The
Herpes
Simplex
Virus
1Protein Kinase
Encoded by the
US3
Gene Mediates Posttranslational Modification
of the Phosphoprotein
Encoded by the UL34 Gene
FRANCESC. PURVES,DAVID SPECTOR, AND BERNARD ROIZMAN*
TheMarjorieB. Kovler Viral Oncology Laboratories, The University of Chicago, 910 East58th
Street,
Chicago,
Illinois 60637Received22 May1991/Accepted5August 1991
Earlier studies haveshown thataherpessimplex virus 1 (HSV-1)openreading frame,
Us3,
encodesanovel protein kinase and have characterized the cognate amino acid sequence which is phosphorylated by thisenzyme.Thisreportidentifiesanapparently essential viral phosphoprotein whose posttranslationalprocessing
involves the viral protein kinase.Analyses of viral proteins phosphorylated in thecourseof productiveinfection
revealed a phosphoprotein whose mobility was viral protein kinase and serotype dependent. Thus, the
corresponding HSV-1 and HSV-2 phosphoproteins differ in their electrophoretic mobilities, and the
phos-phoproteinspecifiedby theHSV-1mutantdeletedin
Us3
(R7041) differs fromthatofthecorrespondingHSV-1 andHSV-2proteins. Analyses of HSV-1x HSV-2 recombinantsmapped the phosphoprotein between 0.42 and0.47map units onthe prototype HSV-1 DNA map. Within this region, the UL34 open reading frame was
predictedtoencode aprotein of appropriate molecular weight which would also contain theconsensustarget sitefor phosphorylationby theviral protein kinaseaspreviouslydefined with synthetic peptides. Replacement
of thenativeUL34genewithaUL34genetaggedwitha17-amino-acid epitopefromthea4proteinidentified
thisgene asencoding the phosphoprotein. Finally,mutagenesis of the predicted phosphorylation siteonUL34
inthe viralgenome, andspecifically the substitution of threonineorserine with alanine inthe productofthe
UL34gene,yielded phosphoproteins whoseelectrophoretic mobilities couldnot bedifferentiatedfrom that of the
Us3-
mutant.We concludethatthe posttranslationalprocessing of the UL34geneproducttoits wild-typephenotyperequires the participation oftheviralproteinkinase. While the viral protein kinase isnotessential for viralreplication incells in culture, the UL34geneproduct itselfmaynot bedispensable.
The virusescomprisingthe subfamilyAlphaherpesvirinae of theHerpesviridae familyincludeherpessimplexviruses 1 and 2(HSV-1 and HSV-2), varicella-zoster virus, and swine herpesvirus 1 (pseudorabies virus [PRV]). Among thegenes
conserved by members of this virus subfamily isone
speci-fying a protein kinase (PK) (10, 21, 22). The PKs of both
HSV-1 and PRV have been purified and assayed by using protamine as a suitable artificial substrate (9, 29, 30). The PRV PK is a dimer consisting of two 38,000-molecular-weight subunits, whereas the HSV-1 enzyme is a dimer
consistingoftwo68,000-molecular-weight subunits (15, 29, 39). The purified PK requires no effectors, and it transfers phosphate from ATP,butnotGTP, tothe serylorthreonyl
residues ofbasic, butnotacidic,synthetic peptides (13).The studies on synthetic oligopeptides have suggested that the
phosphorylation site recognized by the viral PK has the
consensussequence
(R).X(S/T)YY,
wheren 2 3,R = Arg,X prefers Arg, Ala, Val, Pro, or Ser, Y shares the same
preferenceexceptthat acidic residuesorprolineis unaccept-able, and(S/T)isthe targetresidue, being either SerorThr (14, 28). Inaddition, the kinase iscapableof autophosphor-ylation invitro, althoughitsautophosphorylation in the cell has notbeenobserved (9, 29).
On the basis of the nucleic acid sequence of HSV-1, McGeoch and Davison (21) suggestedthat theopenreading frame designated as
Us3
may encode a PK. It wassubse-quently shown that (i) deletion of the
Us3
open readingframe in therecombinant virus R7041resulted in the loss of
*Corresponding author.
the novelPKactivity from lysates of infected cellswhereas restoration of the deleted sequences restored the activity (31)and(ii) antibodyraised againstasynthetic
eight-amino-acid C-terminal
Us3
oligopeptide conjugated to bovine se-rumalbuminreacted withpurifiedpreparationsof the HSV-1PK(9).
Interest in theHSV PK activity stemsfrom the observa-tion that while retroviruses arecapable oftransducinghost PKswhichmayactasoncogenes(3, 6, 12, 34), virus-specific
PKs are rare. In addition to the PK conserved among
members of the subfamilyAlphaherpesvirinae, PK activity has been ascribedtothetranscriptionaltransactivator hbx of theHepadnaviridae family (18, 35, 38). Comparisonsof the
sequences of HSV openreading frame UL13, the Epstein-Barr virus open reading frame BGLF4, and the varicella-zostervirusopenreadingframe 47 withthose of known PKs suggest that these genes may alsoencode novel PKs (36). The function of the HSV-1 PK encoded by
Us3
is notknown. To determine itsfunction,it is necessarytoidentify its natural substrates in the infected cell. To this end, we
have compared the 32P-labeled phosphoprotein profile of
electrophoretically separated polypeptides from cells in-fected with the recombinant PK- Us3 virus R7041 with those from cells infected either with its parent virus, HSV-1(F), orwithavirus inwhich the deleted
Us3
sequencesofR7041 have been restored. These studiesledtothe
observa-tion that in cells infected with the PK- mutant, a slower-migratingphosphoprotein replacedawild-type
phosphopro-tein. To determine whether the two phosphoproteins were
genetically related, we first mapped the approximate loca-tion of thegene specifying thewild-type phosphoprotein to
5757 0022-538X/91/115757-08$02.00/0
Copyright C) 1991, American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
A.
LJl l
IN J
I I
2mP aG 5 .1
3 1 > ,I<K I > I
4 r. ._;
* bUT~~]
K IP B481
69 357
6 K _ S
B.
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
TK UL34
I* -
-_
N F MOL
X ~- Be
.
.
4 3 3
_72Kb
between 0.42 and 0.47 map units on the viral genome by using a set of characterized HSV-1 x HSV-2
intertypic
recombinants. Withinthis region, the UL34 gene waspre-dicted to encode a
protein
of theapproximate
molecularweight of the mapped
phosphoprotein.
In addition, the product of thisgenewaspredicted
tocontainanaminoacidsequenceidenticaltothatofthe consensus
phosphorylation
siteoftheviralPKdetermined earlier (14, 20,28).Replace-mentwithintheviralgenomeof the native
UL34
genewithaUL34 genetaggedatitsamino terminuswitha17-amino-acid epitopefrom the a4
protein
led totheunambiguousdemon-stration that the UL34 gene encodes this
phosphoprotein.
Furthermore, mutations in the viral genome that led tosubstitution of either threonineorserinewith alaninewithin
the consensus
phosphorylation
sequence onthe UL34geneproduct
yielded
phosphoprotein profiles
which could notbedifferentiated
from those of the PK-Us3
virus. Theseresults indicate that the
posttranslational processing
of the UL34gene product to itswild-type phenotyperequires
theparticipation oftheviral PK.
MATERIALS ANDMETHODS
Cells and viruses. The isolation and
properties
ofHSV-1(F)
andHSV-2(G),theprototype HSV-1 and HSV-2strains used inthis laboratory, havebeen described elsewhere(8).
HSV-1(F)A305
and R7041 aregenetically
engineered
dele-tionmutants(25).HSV-1(F)A305
lacksthe500-bpBgIII-SacI
fragment in thethymidine kinase (tk)gene. R7041 lacks 860
bp ofthe coding sequences ofthe
Us3
open reading frame defined by the restriction endonuclease PstI-BamHI; thedeleted sequences encode the
predicted
amino acids 69 to 357ofUS3.
Theviruswasproduced
byrescueofthetkgeneofthe recombinant R7040 (17, 23) in which deletions were
introduced in both the tk and PK genes. Recombinants 5 (K) E UL34 B"
5 (K)'~~B
1 191 275
st27TK9H
D.
1 (K3FX
~UL34
S.R R R R T R R 8 R E
2 CGTCGC CGCCGAACC CGGCGGTCCCGGGAG
R R R R A R R S R E
3 CGT CGCCGCCGAGCGCGC
CG6
TCCCGG GAGR R R R T R R A R E
4 CGT CGC CGC CGA ACCCGGCG GCTCGGGAG S *
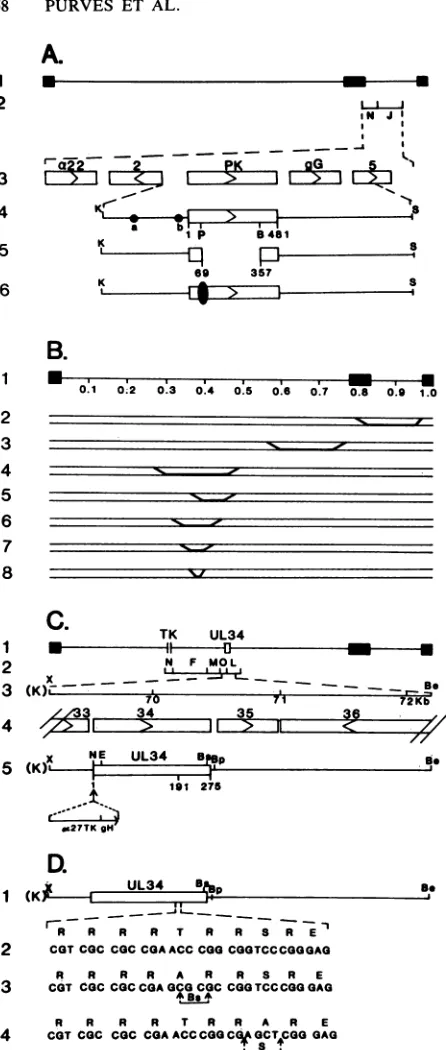
FIG. 1. Schematicrepresentationof theDNAsequence
arrange-ments in the genomes ofHSV-1(F) and the various recombinant viruses. (A) Line 1, the sequence arrangement of the HSV-1 genome. Thefilled rectangles representtheinvertedrepeats flanking
the unique sequences (represented by thin lines) of the long and short components. Line 2, the relevant HSV-1BamHI fragments whichcontain the openreadingframes for a22,Us2,glycoproteinG, thePK, and
Us5
asidentified in line 3. Line 4, the KpnI-SacI DNAfragment containing the 481-amino-acid open reading frame Us3
encoding thePK. Twoalterativetranscription initiationsitesfor the
US3mRNAareindicatedby thecircleslabeleda and b (33).Line 5,
the site ofthedeletion in R7041spanning from thePstIsitelocated
atamino acid69 to the BamHI site located at amino acid 357 of the PK.Line6, the DNAarrangementofR7307, arecombinantvirus in
whichahumanCMVepitope(indicatedby thesolid oval) isinserted into the unique PstI site located atamino acid 69 ofthe protein
kinase. (B) DNA sequence arrangements of HSV-1 x HSV-2
intertypic recombinants. Line1, thegenome arrangementofHSV
divided into viral map units. Lines 2 to 8 indicate the DNA arrangements of the intertypic recombinants R7015, RS1G25, RHIG7, RHIG8, RHIG13, RHIG44, and RHIG48,respectively.The
boldfacedline segmentsidentifyHSV-2sequencespresentinthese
[image:2.612.77.300.60.585.2]genomes, with theapproximatecrossover points falling within the indicated diagonal regions. (C) Arrangement ofgenesbetween0.42 and 0.47 map units and position of the UL34 gene on the viral
genome.Line1,the DNAsequencearrangementofHSV-1(F)A305, containinga500-bpdeletion in thetkgene.ThepositionoftheUL34
geneis indicatedby the openbox. Line2, relevant HSV-1 EcoRI fragments. Line 3, theXbaI-BstEII fragment containing the UL34
openreadingframe usedinconstruction of therecombinant viruses. Line4, theopenreading framesencoded within this fragment. Line 5, the DNAarrangementofrecombinant virus R7309,which harbors an a27-tkgene inserted at the NcoI site located at the initiating methionine codon of the UL34 gene such that the UL34 gene is expressed and regulated by the gH promoter sequences. (D)
Se-quence arrangement at the consensus phosphorylation site of the UL34protein. Line 1, wild-type DNAsequencearrangement. Line 2, DNA and aminoacidsequencesbetweenamino acids 191 and 200
of thewild-typeUL34protein. Line 3, DNAarrangementof recom-binantvirusR7310,inwhich thethreonineresidue has beenmutated
toanalanine residuecoincident with the introduction ofaBssHII
site. Line 4, DNA arrangement ofrecombinant virus R7311, in which the serine residue hasbeenmutatedtoanalanine,coincident withtheintroduction ofaSacl site. N, NcoI; K,KpnI;P, PstI;B, BamHI; E,EcoRI; X, XbaI; Be, BstEII; Bs, BssHII; Bp, BspEI. Restriction sites showninparentheses arederived from polylinker
sequences.
1 2
1
2 3 4 5 6 7 8
1 2 3
4
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Genotypes of genetically engineeredvirusesa
Virus Genotype
HSV-1(F)... PK+ UL34+
R7041... PK- UL34+
R7306... PK+ UL34+
R7307... PK(CMVtag)+ UL34+
R7310... PK+ UL34Ala-195
R7311... PK+ UL34Ala-198
R7314 ...PK+
UL34a4tag
aDerivation of the viruses is described in Materials and Methods.
R7015, RH1G7, RH1G8, RH1G13, RH1G44, and RH1G48 werepreviously described (1, 4). Recombinant RS1G25 was
derived by Conley and Roizman (5) by marker rescue of a mutant mapping at the terminus of the L component. The crossover maps of all HSV-1 x HSV-2 recombinants are
shownin Fig. 1. All virus stocks were titered on Vero cells. Construction of recombinant viruses. The recombinant
viruses constructed for this study and their genotypes are shown in Table 1. In recombinant virus R7306, the se-quencesdeletedin R7041 were restored by cotransfection of rabbit skin cells with intact R7041 DNA and plasmid pRB3446, which carries the HSV-1 4.89-kb Sacl fragment
containingtheentire
Us3
genewith flanking sequences. The progeny ofthis transfection was plated on Vero cells, andindividual plaques were screened for the restoration of the BamHINfragment. Inrecombinant virus R7307, the R7041
deletionwasrepaired witha
Us3
gene(pRB4274)containing a20-amino-acid epitope ofhuman cytomegalovirus (CMV)(16)inserted in frameatnucleotide 69 of thecoding sequence
ofthe
Us3
gene.To construct pRB4274, the 4.89-kb Sacl fragment from pRB3446 was cloned into the Sacl site ofpGEM7Zf+ to generate pRB4269. pRB4269 DNA was cleaved with KpnI
and religated to yield pRB4173; the KpnI collapse deleted
thesequencesfromtheKpnIsite in thepolylinker to thesite located at position -435 relative to the initiation codon of the
US3
gene and ensured that the PstI site located atposition +207 ofthe
Us3
coding sequence wouldprovideaunique site into which a 70-bp oligonucleotide encoding a
20-amino-acid CMV epitope contained within a 10-base
linker sequence could be inserted in-frame with the
Us3
coding
sequences, thus generating pRB4274. DNAsex-tracted fromplaque isolates oftherecombinant viruseswere
screened for boththe restoration oftheBamHINfragment and the presence oftheCMV epitope by Southern
blotting
(37)withthe CMVoligonucleotide (Fig. 2B andC).Toconstructrecombinantviruses withmutations or
dele-tionsin the UL34gene,the oa27-tk genedescribedelsewhere
(17)wasinsertedbetween theUL33 andUL34 openreading frames to generate R7309. This recombinant was
con-structed by cotransfection of rabbit skin cells with intact
HSV-1(F)A305
DNAandpRB4165,whichcontainsanoa27-tk gene inserted between the UL33 andUL34
genes. TK+ progeny virusfromthis transfection wereselectedon 143tk-cells underhypoxanthine-aminopterine-thymidine
and fur-ther plaque purifiedon Vero cells as describedpreviously
(25). pRB4165 was constructed byinserting
a chimeric a27-tk geneinto theNcoI site locatedattheATGofUL34inplasmidpRB4247sothat the initiation codonof the genewas
restored and theUL34genewasdriven bythegH promoter sequences located within the tk
coding
sequences(20).
pRB4247 consists of the3,066-bp
XbaI-BstEIIfragment
subcloned from the EcoRV E
fragment
cloned into pGEM3Z. Recombinant viruses were screenedby
detecting
-I! m < =
'I
-cr, r.c4 r-
r-7- 0
> - o
r.. n r- r
C) a:- 0c
29
3-3 4
1-5o @ _
4-6N - - *G
A B C D
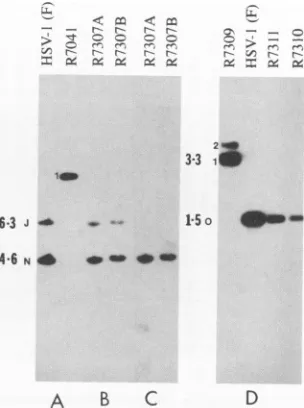
FIG. 2. Autoradiographicimages ofBamHIorEcoRIdigestsof HSV-1 wild-type andrecombinantmutant viral DNAs. Viral DNA
digests were electrophoretically separated on 0.8% agarose gels, transferredtonitrocellulose membranes, and hybridizedwith
spe-cific radiolabeled probes. The viral DNA in panels A and B was
digestedwithBamHIandprobedwithradiolabeledpRB3446,which contains the viralPKgene.BamHINand Jbandsareindicated.The
banddesignated1resultedfrom the deletionofUs3gene sequences
(Fig. 1A, line 5), which caused the fusion ofBamHI-J with the remainder ofBamHI-N. The viral DNA shown in panel C was
digested with BamHIandprobedwithradiolabeled oligonucleotide specifyingthe humanCMVepitope. The viralDNAshown inpanel
D wasdigested withEcoRI and probed witharadiolabeled HSV-1(F) EcoRI 0 fragment (identified by the letter 0). The band designated1in theR7309digestswasgenerated bytheinsertion of
achimeric ax27-tkgene intothe EcoRI 0 fragment. Aband desig-nated 2 was generated as a result of the inversion of the DNA
fragment spanning the native and inserted tk sequences. The ge-nomes containing the inversions were present in relatively small
amounts.Theintroduction ofmutantUL34genesinvirusesR7310
andR7311 restored the1.5-kb EcoRI0fragmentandeliminatedthe
3.3-kbfragmentpresentinR7309.
theincreasein size of the EcoRI 0 bandfrom 1.5 to 3.3 kb
resulting fromthe insertionof the a27-tk gene(Fig. 2D). Recombinantvirus R7314 contains a UL34 gene which is
tagged at its amino terminus with a 17-amino-acid
epitope
fromtheHSV-1 oa4proteininplace ofthenativeUL34gene. R7314wasconstructedbycotransfection of rabbit skincellswith intactR7309viral DNA andpRB4277,whichconsistsof a49-bp oligonucleotide
encoding
the (a4epitope
cloned into the NcoI site located at theinitiating
methionine codon of the UL34 gene in pRB4247. TK- progeny virus from this transfectionwereselectedon 143tk-cells in the presence ofbromodeoxyuridine. Recombinant viruses were
plaque
pu-rifiedonVerocellsandscreenedbydetecting
therestoration of the EcoRI 0 bandby
theprocedure
ofSouthern(37).
Recombinant viruses R7310 and R7311 each contain a
UL34 gene in which the
coding
sequence hasbeen alteredtoincorporateaspecific
single
aminoacidchange
(Fig.
1D).
In recombinant virus R7310, the threonine residue at amino acid 195 has beenchanged
to analanine,
whereasin recom-binant virusR7311,theserineresidueatamino acid 198 has beenchangedtoanalanine. Theseviruseswereconstructed asfollows.A1,249-bp fragment ofHSV-1(F)DNAcontain-ing UL34 spanning from the XbaI site to the BspEI site
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.62.300.91.173.2]-2 - r-- I- , . r
r--- r---
ClJ
:/
-_o
B
- -.e = ., .j F--J-. C
.-l.
--f
a~~~~, I767i___\trI
t94
*~~~~~~~~~~~f-6*|- 7
*s^sA Av ^ ^ ^ 30
II
[image:4.612.133.489.82.266.2]-20
FIG. 3. Autoradiographic imagesofelectrophoretically separated32P-labeled infected celllysatesfromcells infected withwild-typeand recombinantHSV-1. All of the celllysatesineachpanelwereelectrophoretically separated concurrently. Inpanel A,thetriangleidentifies thewild-type HSV-1 UL34 protein,thesquareidentifiesthe HSV-2UL34 protein,and the circlerepresentstheUL34 phosphoproteininlysates
ofcells infected with the R7041 (PK-Us3-)virus. Inotherpanels,the circle also identifies theUL34phosphoproteinswhichcomigratewith
the correspondingphosphoproteinfrom R7041-infected cells(e.g.,theUL34phosphoproteinpresentinlysatesofcellsinfected with theR7015
virus).
within EcoRV E fragment was excised as a KpnI-BspEI
fragment from pRB4247, blunt ended with T4 polymerase, andsubcloned into the SmaI siteofpGEM3Zf+, generating pRB4164. With this construct, site-directed mutagenesis using syntheticoligonucleotides was performedto generate pRB4233, inwhichthethreonineresiduewasmutatedtoan
alanine residue coincident with the generation ofa unique
BssHII restriction site. A similar procedure was used to
generatepRB4234, inwhich the serine residuewasmutated
to an alanine residue coincident with the generation of a
unique SacI restriction site. Each of these mutated UL34
genes was excised as a 682-bp NcoI-HincII fragment and subcloned back into pRB4247 in order to restore flanking
sequences necessary for recombination into the viral
ge-nome, generating pRB4236, used in the construction of
R7310, the UL34 threoninemutantvirus,andpRB4237,used intheconstruction ofR7311, theUL34 serine mutantvirus. pRB4236 and pRB4237 were each cotransfected on rabbit skin cells withintact R7309 viral DNA. TK- progeny virus
were selected on 143tk- cells as previously described (26).
Individual plaques were purified on Vero cells, and viral
DNA was screened for the presence of the introduced
mutations by amplifying a 600-bp fragment containing a
portionofthe UL34 gene by thepolymerase chain reaction andchecking this amplified fragment for thepresenceofthe
genetically engineered restriction sites.
Preparation of32P-labeled infected cell lysates. BHK-C13 cellsgrownin25-cm2flaskswereinfected with10PFUofthe indicated viruspercellin medium199V.After 13 h, the cells were preincubated in Eagle's minimal essential medium
made without phosphate and supplemented with 1% dia-lyzed phosphate-free calfserumfor 1 h. Cellswere labeled
with 100 to 200 ,uCi of
32Pi
(carrier free; New England Nuclear) in a final volume of3 ml for 4 h. Because of thedifficulty of obtaining revertant-free high-titer stocks of viruses R7310 and R7311, cells were infected with
single-plaque isolates of these viruses and after36hwerelabeled
with
32Pi
as described above. Cells were rinsed once andscraped into 1 mlof ice-cold PBS-A, subjected to centrifu-gation inamicrocentrifugefor 5minat4°C,andresuspended
in350 ,ul of PBS-A containing 0.1 mM tosylsulfonyl phenyl-alanyl chloromethyl ketone, 0.1 mM tosylsulfonyl lysyl chloromethyl ketone, 1.0%(vol/vol) Nonidet40, and 1.0% (wt/vol) sodium deoxycholate. Lysateswere sonicated and frozen inaliquots at -70°C.
Polyacrylamide gel electrophoresis. Infected cell lysates containing approximately 50 ,ug of protein were electro-phoreticallyseparated inpolyacrylamide (10%) gels contain-ing 0.1% sodium dodecyl sulfate. The separated polypep-tides were electrically transferred tonitrocellulose, stained with amido black, and subjected to autoradiography on
Kodak X-Omat film for 12 h.
RESULTS
The phosphoprotein profile of R7041 differs from that of HSV-1(F). Toidentify protein substrates of the viral PK in infected cells, replicate cultures of BHK-C13 cells were
infectedwith eitherHSV-1(F) or R7041. At 13 h postinfec-tion,the cellswereincubatedinmedium containing32Pifor 4 h. The cellswerethenharvested, solubilized,and electro-phoretically separated in denaturing polyacrylamide gels as
described in Materials and Methods. Comparison of the 32P-labeledphosphoprotein profilesofHSV-1(F)andR7041 (Fig. 3A) showed that a novel phosphoprotein of approxi-mately 33,000 in apparent molecular weight replaced a
wild-type HSV-1(F) labeled phosphoprotein of approxi-mately 30,000 in apparent molecular weight in the R7041-infectedcell lysates.
To determine whether the observed differences in the phosphoprotein profiles of the HSV-1(F) and the R7041-infected celllysateswereduetothe deletion of the
Us3
geneproductin R7041 and not tosome other unidentified
muta-tion,twoadditionalrecombinantviruses, R7306andR7307,
wereconstructedasdescribed inMaterialsandMethods.In
recombinantR7306, thedeletedsequences of
Us3
in R7041were marker rescued with the authentic gene. In mutant
R7307,thedeletedsequenceswererepairedwitha
Us3
genecontaininganadditional 20amino acidsinserted in frameat
amino acid 69 and expressing a human CMV epitope.
- -
r-S~
~
A
on November 10, 2019 by guest
http://jvi.asm.org/
Comparison of the 32P-labeled phosphoprotein profiles of lysates from replicate cell cultures infected with R7306 and HSV-1(F), respectively (Fig. 3B), indicate that the two are indistinguishable and therefore the differences in the phos-phoprotein profiles of HSV-1(F)- and R7041-infected cells reflect the deletion of the
US3
gene in R7041.The phosphoprotein profiles of lysates of replicate cell cultures infected with HSV-1(F), R7041, R7306, or two independent isolates of R7307 (R7307A and R7307B, respec-tively) (Fig. 3B) show that (i) in the infected cell lysates of R7306, the phosphoprotein profile was that of the wild-type HSV-1(F) and (ii) thephosphoprotein profile of the lysates of cells infected with R7307 contained both the 33,000-appar-ent-molecular-weight phosphoprotein characteristic of the R7041 deletion virus and the 30,000-apparent-molecular-weight phosphoprotein characteristic of thewild-type HSV-l(F)-infected cells. We conclude from these experiments that (i) the 30,000- and the33,000-apparent-molecular-weight
phosphoproteins are structurally related inasmuch as repair of the PK gene resulted in the replacement of the 33,000-apparent-molecular-weight phosphoprotein with the faster-migrating wild-type species and (ii) the twoproteins maybe related in that both phosphoproteins are present in cell lysates infected with R7307 virus. Thus, the 33,000-appar-ent-molecular-weight species may be either a precursor of
the 30,000-molecular-weight protein or a by-product of the phosphorylation of acommon precursor by an infected-cell
PK. In the case of the R7307 virus, insertion of a sequence encoding the 20-amino-acid epitope into the
Us3
gene may have generated an impaired PK incapable ofrapid process-ing of the33,000-apparent-molecular-weightphosphoproteininto the wild-type phosphoprotein. It is noteworthythat the HSV-1 x HSV-2 intertypicrecombinantvirus R7015yielded
a similarpattern. This virus consists of theHSV-1(F)unique
long region, which encodes the UL34 gene, and the HSV-2(G) unique short region, which encodesthe
Us3
gene. As seen in Fig. 3B, the phosphoproteinprofiles ofcells infected with R7015 exhibit both the 30,000- andthe33,000-molecu-lar-weight forms of the UL34 protein. We conclude either
that in the recombinant the PK geneis defective orthat the PK and UL34 genes may havecoevolved within serotypesto
maximally interact with each otherandtherefore the recom-binant PK is less efficient in phosphorylating its HSV-1 substrate. Differentiation between these hypotheses must
awaitfurther study of the HSV-2 PK ofthe R7015 virus. HSV-1 x HSV-2 intertypic recombinants map the gene specifying the 30,000-apparent-molecular-weight wild-type
phosphoprotein to a region of the genome located between 0.42 and 0.47 map units. Earlierstudies have shown that HSV-1 and HSV-2fortuitouslydifferwith respecttothe
electropho-retic mobilities of many of theirproteinsin
denaturing gels
(24). To determine whether the HSV-1
30,000-apparent-molecular-weight phosphoprotein differs from that of its HSV-2(G) counterpart, we compared the
electrophoretic
profiles of HSV-1(F)- and of
HSV-2(G)-infected-cell
poly-peptides harvested 17 hpostinfection. As illustrated in
Fig.
3A, thecorrespondingHSV-2(G)
phosphoprotein
possessed
an electrophoretic mobility distinctly lower than that of either the 30,000- or
33,000-apparent-molecular-weight
HSV-1 phosphoprotein. The difference in the
mobility
of HSV-1 andHSV-2phosphoproteins was inaccord withour expectation that thesephosphoproteins
werespecified
by
the virus and suggested that the gene
specifying
themmight
be mapped by analysis of HSV-1 x HSV-2 recombinants.Theanalyses of HSV-1x HSV-2recombinantsweredone in two stages. The location of the gene
specifying
the30,000-apparent-molecular-weight
phosphoprotein
was firstapproximated
by comparing
32P-labeledphosphoprotein
pro-files oflysates
from cells infected with those of the threeintertypic
recombinants, RH1G7,
RS1G25,
and R7015(Fig.
3B).
Ofthese,
thelower-mobility
HSV-2 form ofthephos-phoprotein
was observedonly
inlysates
of cells infected with RH1G7. On the basisofthisresult,
thelocation ofthephosphoprotein
was furthermapped by using
four moreintertypic recombinants, i.e., RH1G8,
RH1G13,
RH1G44,
and RH1G48. As shown in
Fig.
3C,
thephosphoprotein
with the characteristic slowermobility
ofHSV-2(G)
waspresent
in
lysates
of cells infected with RH1G8. Theseresults,
interpreted
withrespecttothecrossoverpoints
illustratedinFig.
1B,
indicate that the genesspecifying
the30,000-apparent-molecular-weight HSV-1(F)
phosphoprotein
andits
HSV-2(G)
counterpart map between 0.42 and 0.47 map units on the viral genome.The
UL34
openreading
frame encodes thephosphoprotein
which
displays
alteredelectrophoretic mobility
in thepresence ofthe viral PK.Theregion
between 0.42 and 0.47mapunitsincludes part ofthe open
reading
frame forUL30,
the viral DNApolymerase,
and the entire openreading
frames forgenes
UL31
toUL35.
Ofthese genes,only
UL31
andUL34
encode
polypeptides
withpredicted
molecularweights
in the range ofthat of themapped
phosphoprotein. Inspection
of thepredicted
aminoacidsequenceoftheUL34
geneproduct
revealed a
potential
viral PKtarget
site(Arg-Arg-Arg-Arg-Thr-Arg-Arg-Ser-Arg-Glu)
located betweenaminoacids191and 200. Within this sequence, both the
serine
and thethreoninewere
potential
target
residuesofthe viralPK. Todeterminewhether the
UL34
geneencoded thephosphopro-tein,
we constructedtherecombinant virusR7314,
in which thenativeUL34
genewasreplaced
withaUL34
genetagged
atitsaminoterminuswitha17-amino-acid
epitope
from theHSV-1 a4
protein
asdiagrammed
inFig.
4A. AsillustratedinFig. 4B,
thephosphoprotein
present
in the twoindepen-dently
derived R7314-infected celllysates
possessed
amo-bility
distinctly
lower than that of either the30,000-
or33,000-apparent-molecular-weight
HSV-1phosphoprotein
foundineither
HSV-1(F)-
orR7041-infected celllysates.
Inaddition,
thislower-mobility
phosphoprotein
reacted withmonoclonal
antibody
H943,
directedagainst
the17-amino-acid
epitope
present
onthetagged
UL34
geneproduct
(11).
These results
positively
identified thephosphoprotein
asbeing
encodedby
theUL34
gene.Itisimportant
tonotethatthe decreasein
electrophoretic
mobility
ofthetagged
protein
expressed by
R7314 wasgreater
than thatpredicted
by
theinsertion ofthe17-amino-acid
epitope
into the30,000-appar-ent-molecular-weight
phosphoprotein specified
by
HSV-1(F)
and
exactly
thatpredicted
by
theinsertion oftheepitope
intothe
33,000-apparent-molecular-weight
phosphoprotein
spec-ified
by
theUs3-
recombinantR7041. These resultssuggest
that the presenceof theamino-terminala4
epitope
tagontheUL34
protein
mayprevent
itfromconverting
to thehigher-mobility
species
seen in thewild-type
virus in the presenceof the PK.
Site-specific
mutagenesis
of theUL34
openreading
frameindicates that the
Us3
PK is involved inposttranslational
processing
oftheUL34
geneproduct.
To determine whethersingle
amino acidchanges
within thepredicted
consensus target site ofthe viral PK could affect themigration
oftheUL34
phosphoprotein,
we constructed recombinant virusesR7310 and
R7311,
each of which containedsite-specific
mutations in the
UL34
gene within thisregion.
Inrecombi-nantvirus
R7310,
thethreonineresidue of thetarget
sitewaschanged
toan alanineresidue,
whereas inR7311,
theserine
residue was
changed
to an alanine residue. The32P-labeled
phosphoprotein profiles
of thelysates
ofcells infected withon November 10, 2019 by guest
http://jvi.asm.org/
Be 0 fL.
CI_
Io
uO>
CAr;
CAAK
---1 191 275 aThg Insertion
-_1- -
-r-- r- V) V)r
P W. = m $Mrx1O3
METAsPGLuTYRAspASPALAALAASPALAALAGLYASPARGALAPROGLYrMET CATGGACGAGTACGACGACGCAGCCGACGCCGCCGGCGACCGGGCCCCGGG**** ****CTGCTCATGCTGCTGCGTCGGCTGCGGCGGCCGCTGGCCCGGGGCCCGTAC
ffiff c < X
X10a I CC I CC 9C E :
'a:Ia
CW CVM. b I ML ..,
94-67
oo11,0 -UL34
30-20- EJAI&
[image:6.612.77.310.70.353.2]AUTORADIOCRAM IMMUNOSLO
FIG. 4. Autoradiographic and photographic images of
electro-phoretically separated phosphoproteins for identification of the mappedphosphoproteinastheproduct of the UL34gene.(A) DNA
sequence arrangement of the UL34 gene present in recombinant virus R7314, in whicha17-amino-acid epitope from the HSV-1 a4 proteinwasinserted atthe aminoterminus of the UL34gene. Be, BstEII; Bp, BspEI; Bs, BssHII; E, EcoRI; K, KpnI; N, NcoI; X, XbaI. Restriction sites shown in parentheses are derived from polylinker sequences. (B) 32P-labeled phosphoprotein profile and corresponding immunoblot of lysates of cells infected with the indicatedviruses, electrophoretically separated in denaturing gels, and transferred to nitrocellulose. The nitrocellulose was reacted with monoclonal antibody H943 to the 17-amino-acid a4 epitope. The autoradiogram illustrates the threeelectrophoretically distinct
phosphoprotein species present in 2(G)- R7041-, and
HSV-1(F)-infected cell lysates (indicated by squares, filled circles, and triangles, respectively), as well as the the more slowly migrating
phosphoproteinpresentin R7314infected cell lysates (indicated by
opencircles). This phosphoprotein reactswith the a4monoclonal antibody. Note that at 17 h postinfection, the amount of UL34
protein exceeds theamountof a4 protein present in the lysates of theinfected cells.
eitherof these viruses aredisplayed in Fig.5. In each case,
the 30,000-apparent-molecular-weight wild-type phos-phoprotein was replaced by a phosphoprotein whose elec-trophoretic mobilitycannotbe differentiatedfrom that of the
33,000-apparent-molecular-weight phosphoprotein present in lysates of cells infected with the PK- Us3- R7041
recombinant. The observations that the mutations confer
upon the product of the UL34 gene in a PK' virus the
phenotype of thePK-
Us3-
virus indicate thattheviralPKisrequired for the posttranslational processing of the UL34
geneproduct.
DISCUSSION
Thestudies described in thisreportshow the following. (i) In cells infected with the PK- virus, R7041, a wild-type
phosphoprotein withanapparentmolecular weight of30,000
[image:6.612.391.508.81.302.2]41 -20
FIG. 5. Autoradiographic images of electrophoretically
sepa-rated 32P-labeled infected cell lysates from cells infected with
recombinantviruses R7310 andR7311,which harborspecific
muta-tions within theputative targetsite for the viral PK onthe UL34 protein. Labeled lysates were prepared from cells infected with
single-plaque isolates of each virus. The squares, circles, and
triangles represent the HSV-2(G), R7041, and HSV-1(F) forms of
theUL34geneproduct,respectively.
isreplaced byaphosphoprotein withanapparentmolecular weight of 33,000. (ii) The 30,000-molecular-weight phos-phoprotein maps tothe open reading frame UL34. (iii)The protein specified by UL34 is predicted to contain an amino
acid sequencewhich correspondsto theidealized substrate ofthe viral PK determined previously. (iv)Direct evidence of the association ofthe 33,000-apparent-molecular-weight
phosphoprotein with the UL34 gene product and of the
involvement of the viral PK with the replacement of the
33,000-apparent-molecular-weight phosphoprotein with the 30,000-molecular-weight phosphoprotein is based on the
experimental evidence that mutagenesis of thethreonineor
serine within the sequence corresponding to the idealized substrate of the viral PK results in the expression of the
33,000-apparent-molecular-weight phosphoprotein in place of thefaster-migrating wild-typephosphoprotein. These data indicate that the posttranslational processing of the UL34
geneproduct involves the function of the viral PK.
The results presented in this report raise four significant issues: (i) the relationship between the twoproducts of the UL34 open reading frame, i.e., the 33,000- and the 30,000-molecular-weight phosphoproteins, (ii) the localization and functionof theproduct of the UL34gene,(iii)therole of the UL34 gene product in the reproductive cycle, and (iv) the role ofthe
Us3
gene encoding the PK in the reproductivecycle ofthe virus.
Relationship between the 33,000- and 30,000-apparent-molecular-weight phosphoproteinproducts of the UL34 gene.
Severalobservationsreported in thispaper aresignificant. (i)
Theproduct of the UL34openreading frame is
phosphory-latedintheabsenceof the viral PK encoded by the
Us3
openreadingframe. Thisobservation implies that the product of theUL34 open reading frame is, or canbe,phosphorylated
byanother PKpresentintheinfectedcell. (ii) If the viral PK
B.
-94
-67
-43
-30
on November 10, 2019 by guest
http://jvi.asm.org/
phosphorylates the UL34 phosphoprotein at the viral PK phosphorylation consensus site, then the observation that replacement of either the serine or threonineinthepredicted
phosphorylation site led to accumulation of the 33,000-apparent-molecular-weight phosphoprotein rather than of the wild-type 30,000-molecular-weight phosphoprotein indi-cates that the sites of phosphorylation of the 33,000-andthe 30,000-apparent-molecular-weight phosphoproteins are not identical. (iii) Because both the 33,000- and the 30,000-apparent-molecular-weight proteins arephosphorylated,itis not known whether the 33,000-molecular-weight phos-phoprotein is a precursor or a by-product of the wild-type 30,000-apparent-molecular-weight phosphoprotein encoded by UL34. Thus, the UL34 translation product could be phosphorylated by a cellular PK to yield the
33,000-appar-ent-molecular-weight protein, which in turn serves as a substrate for the viral PK, the product of the
Us3
open reading frame. Alternatively, thetranslation product of the UL34 gene is phosphorylated by eitherthe viral orcellularPK, but not at identical sites. Either phosphorylation en-ables the function of the UL34 gene product.
In either case, the increase inelectrophoretic mobility of wild-type UL34 protein may reflect either a change in the charge and shape of the phosphoprotein or a proteolytic
cleavage event induced by theviral PK. The latter hypoth-esis is formally supported bythepredicted structure of the protein described below,althoughdefinitive evidence is still lacking.
Localizationandfunctionof theUL34 gene product during the replicative cycle of thevirus. TheUL34 gene product has been reported to be a virion component, butthe data upon which this isbased have notbeenreportedorcited (19, 20). Analysis of its predictedamino acidsequence indicates that the N-terminal domainof theUL34 polypeptidehas proper-ties similartothose of signal sequences, lendingsupport to the hypothesesthat theprotein istargeted to membranesand
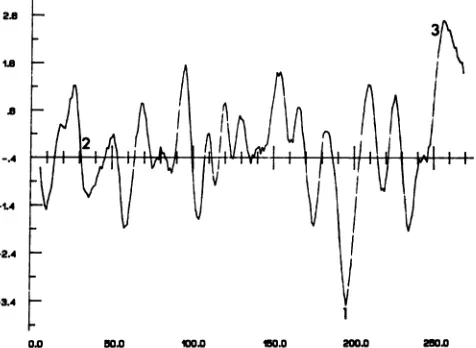
that under certain circumstances this sequence might be cleaved (27). Hydrophobicity analyses (Fig. 6) predict that the protein containsanextremelyhydrophilic domain which
coincides precisely with the amino acid sequence corre-sponding totheidealizedsubstrate ofphosphorylationbythe viral PK. Inaddition, the UL34 polypeptide is predicted to have a remarkablyhydrophobicCterminus,whichishighly
conserved (2, 7). Thishydrophobic sequence could conceiv-ably anchortheUL34protein inmembranes. The predicted
site of phosphorylation, surprisingly, is not as well con-servedand maynot be presentin eitherVZVor EBV.
Role oftheproductof the UL34 open reading framein the viralreproductivecycle.Whereasthe viral PK isdispensable,
numerous attempts to delete the UL34 gene have not been successful(32).Additional support ofthehypothesisthat the UL34 gene is not dispensable rests on the observation that the recombinants with the single amino acid
substitutions,
i.e., R7310 and R7311, areimpaired intheirgrowth proper-ties. Thesemutants grow so slowly that revertants
capable
of normal growth rates frequently arise, which
quickly
outgrow the mutant viruses. The revertants exhibit UL34
geneproductsofwild-type phenotype,i.e.,characterized
by
afasterelectrophoretic mobilitycorresponding to
30,000
in apparentmolecularweight (data not shown).The necessary conclusions are that (i) UL34 plays a
significant, probably essential role in viral
replication,
(ii)
while theposttranslational modificationofUL34 mediated
by
the
Us3
PK is not essential for viral replication in cells inculture, mutations in the PK
phosphorylation
consensus sequencemayinterfere with viral replication, and(iii)
if thephosphorylationof the UL34 geneproductisessentialfor its
2.6
2S
-2.4
-3.4 1/V
[image:7.612.318.556.83.257.2]0.0 50.0 W 1 50.0 200.0 250.0
FIG. 6. Hydropathic analysis of the predicted amino acid se-quence forthe UL34 protein from HSV-1 strain 17. The hydropathic plot was obtained by using the algorithm of Kyte and Doolittle with a moving window of 7. The subsequent profile was smoothed for plotting by taking its average in a moving 5-residue window. The plot isconstructed such that points
lying
above the x axisrepresent regions ofabove-average hydrophobicity while points lying below the axis represent regions of above-average hydrophilicity. Posi-tionsof the putative kinase target site, the observed signal sequence consensus, and the extremely hydrophobic C-terminal tail are indicated by 1, 2, and 3, respectively.function, then either the 33,000- or the 30,000-apparent-molecular-weight phosphoprotein is essential for viral
repli-cation,
at least in cells in culture.Role of the PK in the replicative cycle of the
virus.
Our results demonstrate that replacement of the 33,000-apparent-molecular-weight phosphoprotein by the wild-type 30,000-molecular-weight phosphoprotein requires the participation of the viral PK, and in this sense the product of the UL34 gene is a target of the PK and the first such targetidentified to date. The possibility that additional targets exist are based on two observations. The first relates to the morphology of infected cells in culture. Specifically, the plaques formed by R7041 are characteristically small and consist largely of shriveled cells which tend to float off their solid substrate very early after infection, incontrast to cells infected by the wild-type parent.These properties are accentuated in cul-tures infected with high ratios of virus per cell. Marker rescue of R7041 results in the restoration of the wild-type properties. It is therefore highly significant that cells infected with recombinant viruses R7310 and R7311, in which thePK
target site on the UL34 gene has been mutated, while exhibiting a UL34 gene product with the decreased electro-phoretic mobility characteristic of the R7041 virus, do not display the highly distinctive plaque morphology of cells infected with the R7041 virus. It also noteworthy that while the viral PK gene is nonessential for the growth of HSV-1 in tissue culture (17), the R7041 virus displays decreased
neu-rovirulence
in mice when tested by intracranial injection (23). The search for additional targets of the viral PK gene is in progress.ACKNOWLEDGMENTS
We thank Lenore Pereira for the invaluable gift of monoclonal antibodies and David Barker forplasmid pRB4247.
These studies were aided by Public Health Service grants from the National Cancer Institute (CA47451) and the National Institute for
on November 10, 2019 by guest
http://jvi.asm.org/
Allergy and Infectious Diseases (AI124009 and A11588). D.S. is a predoctoraltrainee under Public Health grant GM7281.
ADDENDUM IN PROOF
After the proofs were received, we tested newly made antibody to UL34 protein. These studies indicated that, in cells infected with PK- virus, the UL34 protein is not phosphorylated, that is, its phosphorylation totallydepends on viral PK. These studies also indicated that the phos-phoprotein in the upper 32P-labeled band in autoradiograms ofelectrophoretically separated lysates of cells infected with PK- virus is not structurally related to the UL34 protein although it may be functionally related toit.
REFERENCES
1. Ackermann, M., R. Longnecker, B. Roizman, and L. Pereira. 1986. Identification, properties, and gene location of a novel glycoprotein specified by herpes simplex virus 1. Virology 150:207-220.
2. Baer, R., A. T. Bankier, M. D. Biggin, P. L. Deininger, P. J. Farrell, T. J. Gibson, G. Hatfull, G. S. Hudson, S. C. Satchwell, C.Seguin,P. S.Tufnell, and B. G. Barrell. 1984. DNA sequence
andexpressionof the B95-8 Epstein Barr virus genome. Nature
(London) 310:207-211.
3. Collett, M. S., and R. L. Erikson. 1978. Protein kinase
associ-ated withtheaviansarcoma virus src gene product. Proc. Natl.
Acad. Sci. USA 75:021-024.
4. Conley,A.J., D. M. Knipe, P. C.Jones, and B. Roizman. 1981.
Molecular geneticsofherpes simplex virus. VII.
Characteriza-tion of a temperature-sensitive mutant produced by in vitro
mutagenesisanddefectiveinDNA synthesis and accumulation
of-ypolypeptides. J. Virol. 37:191-206.
5. Conley, A. J., and B. Roizman. Unpublished results.
6. Courtneidge, S. A., and A. E. Smith. 1983. Polyoma virus
transforming protein associates with the product of the c-src
cellulargene. Nature(London) 303:435-437.
7. Davison, A. J., and J. E. Scott. 1986. The complete sequence of
varicella-zoster virus. J. Gen. Virol. 67:1759-1816.
8. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1968.
Characteri-zation of herpes simplexvirusstrains differing in their effects on
social behaviour ofinfectedcells. J. Gen. Virol. 2:357-364.
9. Frame, M. C., F. C. Purves, D. J.McGeoch, H. S. Marsden, and D. P. Leader. 1987. Identification of the herpes simplex virus
protein kinase as the product of the viral gene Us3. J. Gen.
Virol. 68:2699-2704.
10. Hanks, S. K., A. M. Quinn, and T. Hunter. 1988. The protein
kinasefamily: conserved feature and deduced phylogeny of the
catalyticdomains. Science 241:42-52.
11. Hubenthal-Voss, J., R. A. Houghton, L. Pereira, and B. Roiz-man. 1988. Mapping offunctional and antigenic domains of the a4protein ofherpes simplex virus. J. Virol. 62:454-462. 12. Hunter, T., and J. A. Cooper. 1986. Viral oncogenes and
tyrosine phosphorylations. Enzymes17:191-246.
13. Katan, M., W. S. Stevely, and D. P. Leader. 1985. Partial
purification and characterization of a new phosphoprotein ki-nasefrom cells infected with pseudorabies virus. Eur. J.
Bio-chem. 152:57-65.
14. Leader, D. P., A. Donella-Deana, F. Marchiori, F. C. Purves, and L. A. Pinna. 1991. Further definition of the substrate
specificityof thealphaherpesvirus protein kinase and
compari-son with protein kinases A and C; Biochim. Biophys. Acta
1091:426-431.
15. Leader, D. P., and F. C. Purves. 1988. Theherpesvirus protein
kinase: a new departure in protein phosphorylation? Trends
Biochem. Sci. 151:244-246.
16. Liu, F., and B. Roizman. 1991. The promoter, transcriptional unit,andcoding sequence of herpes simplex virus 1 family 35
proteinsarecontainedwithin and in frame with the UL26 open
reading frame.J. Virol.65:206-212.
17. Longnecker, R., and B. Roizman. 1987. Clustering of genes
dispensable for growth in culture in the S component of the
HSV-1genome. Science236:573-576.
18. Maguire, H. F., J. P. Hoeffler, and A. Siddiqui. 1991. HBV X
proteinaltersthe DNA binding specificity of CREB and ATF-2
byprotein-protein interactions. Science 252:842-844.
19. Marsden, H.S., N. D.Stow, V. G. Preston, M. C. Timbury,and N. M. Wilkie. 1978. Physical mapping of herpes simplex
virus-inducedpolypeptides. J. Virol. 28:624-642.
20. McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan, M. C. Frame,D.McNab,L.J. Perry, J.E.Scott,and P.Taylor. 1988. Thecomplete DNAsequence of the longuniqueregionin the genome of herpes simplex virus type 1. J. Gen. Virol. 69:1531-1574.
21. McGeoch, D. J., and A. J. Davison. 1986. Alphaherpesviruses
possess a genehomologous totheprotein kinasegenefamilyof eukaryotes andretroviruses. Nucleic Acids Res. 14:1765-1777.
22. McGeoch, D. J., A. Dolan, S. Donald, and F. J. Rixon. 1985.
Sequencedetermination andgenetic contentof the shortunique
region in the genome of herpes simplex virus type 1. J. Mol.
Biol. 181:1-13.
23. Meignier, B.,R.Longnecker, P. Mavromara-Nazos,A. E.Sears,
and B.Roizman. 1988. Virulence of andestablishment oflatency
by genetically engineered deletion mutants ofherpes simplex
virus 1. Virology 162:251-254.
24. Morse, L. S., L.Pereira, B. Roizman, and P.A, Schaffer. 1978. AnatomyofHSV DNA. XI. Mappingof viralgenesbyanalysis
of polypeptides and functions specified by HSV-1 x HSV-2 recombinants. J. Virol. 26:389-410.
25. Post, L. E., S.Mackem, and B. Roizman. 1981. Regulationofa genes of herpes simplex virus: expression of chimeric genes
produced by fusion of thymidine kinase witha gene promoters.
Cell 24:555-565.
26. Post, L. E., and B. Roizman. 1981. A generalized technique for the deletion of specific genes in large genomes: a gene 22 of HSV is not essential for growth. Cell 25:227-232.
27. Pugsley, A. P. Protein targeting, p. 45-168. Academic Press, San Diego, Calif.
28. Purves, F. C., A. Donelia-Deana, F. Marchiori, D. Leader P., and L. A.Pinna. 1986. The substrate specificity of the protein kinase induced in cells infected with herpesviruses: studies with syn-thetic substrates indicate structural requirements distinct from other protein kinases. Biochim. Biophys. Acta889:208-215.
29. Purves, F. C., M. Katan, and D. P. Leader. 1987. Complete
purification of the pseudorabies virus protein kinase. Eur. J.
Biochem. 167:507-512.
30. Purves, F. C., M. Katan, W. S. Stevely, and D. P. Leader. 1986.
Characteristicsof theinduction of a new protein kinase in cells infected with herpesviruses. J. Gen. Virol. 67:1049-1057. 31. Purves, F. C., R. M. Longnecker, D. P. Leader, and B. Roizman.
1987. Herpes simplex virus 1 protein kinase is encoded by open reading frame Us3 which is non essential for growth in cell culture. J. Virol. 61:2896-2901.
32. Purves, F. C., and B. Roizman. Unpublished results.
33. Rixon, F. J., and D. J. McGeoch. 1985. Detailed analysis of the mRNAs mapping in the short unique region of herpes simplex virus type 1. Nucleic Acids Res. 13:953-973.
34. Sefton, B. M. 1985. Oncogenes encoding protein kinases. Trends Genet. 1:306-308.
35. Seto, E., P. J. Mitchell, and T. S. Benedict Yen. 1990. Transac-tivation by the hepatitis B virus X protein depends on AP-2 and othertranscriptionfactors. Nature (London) 344:72-74. 36. Smith, R. F., and T. F. Smith. 1989. Identification of new
protein kinase-related genes in three herpesviruses, herpes simplex virus,varicella-zostervirus, and Epstein-Barr virus. J. Virol.63:451-455.
37. Southern, E. M. 1975. Detection of specific sequences among DNAfragmentsseparated by gelelectrophoresis. J.Mol. Biol. 98:503-517.
38. Wu, J. Y., Z.-Y. Zhou, A. Judd, C. A.Cartwright, and W. S.
Robinson. 1990. The hepatitis B virus-encoded transcriptional
trans-activatorhbx appears to be anovel protein serine/threo-nine kinase. Cell63:687-695.
39. Zhang, G., R. Stevens, and D. P. Leader. 1990. The protein kinase encoded in the short unique region of pseudorabies virus: description of the gene and identification of its product in virions and ininfected cells. J. Gen. Virol.71:1757-1765.