
rI
403
.
+ ENZYME
403 34
PU LSE -CHA SE zE PU LS E - CHAS E
35
S-
METHI0N
I NE- ENZYME
32 P.
FIG. 9. Posttranslational modification of65KDBP. Infected cellswerepulse-labeledwith [35S]methionine (upperleft and middlepanels), and then thelabel waschased(lower left and middlepanels).Cellextractsweremade(left panels),andaliquotswereimmunoprecipitatedwith antiserum 13809(middle panels). Phosphorylation of65KDBPwasevaluated by electrophoresis of32P-labeled infected-cell proteins after digestion with alkaline phosphatase (+enzyme; upper right panel) or treatment with H20 as a control (-enzyme; lower right panel).
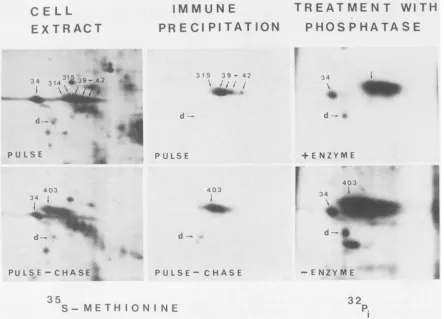
Numbered arrows show previously identified virus-induced polypeptides. Spot d is a cellular polypeptide used here to align the autoradiographs.Theacidic end ofthegel isontheleftof thefigure.
for2D gel
electrophoresis
were prepared. Oneportion wasincubatedwithalkalinephosphataseand the other withH20
as a control. The autoradiographs show that spot 403
(65KDBP) was heavily phosphorylated and that digestion
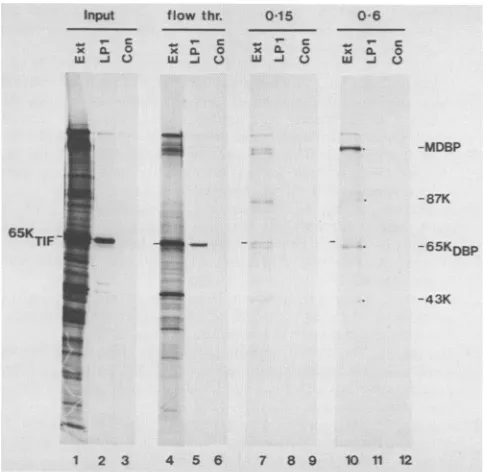
with alkaline phosphatase caused much ofthe label to be lost, particularly fromthemoreacidicforms of theprotein. 65KTIF does not bind detectably to calf thymus DNA. To address the question whether 65KTIF binds to DNA an extractof HSV-1-infected BHK cellswaschromatographed
on double-stranded calf thymus DNA-cellulose; we then
testedbyimmunoprecipitation forthe presenceof65KTIF in
theinputtothe column and in the variousfractions eluting
from the column. Figure 10 shows that 65KTIF was precip-itated with the monoclonal antibody LP1 from the material loaded onto the column (Input) and also from the material which did not bind to the column (flow thr.). It was barely detectable in the material eluted by 0.15 M NaCl and was not detectable in the 0.6MNaClelution. In this experiment no proteinswereelutedfrom the column with 2.0 M NaCl. That thecolumn was able to bind proteins was demonstrated by thesimilarity of the polypeptide profiles of the 0.15 and 0.6 MNaCl eluatescontainingthe major DNA-binding protein, 87K and 43Ktothoseobservedin ourprevious experiments (6)andalsoby the presenceof immunoprecipitable 65KDBP in the eluates (data not shown). We conclude that 65KTIF
does not bind
detectably
to double-stranded calfthymus
DNAundertheconditions ofourassay.
DISCUSSION
Theexperiments reportedhereestablishthat thereare two
distinct
proteins
inducedbyHSV-1 which have verysimilarelectrophoretic mobilitiesin
SDS-polyacrylamide
gels.One,
65KTIF,isacomponentof
purified
virions and isresponsible
for the stimulationof
transcription
fromIE genes(5, 11,42).The other, 65KDBP, is a major
DNA-binding
protein (6)whichmightbeexpectedtoplaysomeregulatoryorcatalytic role.
DNA-cellulose chromatography yielded a preparation of 65KDBP containing only minor amounts ofother proteins.
Thisproteinhasunusualchromatographic propertiesin that
it elutes across the entire salt gradient, with a gradual
peakingaround 0.65Mfollowedbyasecondpeak in the 2 M
NaCl wash. Thepossibilitythatthis behaviorwasduetothe presence of 65Kproteins other than 65KDBPwasexamined
by subjecting everyalternate fraction toanalysis by 2D gel
electrophoresis. Noproteinother than65KDBPwasdetected
andno difference in the distribution ofradioactivity within 65KDBP was apparent in any of the fractions (data not shown). Thusthereasonwhy65KDBP is eluted oversuch a
:1
34 i
mok"
k
4b
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.85.529.71.390.2]HSV-1 65K DNA-BINDING AND trans-INDUCING PROTEINS 2435 Input
1; 0. 0
.. .
flowthr.
x 0. 0
lU U3
015
_ c
x ffi o
W -J V
0.55 0-6
W
%-X CLO
L(
0.0)
54K
54K
I
-MDBP
0.60 0.65 0.70
58K 70K 42K
I ^_
85K 9C
17.8K
UTPas
42K 33K/31K
*~ I-4)
-87K
64K 65KTIF
_ _
6KTIF
- I __
~~~~~~~~~~~~~~~~~~...
...._ _
-65KDBP
OSKDBP
-43K
1 2 3 4 5 6 7 8 9 10 11 12
FIG. 10. Failure of65KTIFtobindto double-stranded calf
thy-musDNA-cellulose. Ahigh-saltextractof BHK cells infected with HSV-1and labeled with[35S]methionine (Input) wasloadedontoa
1-ml DNA-cellulose column. Proteins not binding to the column (flowthr.)were collected,aswellasproteinseluted with0.15,0.6,
and 2.0 MNaCl. Thesevariousfractions(Ext;tracks1, 4, 7, and10)
were precipitated with the anti-65KTIF monoclonal antibody LP1
(LP1; tracks 2, 5, 8, and11)andacontrol nonimmune ascitesfluid
(Con;tracks3, 6, 9,and12).Proteinswereanalyzed bySDS-PAGE.
DNA-binding proteins elutedwith 0.15 and 0.6 MNaCl(tracks 7and
10)include themajorDNA-binding protein (MDBP) and the 87K, 65KDBp, and 43Kproteins. Noproteins weredetected in the 2.0 M eluate(not shown). All tracks received equalexposures andwere
from the same gel though they have been realigned to facilitate
comparison. All the extracts were diluted 10-fold before
electro-phoresis.
broad range of salt concentrations remains obscure but might reflect its differential affinity for subsets of DNA sequences in calf thymus DNA or interactions involving other proteins on thecolumn. These possibilities remain to
be tested.
We have now performed six separate purifications of
65KDBP.Inonly three of thesewastheproteinsocleanlyand
preferentially eluted in the 2 M NaCl step. Inother prepa-rations it eluted before the end of the 0.05to 0.90 M NaCl saltgradient. Again thebasisforthisvariabilityisnotknown. The absence of65KTIFin anyof the fractionseluted from the DNA-cellulosecolumn(Fig.4 and 10)demonstrates that
65KTIFdoesnotdetectably bindto calfthymusDNA under
the conditions of this assay. We have furthermore been unable to detect binding of65KTIF isolated from partially purified virions to DNA fragments which contain the TAATGARATTC element(unpublished data).
It is possible that65KTIFcould bind toDNAby
associa-tion with other viral orcellularproteins. Precedents forthis
possibility exist. Purified adenovirus type 2 ElA protein, a
trans-activating proteinwhich induces efficienttranscription ofearly adenovirus genes (7, 30) does not bind DNA(16). However ElA contained in crude extracts of infected cells
does associate with DNA, suggesting that ElA interacts
indirectly with DNA (32). Another example isthe HSV IE
regulatory protein IE175 (or ICP4), whichdoes notbyitself
FIG. 11. Transcripts and proteins mapping within and close to the map location of65KDBp. The scale of the genome has been expanded to allow individual transcripts and proteins mapping between approximate coordinates 0.55 and 0.73 to be depicted. The hatched barat thebottomof thefigure shows the limits for the map location of65KDBP.
bind to DNA but appears torequire a cell protein forbinding (19). So far we have made no attempt toidentify conditions under which
65KTIF
might bind to DNA. Our observations suggest, however, that 65KTIF does not stimulate IE gene expression by directinteraction with naked DNA and there-fore thatrecognitionofIE-gene-specifictranscriptionsignals such asTAATGARATTC may involve cell proteins.By means of HSV-1/HSV-2intertypicrecombinants and a type-specific antiserum, the gene encoding
65KDBP
was mapped to between restriction enzyme sitesHSV-2 HpaI d-e and HSV-1z-u(approximate coordinates of 0.574 and 0.682,respectively). Figure 11 shows the transcripts and proteins
mappingwithin andcloseto thisregion. The map is based on
datafrom several sources(2, 3, 11, 20, 21, 24; P. A.Schaffer, E. K. Wagner, G. B. Devi-Rao, and V. G. Preston, in Genetic Maps 1986, in press). For the following reasons, many of the transcripts can be excluded as potentially
encoding 65KDBP. (i)Some encode a differentprotein, e.g.,
gC (21),
rr,
(the large subunit of ribonucleotide reductase [48], rr2 (the small subunit of ribonucleotide reductase[4,17,38]),65KTIF(11; thismanuscript),and dUTPase(47).(ii)The
two5'coterminaltranscripts specifyingthe 54Kprotein (24)
probablyshare the samecoding region, which istotheleft of
0.574. (iii)The 17.8Kprotein (21)is too small tobe65KDBP
(Fig. 3). Thus only five identified transcripts remain which
might potentially encode 65KDBP. These specify in vitro
translationproducts of 58,64,85,and 70kilodaltonsand the
proteinmapping betweenthe 64kilodalton productand
gC.
ACKNOWLEDGMENTS
WethankJ. H.Subak-Sharpe,inwhose institutethe bulk ofthis workwascarried out,for his interest intheworkandfor his critical reading of the manuscript. We thankTony Minson forthe mono-clonalantibodyLP1,AnneCrossfor thenonimmune ascites fluid, and Sigrid Overnes who provided skillful technical assistance to
L.H.
L.H. acknowledges financial support from the Norwegian Re-search Council for Science and the Humanities and from the NorwegianSocietyfor FightingCancer. D.S.P. and M.T.M. were supportedinpartbyPublicHealthService grantGM34930from the
NationalInstitutesof Health.
LITERATURE CITED
1. Alberts,B.,andG. Herrick. 1971. DNAcellulose chromatogra-phy. Methods Enzymol.23:198-217.
VOL.61, 1987
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.55.298.67.303.2] [image:8.612.321.555.69.207.2]2. Anderson, C. W., P. R. Baum, and R. F. Gesteland. 1973. Processing of adenovirus 2-induced protein. J. Virol. 12:241-252.
3. Anderson, K. P., R. J. Frink, G. B.Devil,B. H.Gaylord,R. H. Costa, and E. K. Wagner. 1981. Detailed characterization of the mRNAmappingintheHindlIl fragmentKregionof theherpes simplex virustype 1 genome. J. Virol. 37:1011-1027.
4. Bacchetti, S., M. J. Evelegh, and B. Muirhead. 1986.
Identifica-tion andseparation of the two subunitsof the herpes simplex
virusribonucleotide reductase. J. Virol. 57:1177-1181. 5. Batterson, W., and B. Roizman. 1983. Characterization of the
herpes simplex virion-associated factor responsible for the
induction of alphagenes. J. Virol.46:371-377.
6. Bayliss,G.J.,H.S. Marsden,andJ.Hay.1975.Herpessimplex virus proteins: DNA-bindingproteins in infected cells and inthe
virusstructure.Virology68:124-134.
7. Berk, A. J., F. Lee, T.Harrison, J.Williams,andP. A.Sharp.
1979.Pre-early adenovirus5geneproduct regulatessynthesis of earlyviral messenger RNAs. Cell 17:935-944.
8. Bonner, W. M., and R. A. Laskey. 1974. A film detection methodfor tritium-labelled proteins and nucleic acids in poly-acrylamide gels. Eur. J.Biochem. 46:83-88.
9. Brown, S. M., D. A. Ritchie, and J. H. Subak-Sharpe. 1973.
Genetic studies with herpes simplex virustype 1.The isolation
oftemperature-sensitive mutants, their arrangement into com-plementation groups and recombination analysis leading to a linkagemap.J. Gen. Virol. 18:329-346.
10. Bzik, D. J., and C. M. Preston. 1986. Analysis of DNA
se-quenceswhichregulate thetranscription of herpessimplexvirus immediateearlygene 3: DNA sequencesrequiredfor enhancer-likeactivity and response totrans-activation by avirion poly-peptide. Nucleic AcidsRes. 14:929-943.
11. Campbell, M. E. M., J. W. Palfreyman, and C. M. Preston.
1984. Identification of herpes simplex virus DNA sequences which encode atrans-acting polypeptide responsible for
stimu-lation of immediateearlytranscription. J. Mol. Biol. 180:1-19. 12. Chartrand, P., N. M. Wilkie, and M. C.Timbury. 1981.Physical mapping oftemperature-sensitive mutations ofherpessimplex virustype 2by marker rescue. J.Gen. Virol. 52:121-133. 13. Clements,J. B., R. J. Watson, and N. M. Wilkie. 1977.Temporal
regulation of herpessimplex virustype 1transcription:location oftranscripts onthe viralgenome.Cell 12:275-285.
14. Dalziel, R. G., and H. S. Marsden. 1984. Identification oftwo herpes simplex virus type 1-induced proteins (21K and 22K) which interact specifically with the a sequence of herpes simplex virus DNA. J. Gen. Virol.65:1467-1475.
15. Davison, A. J., H. S. Marsden, and N. M. Wilkie. 1981. One functionalcopyof the long terminalrepeat genespecifying the
immediate early polypeptide IE 110 suffices for a productive infection ofhumanfoetallung cells by herpessimplex virus.J.
Gen. Virol. 55:179-191.
16. Ferguson, B., B. Krippl, 0. Andrisani, N. Jones, H. Westphal, and M. Rosenberg. 1985. ElA 13S and 12S mRNA products
made in Escherichia coli both function as nucleus-localized
transcription activators but do not directly bind DNA. Mol.
Cell. Biol. 5:2653-2661.
17. Frame, M. C., H. S. Marsden, and B. M. Dutia. 1985. The
ribonucleotide reductase induced byherpessimplex virustype 1
involves minimally acomplex of two polypeptides (136K and
38K). J.Gen. Virol.66:1581-1587.
18. Frame, M. C., D. J. McGeoch, F. J. Rixon, A. C. Orr, and H. S. Marsden. 1986. The 10K virion phosphoprotein encoded by gene US9 from herpes simplex virus type 1. Virology 150:321-332.
19. Freeman, M. J., and K. L. Powell. 1982. DNA-binding
proper-tiesof a herpes simplex virus immediate-early protein. J. Virol.
44:1084-1087.
20. Frink,R.J., K. P. Anderson, and E. K. Wagner. 1981. Herpes
simplex virus type 1 HindIIIfragment L encodes spliced and
complementary mRNAspecies.J. Virol. 39:559-572.
21. Frink, R. J., R. Eisenberg, G. Cohen, and E. K. Wagner. 1983.
Detailed analysisof theportionof the herpes simplex virus type 1genomeencoding glycoprotein C. J. Virol. 45:634-647.
22. Haarr, L., and H. S. Marsden. 1981. Two-dimensional gel analysisofHSVtype 1-inducedpolypeptidesandglycoprotein processing.J.Gen. Virol. 52:77-92.
23. Haarr, L.,H.S.Marsden,C. M. Preston,J. R.Smiley,W. C.
Summers, and W. P. Summers. 1985. Utilization of internal
AUG codons for initiation of protein synthesis directed by
mRNAs from normal and mutant genes encoding herpes simplexvirus-specified thymidinekinase. J. Virol. 56:512-519. 24. Hall,L.M.,K.G.Draper,R.J.Frink,R. H.Costa,and E. K.
Wagner. 1982.Herpes simplexvirus mRNAspecies mappingin EcoRIfragment I.J. Virol. 43:594-607.
25. Henderson, L. E., S. Oroszlan, and W. Konigsberg. 1979. A
micromethod forcomplete removal ofdodecyl sulphate from
proteins by ion-pairextraction. Anal. Biochem. 93:153-157. 26. Hirs, C. A. W. 1967. Performic acid oxidation. Methods
En-zymol. 11:189-199.
27. Honess,R.W.,and B.Roizman.1974.Regulationofherpesvirus macromolecularsynthesis. I. Cascaderegulationof the
synthe-sisofthree groupsofviralproteins. J.Virol. 14:8-19. 28. Honess, R. W., and B. Roizman. 1975. Regulation ofherpes
virusmacromolecular synthesis: sequential transition of poly-peptide synthesis requiresfunctional viralpolypeptides. Proc.
Natl.Acad. Sci. USA72:1276-1280.
29. Hunter, W. M., and F. C. Greenwood. 1962. Preparation of
iodine I3'l-labelled human growth hormone of high specific activity.Nature(London) 194:495-496.
30. Jones,N.,and T. Shenk. 1979. An adenovirustype5earlygene
functionregulatesexpressionof otherearly viral genes. Proc. Natl.Acad. Sci. USA 76:3665-3669.
31. Jones,P. C.,andB. Roizman.1979. Regulationofherpesvirus macromolecular synthesis. VIII. The transcription program
consists ofthreephasesduringwhich bothextentof
transcrip-tion andaccumulation ofRNAin thecytoplasmareregulated.J. Virol. 31:299-314.
32. Ko, J.-L., B. L. Dalie, E. Goldman, and M. L. Harter. 1986. Adenovirus-2 early region 1A protein synthesized in Esche-richia coliextracts indirectly associates with DNA. EMBO J. 5:1645-1651.
33. Laemmli,U. K. 1970.Cleavageof structuralproteins duringthe
assembly ofthe head ofbacteriophage T4. Nature (London) 227:680-685.
34. Macpherson, I., and M. G.Stoker. 1962. Polyoma transforma-tion ofhamstercell clones: an investigationofgeneticfactors affectingcellcompetence. Virology16:147-151.
35. Mackem, S., andB. Roizman. 1982. Structural features ofthe
herpes simplex virus agene 4, 0, and 27 promoter-regulatory
sequences which confer a regulation on chimeric thymidine kinasegenes. J. Virol.44:939-949.
36. Marsden,H.S.,L.Haarr,andC. M. Preston. 1983.Processing ofherpes simplexvirusproteinsandevidencethattranslation of thymidine kinase mRNA is initiated at three separate AUG codons. J.Virol.46:434-445.
37. Marsden,H.S.,N. D.Stow,V.G.Preston,M.C.Timbury,and N. M. Wilkie. 1978.Physical mappingofherpes simplex virus-inducedpolypeptides. J. Virol.28:624 642.
38. McLauchlan, J.,andJ.B.Clements. 1983. Organizationofthe
herpes simplex virus type 1 transcription unit encoding two
early proteinswith molecular weightsof140000and40000. J.
Gen. Virol. 64:997-1006.
39. McLean, C.,A.Buckmaster,D.Hancock,A.Buchan,A.Fuller,
and A. Minson. 1982. Monoclonal antibodies to three
non-glycosylated antigens ofherpes simplex virus type 2. J. Gen. Virol.63:297-305.
40. O'Farrell,P. Z., H. M. Goodman, andP. H. O'Farrell. 1977.
High-resolution two-dimensional electrophoresis of basic as
wellasacidicproteins. Cell 12:1133-1141.
41. Pelham,H. R.B.,and R.J. Jackson.1976. Anefficient
mRNA-dependenttranslation system fromreticulocytelysates. Eur.J. Biochem. 67:247-256.
42. Post, L. E.,S. Mackem, and B. Roizman. 1981. Regulation of
genes ofherpes simplex virus: expression of chimeric genes
produced byfusion ofthymidinekinasewithagene promoters.
Cell24:555-565.
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 65K DNA-BINDING AND trans-INDUCING PROTEINS 2437 43. Preston, C. M. 1979. Control ofherpes simplex virus type 1
mRNA synthesis in cells infected with wild-type virus orthe temperature-sensitive mutant tsK. J. Virol.29:275-284. 44. Preston, C. M., M. G. Cordingley, and N. D. Stow. 1984.
Analysis of DNA sequences which regulate the transcription of a herpes simplex virus immediate early gene. J. Virol. 50:
708-716.
45. Preston, C. M., and D. J. McGeoch. 1981. Identification and
mapping oftwopolypeptides encoded withinthe herpessimplex
virus type 1 thymidine kinase gene sequences. J. Virol.
38:593-605.
46. Preston, V. G., A. J. Davison, H. S. Marsden, M. C. Timbury, J. H. Subak-Sharpe, and N. M. Wilkie. 1978. Recombinants between herpessimplex virustypes 1and2:analysis ofgenome structures and expression ofimmediate early polypeptide. J. Virol. 28:499-517.
47. Preston, V. G., and F. B. Fisher. 1984. Identification of the
herpes simplex type gene encoding the dUTPase. Virology
138:58-68.
48. Preston, V. G., J. W. Palfreyman, and B. M. Dutia. 1984. Identification of a herpes simplex virus type 1 polypeptide which is acomponent ofthe virus-induced ribonucleotide
re-ductase.J. Gen. Virol. 65:1457-1466.
49. Stevely, W. S., M. Katan, V. Stirling, G. Smith, and D. P. Leader. 1985. Protein kinase activities associated with the
virionsof pseudorabies and herpes simplex virus. J. Gen. Virol.
66:661-673.
50. Swanstrom,R.I., K.Pivo, and E. K. Wagner.1975. Restricted transcription of the herpes simplex virus genome occurring earlyafterinfection and in thepresenceof metabolicinhibitors.
Virology66:140-150.
51. Timbury,M. C. 1971.Temperature-sensitivemutantsofherpes simplex virustype2. J.Gen. Virol. 13:373-376.
52. Towbin, H., T. Staehelin, and J. Gordon. 1979.Electrophoretic transfer of proteins from polyacrylamide gels tonitrocellulose sheets: procedure andsomeapplications.Proc. Natl.Acad. Sci.
USA76:4350-4354.
53. Whitton, J. L., F. J. Rixon, A. J. Easton, andJ. B.Clements.
1983.Immediate-earlymRNA-2of herpessimplexvirus types1
and 2 is unspliced: conserved sequences around the 5' and 3'
terminicorrespondtotranscription regulatory signals. Nucleic
Acids Res. 11:6271-6287.
54. Wilkie, N. M., N. D. Stow, H. S. Marsden, V. Preston, R.
Cortini,M.C.Timbury, and J. H. Subak-Sharpe. 1978. Physical mappingofherpes virus-coded functions andpolypeptides by markerrescueandanalysis of HSV-1/HSV-2 intertypic
recom-binants. In G. De-Thd, W. Henle, and F. Rapp (ed.), Oncogenesisandherpesviruses. III. Proceedings,p. 1-11.
Pub-lication no. 24, part 1. International Agency for Research on
Cancer, Lyon. VOL. 61, 1987