An RNA Element That Facilitates Programmed Ribosomal
Readthrough in Turnip Crinkle Virus Adopts Multiple Conformations
Micki M. Kuhlmann, Maitreyi Chattopadhyay, Vera A. Stupina, Feng Gao, Anne E. Simon
Department of Cell Biology and Molecular Genetics, University of Maryland—College Park, College Park, Maryland, USA
ABSTRACT
Ribosome recoding is used by RNA viruses for translational readthrough or frameshifting past termination codons for the syn-thesis of extension products. Recoding sites, along with downstream recoding stimulatory elements (RSEs), have long been stud-ied in reporter constructs, because these fragments alone mediate customary levels of recoding and are thus assumed to contain complete instructions for establishment of the proper ratio of termination to recoding. RSEs from theTombusviridaeand Luteo-viridaeare thought to be exceptions, since they contain a long-distance RNA-RNA connection with the 3=end. This interaction has been suggested to substitute for pseudoknots, thought to be missing in tombusvirid RSEs. We provide evidence that the phy-logenetically conserved RSE of the carmovirusTurnip crinkle virus(TCV) adopts an alternative, smaller structure that extends an upstream conserved hairpin and that this alternative structure is the predominant form of the RSE within nascent viral RNA in plant cells and when RNA is synthesizedin vitro. The TCV RSE also contains an internal pseudoknot along with the long-dis-tance interaction, and the pseudoknot is not compatible with the phylogenetically conserved structure. Conserved residues just past the recoding site are important for recoding, and these residues are also conserved in the RSEs of gammaretroviruses. Our data demonstrate the dynamic nature of the TCV RSE and suggest that studies using reporter constructs may not be effectively recapitulating RSE-mediated recoding within viral genomes.
IMPORTANCE
Ribosome recoding is used by RNA viruses to enable ribosomes to extend translation past termination codons for the synthesis of longer products. Recoding sites and a downstream recoding stimulatory element (RSE) mediate expected levels of recoding when excised and placed in reporter constructs and thus are assumed to contain complete instructions for the establishment of the proper ratio of termination to recoding. We provide evidence that most of the TCV RSE adopts an alternative structure that extends an upstream conserved hairpin and that this alternative structure, and not the phylogenetically conserved structure, is the predominant form of the RSE in RNA synthesizedin vitroand in plant cells. The TCV RSE also contains an internal pseudo-knot that is not compatible with the phylogenetically conserved structure and an RNA bridge to the 3=end. These data suggest that the TCV RSE is structurally dynamic and that multiple conformations are likely required to regulate ribosomal read-through.
P
ositive-sense RNA viruses employ a variety of gene expression strategies for the diversification of their proteomes (1,2). Two noncanonical mechanisms,⫺1 programmed ribosomal frame-shifting (⫺1PRF) and programmed ribosomal readthrough (PRT), circumvent stop codons for the expression of carboxy-terminal extension products, effectively economizing on the lim-ited genome size of most RNA viruses (3,4). In⫺1PRF, the elon-gating ribosome shifts back 1 residue at a 7-residue slippery sequence upstream of the stop codon and then continues translat-ing in the new readtranslat-ing frame, generattranslat-ing a fusion polypeptide. In PRT, a tRNA decodes the stop codon, and translation continues in the same reading frame. These processes are used by viruses mainly to express their RNA-dependent RNA polymerase (RdRp) or a coat protein (CP) extension product that is necessary for vector transmission and/or virion assembly (5). Ribosome recod-ing leads to the tightly controlled synthesis of 5% to 10% fusion products, and the maintenance of this level is critical for efficient virus amplification (6–10).Early studies used deletions to define minimal sequences capa-ble of stimulating⫺1PRF from recoding regions inserted into heterologous RNA. These studies identified a stem-loop structure
inRous sarcoma virus(11) and a pseudoknotted structure inAvian
infectious bronchitis virus(IBV) (12) that enhanced recoding
inde-pendently of additional viral sequences. Since then, nearly all re-coding studies have used similar approaches, which generally in-volve inserting the recoding site and downstream sequences between two luciferase reporter open reading frames (ORFs) and assaying for recoding-required luciferase activity either in cell ex-tracts (in vitro) or in cell culture (in vivo) (13–17). Using this approach, studies have confirmed that the majority of⫺1PRF and PRT events require the presence of a stable hairpin or pseudo-knottedrecodingstimulatoryelement (RSE) just downstream of the recoding site (18,19).
Despite 25 years of study, the specific mechanism(s) that me-diates the critical ratio of recoding to termination products
re-Received9 June 2016Accepted12 July 2016
Accepted manuscript posted online20 July 2016
CitationKuhlmann MM, Chattopadhyay M, Stupina VA, Gao F, Simon AE. 2016. An RNA element that facilitates programmed ribosomal readthrough in turnip crinkle virus adopts multiple conformations. J Virol 90:8575– 8591.
doi:10.1128/JVI.01129-16.
Editor:R. M. Sandri-Goldin, University of California, Irvine Address correspondence to Anne E. Simon, [email protected]. Copyright © 2016, American Society for Microbiology. All Rights Reserved.
on November 7, 2019 by guest
http://jvi.asm.org/
mains unresolved. For⫺1PRF, pseudoknotted structures are pro-posed to compromise the translocation process by forming a physical barrier that blocks the mRNA entrance channel, imped-ing the mechanical unwindimped-ing of the mRNA by the translatimped-ing ribosome (20). The stability of the stem of simple RSE hairpins has also been suggested to serve a similar function in promoting PRT and⫺1PRF (17,19,21). However, no consistent correlation has been found between frameshifting efficiency in reporter con-structs and the mechanical force required to unfold the RSE using optical tweezers (22,23). Instead, the suggestion has been made that frameshifting efficiency correlates with conformational plas-ticity and the tendency of the pseudoknot to form alternative local folds. The structural plasticity of the West Nile virus RSE was recently demonstrated, with both tandem stem-loop and pseudo-knotted structures fitting the structural data, suggesting that this RSE may comprise a molecular switch necessary for the control of levels of frameshifting (24). In addition, protonation of the H-type pseudoknot RSE ofMurine leukemia virus(MuLV) was sug-gested to be important for the formation of the readthrough-per-missive conformation (25); however, whether this represents a general mechanism remains an open question.
All of these studies make the assumption that fragments active for recodingin vitrocontain all instructions necessary for obtain-ing the proper ratio of termination to recodobtain-ing products durobtain-ing the translation of the virus. However, long-distance RNA-RNA interactions are required for efficient⫺1PRF inPea enation
mo-saic virusRNA 2 (PEMV2),Barley yellow dwarf virus(BYDV), and
Red clover necrotic mosaic virus(21,26–29) and for PRT in
Carna-tion Italian ringspot virusandTobacco necrosis virus D(30,31).
Since internal H-type pseudoknots have not been reported in RSEs from any of these plant viruses, it has been proposed that these long-distance RNA bridges form an atypical pseudoknot that functionally replaces the internal pseudoknot that is common in animal virus RSEs (2,28,32). In addition, hairpins or artificial duplexes just upstream of the recoding site repress frameshifting in coronaviruses and PEMV2 (21,33–35). Strikingly, in the ab-sence of the upstream hairpin, the PEMV2 RSE is no longer required to stimulate frameshifting from the slippery site, sug-gesting that upstream hairpins may play significant roles in suppressing⫺1PRF.
To circumvent methods that exclude proximal or distal viral sequences that might also be important for readthroughin vivo, we assessed PRT within full-length viral genomic RNA (gRNA) and choseTurnip crinkle virus(TCV) because of its small size, its well-characterized genome, and the simplicity of studying virus accumulation in single cells. TCV is a member of theCarmovirus
genus within theTombusviridaefamily and has a 4,053-nucleotide (nt) positive-sense genome containing five overlapping ORFs (Fig. 1A). The 5=-proximal ORF (p28, encoding a replicase-asso-ciated protein) and the p88 RdRp readthrough product are trans-lated from the gRNA, which lacks a 5=cap and 3=poly(A) tail (36, 37). The TCV RSE, which is located just downstream of the p28 amber termination codon, contains two asymmetric internal loops, one of which pairs with the apical loop of the terminal 3=-end Pr hairpin (30).
Here we report that in addition to the long-distance interac-tion between the RSE and 3=sequences, the TCV RSE contains an internal H-type pseudoknot, and both tertiary interactions are important for competent ribosome recoding. We also demon-strate that a short conserved sequence just downstream from the
recoding site is required for efficient PRT and that this sequence is similar to the sequence previously found at thegag-poljunction in gammaretroviruses (38,39). Most significantly, we provide struc-tural evidence that the lower stem of the TCV RSE adopts an alternative conformation that extends an upstream, phylogeneti-cally conserved hairpin and that this alternative structure is the predominant form of the RSEin vitroand in plant cells. These data reveal that at least three RSE conformations exist for TCV, show-casing the dynamic nature of RSEs and adding to the growing evidence that the current approaches— examining RSEs in isola-tion—may not effectively recapitulate the recoding process as it occurs within full-length viral gRNA.
MATERIALS AND METHODS
Generation of plasmid constructs.Mutant clones were generated using PCR-based site-directed mutagenesis from plasmid pTCV66, which con-tains the full-length TCV genome downstream of a T7 RNA polymerase promoter. Overlapping oligonucleotides (Integrated DNA Technologies) were used to introduce the desired mutations by use of Phusion high-fidelity polymerase (New England BioLabs) according to a one-step mu-tagenesis protocol (40). PCR products were subjected to DpnI digestion for 2 h at 37°C prior to transformation into DH5␣competent cells. Mu-tations were confirmed by sequencing in the region (Eurofins Genomics). In vitrotranslation.SmaI-digested TCV plasmids were subjected to
in vitrotranscription for 2 h at 37°C using T7 RNA polymerase. Uncapped RNA transcripts (1 pmol) were translated in 10l of wheat germ extract (WGE; Promega), supplemented with 100 mM potassium acetate and [35S]methionine, according to the manufacturer’s instructions. The
reac-tion mixture was incubated at 25°C for 2 h and was resolved on a 10% SDS-PAGE gel. The dried gel was exposed to a phosphorimager screen and was scanned by a FLA-5100 fluorescent image analyzer (Fujifilm). Band intensity was quantified using Quantity One software (Bio-Rad). Experiments were performed in triplicate using independently tran-scribed RNAs.
Protoplast transfection and RNA gel blots.Protoplasts generated from seed-derivedArabidopsis thalianacallus tissue (ecotype Col-0) were transfected within vitro-transcribed full-length TCV gRNA as described previously (41). Briefly, 20g of uncapped gRNA was transfected into 5⫻ 106cells and was incubated in the dark for 40 h. Total RNA was extracted,
subjected to electrophoresis, transferred to a nitrocellulose membrane, and probed using [␥-32P]ATP-labeled oligonucleotides complementary
to positions 3931 to 3953, 3869 to 3883, and 4035 to 4054 in the 3= un-translated region (3=UTR). The membrane was exposed to a phosphor-imager screen as described above. Experiments were performed in tripli-cate using independently transcribed RNAs.
SHAPE structure probing.Structure probing was performed using SHAPE (selective 2=-hydroxylacylation analyzed byprimerextension) as described previously (42). Briefly, forin vitroSHAPE, 6 pmol ofin vitro -transcribed, full-length TCV gRNA was denatured for 5 min at 65°C, snap-cooled on ice, and then incubated in folding buffer at 37° for 20 min. The folded RNA was treated either with N-methylisatoic anhydride (NMIA; final concentration, 15 mM) for base modification or with an equal volume of dimethyl sulfoxide (DMSO) as a negative control. The reaction mixtures were incubated at 37°C for 35 min, ethanol precipi-tated, and resuspended in 8l 0.5⫻Tris-EDTA (TE) buffer. Primer ex-tension was carried out using [␥-32P]ATP-labeled oligonucleotides and
SuperScript III reverse transcriptase (Invitrogen) as described previously (43). Primers complementary to TCV positions 940 to 964, 875 to 894, 839 to 862, and 995 to 1018 were used for probing of the RSE,stem-loop
A(SLA), and upstream and downstream region structures, respectively. Reaction products and dideoxy sequencing ladders (Roche) were resolved on an 8% denaturing polyacrylamide gel, exposed overnight to a phos-phorimager screen, and imaged using the FLA-5100 fluorescent analyzer (Fujifilm). Forin vivoSHAPE, protoplasts prepared fromArabidopsis
on November 7, 2019 by guest
http://jvi.asm.org/
thalianaseed callus cultures (5⫻106cells) were transfected with 20g of
uncapped full-length wild-type (WT) TCV gRNA or an equal amount of a nonreplicating (GDD¡GAA) polymerase active-site mutant. After 40 h of incubation at room temperature in the dark, either 1-methyl-7-nitroisatoic anhydride (1M7; final concentration, 5 mM) was added to the protoplasts for base modification or an equal volume of DMSO was added as a negative control, and cultures were gently shaken for 5 min at room temperature. IM7 was used in place of NMIA because NMIA has been reported to be inefficient at modifying RNA in a cellular environment (44, 45). Unmodified WT TCV was used to generate dideoxy sequencing lad-ders. Cells were collected by centrifugation; total RNA was extracted; and the RNA was resuspended in H2O. Primer extension, structure probing, resolution, and visualization were performed as described above. All SHAPE structure mapping was conducted in duplicate, independent ex-periments.
RESULTS
The TCV RSE contains an internal pseudoknot.Computational and phylogenetic approaches indicate that members of the
Tom-busviridaecontain two hairpins in the vicinity of the recoding site
irrespective of whether they use frameshifting or readthrough to synthesize their RdRp (Fig. 2). The 5=hairpin, known as stem-loop A (SLA), is within 10 nt of the amber termination codon. The 3=hairpin, previously identified as an RSE (30), is an unbranched structure with two large, asymmetric internal loops (L2 and L3),
of which L2 is engaged in the long-distance interaction with se-quences at the 3=end. The RSE is located just downstream of the UAG, with the guanylate nearly always participating in the termi-nal base pair at the base of the G·C-rich lower stem (S1).
Although RSE pseudoknots have not been reported in tombus-virids, visual inspection of TCV and other carmovirus RSEs re-vealed the potential for an internal pseudoknot between guany-late-rich residues in upper internal loop L3 and cytidylates that are also predicted to be base paired in the highly conserved S1 stem (Fig. 1Band2, blue). Internal loop mutation m1, which disrupts the putative pseudoknot, reduced the synthesis of p88 (and thus readthrough) by 97% in WGE, whereas stem S1 mutation m2, which would convert G·C to G:U pairs in S1 and the putative pseudoknot, did not affect readthrough (Fig. 1C). When the gRNA contained both m1 and m2, which should maintain the pseudoknot and S1, the level of p88 synthesis increased to 127% of the WT level. Although the m1 mutation was not a silent mutation (a proline was converted to a serine in the RdRp), mutant and WT gRNAs were evaluated for accumulation inArabidopsis thaliana
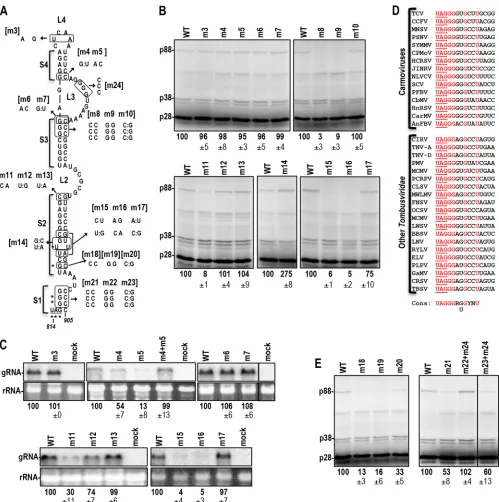
protoplasts (Fig. 1D). m1 reduced gRNA accumulation by 97%, and m2 reduced gRNA accumulation by 34%. The combination of the m1 and m2 mutations (m1⫹m2) was compensatory, with accumulation increasing (over m1 levels) to 71% of WT accumu-FIG 1TCV RSE contains an internal pseudoknot. (A) Genome organization of TCV. Genomic and subgenomic RNAs are shown. The long-distance RNA-RNA interaction connecting the RSE with the 3=-terminal Pr hairpin is indicated by a dashed red line. (B) Phylogenetically conserved structure of the TCV RSE and the upstream hairpin SLA. Designations of stems and loops in the RSE are shown. Hairpins in the location of the SLA relative to the RSE are conserved in the
Tombusviridae(seeFig. 2). Sequences in L2 and the Pr loop that engage in the long-distance interaction are shown in red. Putative pseudoknot residues are shown in blue. The locations of m1 and m2 mutations, generated to test for the pseudoknot, are shown. (C)In vitrotranslation of full-length wild-type (WT) and mutant gRNAs in WGE. The positions of p28 and the readthrough product, p88, are shown. p38 is the capsid protein and is translated by internal initiation. The product below p38 is an internal initiation product that is coterminal with the RdRp (data not shown). p38 was used as a loading control throughout this study. Average percentages relative to the WT value and standard deviations for three independent experiments are shown below panels throughout the figures. (D) RNA gel blot of WT and mutant gRNAs accumulating at 40 hpi inArabidopsisprotoplasts. The locations of the gRNA and the 26S rRNA are shown. Data are from three independent experiments.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.76.512.65.342.2]FIG 2Phylogenetic conservation of RSE and SLA structures. (A) Carmoviruses:Cardamine chlorotic fleck virus(CCFV),Saguaro cactus virus(SCV),Melon necrotic spot virus(MNSV),Pelargonium flower break virus(PFBV),Calibrachoa mottle virus(CbMV). Sequences in blue are predicted to form an internal pseudoknot in an alternative structure. Sequences in red are predicted to pair with complementary sequences at the 3=end of the genome. (B)Carnation Italian ringspot virus(CIRV) is a member of the genusTombusvirus.Panicum mosaic virus(PMV) is a member of the genusPanicovirus. Umbravirus PEMV2 is unclassified.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.64.519.57.686.2]lation. These results strongly suggest that in addition to the phy-logenetically conserved RSE structure that contains stem S1, the RSE must form an alternative structure without S1 and with an internal pseudoknot for efficient readthroughin vitro.
The structure of the TCV RSE is important for function.To further analyze the importance of RSE sequence and structure,
mutations in loops and mutations designed to be compensatory in stems were generated throughout the hairpin (Fig. 3A). If stem alterations could be designed that were both compensatory in the RSE and silent in the RdRp, levels of mutant gRNAs were also assessed in protoplasts. Mutating the terminal loop (m3) or the base pair just below internal loop L3 (m6, m7) did not affect read-FIG 3Analysis of the TCV RSE. (A) Mutations generated in the RSE. Asterisks indicate residues conserved at the readthrough site throughout theTombusviridae
(see panel D). When possible, base alterations were chosen for being silent in the encoded RdRp and for maintaining codon usage similar to that of the WT residue, which allowed for the gRNA to be assayedin vivo(see panel C). The designations of the mutations are given in brackets. (B)In vitrotranslation of full-length WT and mutant gRNAs in WGE. (C) Accumulation of selected gRNA mutants in protoplasts, assayed using RNA gel blots. The line within the upper right blot indicates that a lane was discarded. (D) Sequence conservation adjacent to the amber codon in carmoviruses and all other members of the Tombus-viridaethat were examined. (E)In vitrotranslation of full-length WT and mutant gRNAs with alterations in the conserved residues. The line indicates that a lane from the blot was discarded.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.44.543.63.565.2]through or gRNA accumulationin vivo(Fig. 3BandC). Mutating the three base pairs just below L3 (m8, m9) reduced PRT to back-ground levels, whereas the compensatory mutation (m10) re-stored p88 synthesis to WT levels. Altering the base pair in S4 that is adjacent to L3 (m4, m5) decreased gRNA accumulationin vivo
but did not affect readthroughin vitro. The combination of the two alterations was compensatory for gRNA accumulationin vivo, indicating that S4 disruption might have consequences that were not discernible in the WGE assay. Altering the guanylate just be-low L2 to an adenylate (m11) disrupted pairing at this location and reduced PRT to 8% of the WT level and gRNA accumulation to 30% of WT accumulation. Maintaining pairing in this location (m12) had no effect on readthrough and reduced gRNA accumu-lation by only 26%. gRNA containing both mutations (m13) gave WT levels of readthrough and accumulationin vivo, suggesting that a base pair at this position is important. Altering two base pairs simultaneously in stem S2 (m15, m16) reduced readthrough and gRNA accumulation by 20-fold, whereas the compensatory mutation (m17) restored readthrough to 75% of WT levels and gRNA accumulation to WT levels. Stabilizing the S2 stem by con-verting the adjacent G:U pair and U-U mismatch to G·C and A·U (m14) enhanced readthrough to 275% of the WT level. The latter result correlates with published reports indicating that strength-ening the stability of RSE lower stems positively impacts PRT and⫺1PRF (17, 21). These results establish the importance of RSE stems S2 and S3 for stimulating PRT.
Sequence adjacent to the readthrough site is conserved in tombusvirids and is important for readthrough. The read-through sites in all 15 carmoviruses and in all other tombusvirids examined (Fig. 3D) contain a conserved sequence [UAG GGR (G/U)G YNU (the termination codon is underlined)]. Conserva-tion of the GGR, but not the (G/U)G, 3=of the UAG has been reported previously for viruses in theTombusviridaeundergoing readthrough (19), and a similar 5-base purine-rich sequence fol-lows the stop codon between thegagandpol genes inMurine
leukemia virusand is important for readthrough (38,39). Either
the G821C (m18) or the C986G (m19) alteration reduced read-through to 13% or 16% of WT readread-through, respectively (Fig. 3A andE). The two alterations together (m20), which should re-form the S2 stem, improved readthrough to only 33% of the WT level. This is in contrast with the two nearby base pair alterations (m15 and m16) that decreased readthrough to 5% of the WT level, with the compensatory change (m17) improving readthrough to 75% of the WT level. These results suggest that both base pairing and the identity of G821 and/or C986 are important for readthrough. To determine the importance of the guanylates adjacent to the termination codon, the three guanylates in positions⫹1,⫹2, and
⫹3 were converted to cytidylates (m21), and the opposing cytidy-lates were converted to guanycytidy-lates (m22). Because the m22 muta-tions would disrupt the required pseudoknot, they were com-bined with corresponding changes in internal loop L3 (m24) to maintain the tertiary interaction. m21 reduced readthrough by nearly 50%, whereas m22 (⫹m24) had no detrimental effect (Fig. 3E). These results indicate that the S1 stem can be disrupted with-out negatively impacting readthrough as long as mutations are on the 3=side and the required pseudoknot is not disrupted. The combination of m21 with m22 (compensatory mutation m23), which, along with m24, should restore the S1 stem and maintain the pseudoknot, reduced readthrough from that with m22⫹m24. The noncompensatory nature of the residues in stem S1 suggests
that one or more of the conserved guanylates just downstream of the amber codon are required for efficient readthrough.
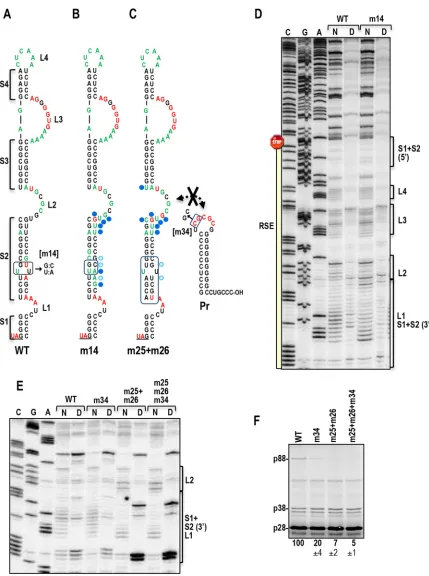
SHAPE structure probing of the RSE regionin vitroandin vivo.SHAPE (selective 2=-hydroxyl acylation analyzed by primer extension) was used to structurally map the SLA/RSE region in full-length TCV gRNA preparedin vitro(Fig. 4AtoC). SHAPE detects flexible residues whose 2=OH can form covalent bonds with NMIA, which is an impediment for reverse transcriptase-mediated primer extension. Residues with moderately high to high reactivity with NMIA are shown in red, and residues with low to moderate reactivity are shown in green, inFig. 4A. SHAPE data supported the predicted structure of the SLA but was consistent only with the upper portion of the RSE (Fig. 4AtoC). In the lower portion of the RSE, three residues on the 3=side of stem S2 that were predicted to be paired were strongly reactive with NMIA, whereas their partner resides were not. This suggests that despite the importance of the S2 stem for PRTin vitro, it was not present at discernible levels within the gRNA population that was used for both SHAPE and WGE. The strong reactivity of L3 pseudoknot guanylates, but not their paired cytidylates, also suggests that the PRT-required pseudoknot was not present in thein vitro -synthe-sized gRNA. Internal loop L2, which engages in the long-distance interaction with the Pr loop, contained a mixture of flexible and nonflexible residues. These residues were similarly flexible when gRNA that contained mutations in the Pr loop designed to disrupt the long-distance interaction was used (seeFig. 7E). Thus, this necessary tertiary interaction also was not detected in gRNA pre-paredin vitro.
Since RNA folds cotranscriptionally, the absence of both the phylogenetically conserved stem S2 and the two required tertiary interactions could be due to misfolding of thein vitro-synthesized, denatured, and then renatured gRNA. Therefore, SHAPE was re-peated using viral gRNA accumulating within infected protoplasts
(in vivo SHAPE). gRNA that was capable (WT) or incapable
(GDD) of amplification (due to mutations in the RdRp active site) was subjected to SHAPE at 40 h postinoculation (40 hpi), since plant viruses are restricted to initially inoculated protoplasts, and little virus accumulates after this point. Cells were treated with 1M7 (a membrane-permeant electrophile that generated an in
vitro SHAPE profile identical to that with NMIA [data not
shown]), followed by cell lysis, total-RNA extraction, and primer extension. As shown inFig. 4EandF, and in the secondary struc-ture inFig. 4D, thein vivoSHAPE profile for the RSE region at this time point was very similar to thein vitroSHAPE profile (exclud-ing consideration of the guanylates in L3 that could not be as-sessed), suggesting that the structure of the RSE region in thein
vitro-synthesized gRNA does not represent a kinetically trapped
artificial structure. Thein vivoSHAPE profile was not due to re-sidual input RNA, since no SHAPE data were obtained using the GDD mutant (with the exception of the strong signal in all lanes at the position of the pseudoknot guanylates, which was likely asso-ciated with the radiolabeled oligonucleotide). The one major dif-ference between thein vivoandin vitroSHAPE profiles was addi-tional reactive residues on the 3= side of stem S2, strongly suggesting that at least some portion of the phylogenetically con-served S2 stem is not present in the majority of gRNAs sampledin
vivoandin vitro.
To investigate whether the S1 stem is present inin vitro -syn-thesized gRNA, all four S1 3=-side cytidylates were changed to guanylates (m32), and the mutant gRNA was subjected toin vitro
on November 7, 2019 by guest
http://jvi.asm.org/
on November 7, 2019 by guest
http://jvi.asm.org/
SHAPE (Fig. 5). The flexibilities of the altered residues on the 3= side of S1 (and adjacent sequences) were enhanced; however, no change in the flexibility of the 5=-side guanylates was detected (Fig. 5B). In addition, the flexibility of L3 internal-loop guanylates that form a pseudoknot with these cytidylates was also unchanged. When the L3 guanylates were also converted to cytidylates (m32⫹m33) with the hope of driving the formation of the pseu-doknot, the mutated L3 residues were no longer flexible. How-ever, the S1 3=guanylates in the m32⫹m33 mutant showed little change in flexibility from that with m32 alone, suggesting that the pseudoknot still was not forming in the majority of gRNAs in the
population and implying that the new cytidylate residues in L3 were likely pairing elsewhere. m32 reduced readthrough by 20-fold in WGE, and m32⫹m33 restored WT readthrough levels, indicating that the pseudoknot is still required and still capable of formation during the translation of the gRNA (Fig. 5C). These results strongly suggest that the S1 stem and RSE pseudoknot are not present in the majority of the gRNAs synthesizedin vitro.
Sequences upstream of SLA pair with RSE 5=-side S1 and S2 residues.To determine if the middle portion of the S2 stem exists in gRNA synthesizedin vitro, two additional mutant gRNAs were generated (m25, m26), each with seven altered residues in S2 that,
FIG 4SHAPE structure probing of the readthrough region in full-length gRNAin vitroandin vivo. (A) Locations of NMIA-reactive residues in the phyloge-netically conserved structure of the RSE, obtained using SHAPE profile data generated fromin vitro-synthesized gRNA (see panels B and C). Residues with moderately high to high reactivity are shown in red, and residues with moderately low to moderate reactivity are shown in green. (B and C) SHAPE mapping of the RSE and SLA regions inin vitro-synthesized gRNA. The gel in panel C is a longer run of the samples shown in panel B. The first three lanes are nucleotide ladder lanes. N, NMIA treated; D, DMSO treated. The locations of different regions in the RSE and SLA are shown to the right of the panels; 5=and 3=denote the side of hairpin. The location of the amber stop codon is shown. (D) Locations of IM7-reactive residues in the phylogenetically conserved structure of the RSE, obtained using SHAPE profile data generated from gRNA accumulating in protoplasts (see panels E and F). Since IM7 is membrane permeant, it was used in place of NMIA. (E and F) SHAPE mapping of the RSE and SLA regions in gRNA accumulatingin vivoat 40 hpi. The gel in panel F is a longer run of the samples shown in panel E. The first three lanes are nucleotide ladder lanes. I, IM7 treated; D, DMSO treated.
FIG 5Compensatory mutations in the RSE pseudoknot do not result in detection of the pseudoknot inin vitro-synthesized gRNA. (A) Locations of mutations. (B) The first three lanes are nucleotide ladder lanes. N, NMIA treated; D, DMSO treated. Specific locations in the RSE are marked by brackets. Note that the m33 alteration reduced the flexibility of local (mutated) residues but did not substantially reduce the flexibility of altered S1 partner residues. This indicates that the pseudoknot is not present in this folded RNA. In addition, the flexibility of the 5=S1 residue was unaltered, confirming that the S1 stem also is not present. (C) Effects of mutations on readthrough in WT and mutant gRNAs preparedin vitro.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.81.501.67.418.2]together, are complementary (Fig. 6A).In vitrotranslation of m25, m26, and m25⫹m26 gRNAs generated only low (6 to 8% of WT) levels of p88 (Fig. 6B). Probing of the structure of m25 gRNA, which contained mutations on the 5=side of S2, indicated that the mutated resides, as well as a few adjacent residues, became flexible (a total of 12 nt). On the 3=side of the RSE, there were only minor changes in the S2 SHAPE profile, while a guanylate (G783) in loop L2 became strongly reactive (Fig. 6C). m26 gRNA, with mutations on the 3=side of S2, contained residues in the altered region that both gained and lost flexibility, with flexibility changes extending into the L2 internal loop. No flexibility changes were observed on the opposing 5=side of S2. In m25⫹m26 gRNA, the mutated res-idues on the 3=side were no longer flexible, suggesting that the majority of the S2 stem was now forming. In addition, the upper portion of S2 and the lower portion of L2 contained large numbers of residues with enhanced flexibility, including G783. Taken to-gether, these results strongly suggest that the entire S2 stem, in addition to the S1 stem, does not form in the gRNA synthesizedin
vitro.
In m25⫹m26 gRNA, the structure of a portion of L2, which engages in the long-distance interaction with 3=sequences, was significantly altered when either the S2 stem formed or the alter-native pairing of S2 residues was disrupted. The SHAPE profile of m14 gRNA, which contains a strengthened S2 stem (Fig. 3), dis-played structural changes in this region similar to those of m25⫹m26 gRNA (Fig. 7BandD). Since readthrough was en-hanced in m14 gRNA (Fig. 3) and depressed in m25⫹m26 gRNA (Fig. 6B), the altered L2 structure was not by itself responsible for weak readthrough in m25⫹m26 gRNA. To determine if the S2/L2 structural changes affected the long-distance interaction between L2 and the 3=end, SHAPE was conducted on m25⫹m26 gRNA that also contained a 2-base alteration in the 3=-end interacting sequence (mutation m34). No difference in L2 flexibility from that in WT gRNA was found in m34 or m25⫹m26⫹m34 gRNA, suggesting that the flexibility changes in L2 did not lead to detec-tion of the long-distance interacdetec-tion (Fig. 7E). In addidetec-tion, m34 reduced WT gRNA readthrough by 80% but had no comparable effect on m25⫹m26 gRNA (Fig. 7F). These results support our conclusion that the S2 stem forms in m25⫹m26 gRNA but not in WT gRNA synthesizedin vitroand that either the presence of the S2 stem or the disruption of alternative pairing of S2 residues affects residue flexibility in and near the L2 internal loop.
A region upstream of SLA is implicated in alternative pairing with RSE S2 residues. In the SHAPE profiles for m25 and m25⫹m26 gRNAs, a small region located just upstream of SLA (residues 720 to 735) showed enhanced flexibility (Fig. 6C, blue bracket). No discernible differences were detected either upstream or downstream of this region, and no flexibility changes were dis-cernible in this region in m26 gRNA. When residues 720 to 735 were altered (m27) and the SHAPE profile of m27 gRNA was compared with that of WT gRNA, the mutated region in m27 gRNA became more flexible (Fig. 8). In addition, 13 of 26 residues in S2 showed altered flexibility, as did residues in the lower por-tion of L2. Since most S2 residues had increased flexibility, muta-tion of residues 720 to 735 did not by itself result in the formamuta-tion of the phylogenetically conserved S2 stem. The presence of very similar flexibility changes in upper 3=S2 and lower L2 residues suggests that this flexibility signature (also found for m14 and m25⫹m26 gRNAs [Fig. 7DandE]) is not directly associated with
the formation of the S2 stem but rather reflects a loss of pairing between S2 resides and residues in positions 720 to 735.
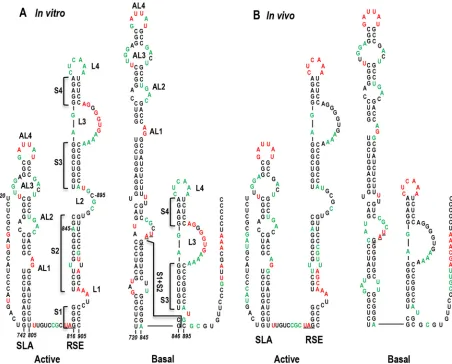
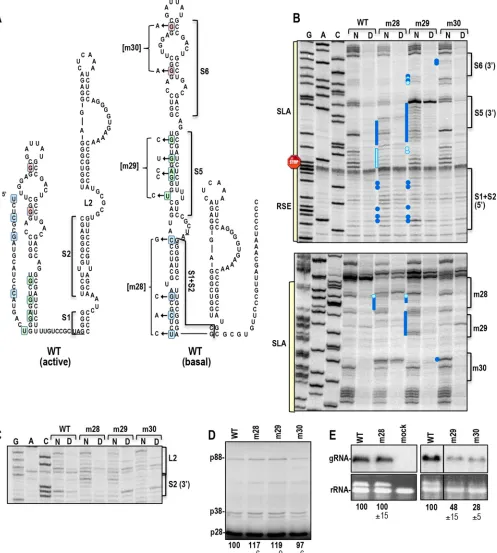
Alternative structure of the TCV RSE.When the sequence between residues 720 and 735 was examined, clear base-pairing possibilities with residues on the 5=side of S1 and S2 were discern-ible, which extended the length of SLA (Fig. 9). This alternative conformation, labeled the “basal” structure, agrees well with the SHAPE structure profile determined bothin vitro(Fig. 9A) andin vivo(Fig. 9B). To determine the importance of SLA and the basal structure, three sets of silent mutations were generated on the 5= side of the presumptive elongated SLA: mutations of the lower stems S1 and S2 (S1⫹S2) (m28), the middle stem S5 (m29), and the upper stem S6 (m30) (Fig. 10A). m28 gRNA caused several residues to gain flexibility on the opposing (3=) side of the stem (Fig. 10B). These changes were accompanied by the signature structural changes in the upper S2 (3=) and lower L2 regions of the RSE (Fig. 10C), as was found when RSE stem S2 was strengthened (m14, m25⫹m26 [Fig. 7DandE]) and/or when alternative pair-ing for S2 residues was disrupted (m27 [Fig. 8]). In addition, m28 mutations increased the flexibility of residues in the S5 stem of the SLA, suggesting the presence of higher-order structure within the extended hairpin (Fig. 10B). SHAPE structure probing of gRNA with m29 mutations in SLA S5 showed significant disruptions on both sides of the S5 stem (Fig. 10B). In addition, structural changes very similar to those found for m28 were evident in the S1⫹S2 region of the extended SLA, accompanied by signature changes in the RSE (Fig. 10BandC). These results support a high-er-order structure encompassing residues in the lower half of the basal SLA structure and the L2/upper S2 3=region of the RSE. In contrast, m30 mutations in the upper stem S6 of SLA enhanced the flexibility of several S6 stem residues without discernibly af-fecting the flexibility of the remainder of the SLA.
m28 and m29 caused minor (⬍20%) increases in p88 transla-tion in WGE, whereas m30 did not affect readthrough (Fig. 10D). This suggests that weakening the alternative pairing of RSE S1⫹S2 may only slightly promote the formation of the active form of the RSE. m29 and m30 reduced gRNA levels in protoplasts to 48% and 28% of WT levels, respectively (Fig. 10E). In contrast, m28 mutations did not affect gRNA levels. Although these results sup-port the imsup-portance of SLA for gRNA accumulationin vivo, free-ing the 5=side of the RSE lower stem is not sufficient to cause a discernible effect on virus accumulation.
DISCUSSION
Conserved features of tombusvirid RSEs.We initiated this study with the hypothesis that few overarching mechanisms exist for ribosome recoding because most currently used approaches might be excluding sequences necessary for proper regulation of the event. To provide evidence for or against this hypothesis, we ex-amined RSE regions in tombusvirids involved in either⫺1PRF or PRT. With few exceptions, these RSEs are ca. 90 nt long, with a G·C-rich lower stem that incorporates the guanylate of the amber stop codon at the base. Tombusvirid RSEs contain at least two large, asymmetric internal loops, with the lower loop engaged in an important long-distance interaction with the 3=end (28,30, 31). This RNA bridge had been proposed to form an atypical pseu-doknot in the RSE that functionally replaces the pseupseu-doknot com-monly found in animal virus RSEs (30). However, our compen-satory mutagenesis revealed that a phylogenetically conserved pseudoknot is also a critical feature of the TCV RSE (Fig. 1),
on November 7, 2019 by guest
http://jvi.asm.org/
FIG 6Alternative pairing partners for RSE 5=-side stem S2 residues. (A) (Upper left) WT SHAPE profile. The locations of mutations (m25 and m26) generated in full-length TCV gRNA are shown. Residues are color coded as described in the legend toFig. 4A. (Top right) Effects of m25 mutations on NMIA-reactive residues in the RSE region. Dark blue and light blue circles designate residues that were more or less reactive, respectively, than WT residues. (Bottom left) Effects of m26 mutations on reactive residues in the RSE region. (Bottom right) Effects of a combination of m25 and m26 mutations that should be compensatory. (B)
In vitrotranslation of full-length WT and mutant gRNAs in WGE. (C) SHAPE mapping of the RSE and SLA regions in WT and mutant gRNAs preparedin vitro. The first three lanes are nucleotide ladder lanes. N, NMIA treated; D, DMSO treated. The blue bracket delineates a region upstream of SLA that becomes flexible when 5=S2 residues are altered (m25). Note that the mutated residues on the 5=side of S2 are flexible in m25, whereas the partner residues on the 3=side of S2 retain WT-like flexibility.
on November 7, 2019 by guest
[image:10.585.65.519.36.624.2]FIG 7m14 and m25⫹m26 share similar structural alterations in the upper 3=S2 and lower L2 regions. (A) Locations of mutations in the RSE that stabilize the stem of S2. (B) Effects of m14 mutations on the SHAPE profile of the RSE. Dark blue and light blue circles mark residues that were more or less reactive, respectively, than WT residues. (C) SHAPE profile of m25⫹m26 (compensatory mutations in the lower S2 stem) gRNA fromFig. 6A, provided here for purposes of comparison. The hairpin at the right is the 3=-terminal Pr hairpin, whose apical loop is the pairing partner of L2. Residues complementary with the L2 sequence are shown in red. The m34 mutations that disrupt the long-distance interaction are shown. (D) SHAPE mapping of the RSE region in WT and m14 gRNAs preparedin vitro. The first three lanes are nucleotide ladder lanes. N, NMIA treated; D, DMSO treated. Note that the structural changes in upper 3=S2 and lower L2 are very similar to those found in m25⫹m26 gRNA (see panel E). (E) SHAPE mapping of the L2 region when the RSE contains m25⫹m26, and the long-distance interaction is either permitted (m25⫹m26) or not permitted (m25⫹m26⫹m34). This gel also shows that there is no difference in the WT L2 sequence when the long-distance RNA-RNA interaction is disrupted. (F)In vitrotranslation of WT and m25⫹m26 gRNAs that can or cannot (⫹m34) engage in the long-distance RNA-RNA interaction.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:11.585.76.505.60.636.2]gesting that the function of the long-distance interaction is not a simple substitution for a local H-type pseudoknot.
Conserved residues [UAG GGR (G/U)G YNU (the termina-tion codon is underlined)] were also identified just downstream of the amber termination codon in tombusvirids that use PRT to synthesize their RdRp. The conserved guanylates adjacent to the UAG contributed to PRT efficiency, but not as simple participants in the S1 stem. In addition, the G at position⫹5 (⫹5G) was also important for readthrough (Fig. 3). Gammaretroviruses share a similar conserved sequence downstream of their amber termina-tion codon at thegag-poljunction (UAG GGN NGN) (Fig. 11A). Alterations of⫹1G,⫹2G, and⫹5G (but not the intervening⫹3A and⫹4G) have been reported to be highly detrimental for read-through inMurine leukemia virus(MuLV) by use of
dual-lucifer-ase reporter constructs (8,39).⫹5G is located within the ribo-some when the amber codon is in the A site, suggesting that specific contacts may exist between this guanylate and ribosome components that support readthrough in these viruses.
The TCV RSE adopts several alternative conformations. Tombusvirid RSEs are structurally conserved, particularly in the lower stem regions (S1 and S2) (Fig. 2). However, SHAPE con-ducted bothin vivoandin vitrodid not support the presence of these stems in the TCV RSE. Rather, SHAPE profiles supported the predominance of an alternative structure, where residues on the 5=side of RSE S1 and S2 pair with sequences located 100 nt upstream, extending the base of the upstream SLA hairpin. Hair-pins in the same location as SLA are predicted to form upstream of RSEs in all tombusvirids examined (Fig. 2). A hairpin in this loca-FIG 8Altering the proposed pairing partners for the 5=-side S1/S2 residues changes the flexibility of S1/S2. (A) (Left) WT SHAPE profile. The location of m27 mutations generated in full-length TCV gRNA is shown. (Right) Effects of m27 mutations on the SHAPE profile. Residues are color coded as described in the legend toFig. 4A. Dark blue and light blue circles are explained in the legend toFig. 6A. (B) SHAPE mapping of the RSE and SLA regions in WT and mutant gRNAs preparedin vitro. The first three lanes are nucleotide ladder lanes. N, NMIA treated; D, DMSO treated. The blue bracket delineates the region upstream of SLA containing the m27 mutations. Note that the m27 mutations resulted in enhanced flexibility on both sides of the active RSE stem extending into the L2 region.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:12.585.76.511.66.480.2]tion was also previously reported in BYDV (26), as well as in um-braviruses (21) and coronaviruses (33), where they serve as re-pressors of recoding. The PEMV2 SLA strongly repressed frameshifting from the slippery sequence when the RSE structure was absent, possibly by blocking the backward movement of the ribosome (21). In TCV, mutations disrupting the middle and up-per portions of the extended TCV SLA reduced gRNA
accumula-tionin vivo(Fig. 10E). Although SLA in tombusvirids differ in
sequence and structure, strong conservation of a hairpin just up-stream of the recoding site suggests a role for this hairpin in both readthrough and frameshifting.
While SHAPE only provides a structural snapshot of the ma-jority of RNAs sampled, our results demonstrate that the structure with expanded SLA and truncated RSE (designated the basal structure) is the predominant formin vitroand at 40 hpiin vivo, when little if any translation would be occurring (plant viruses are restricted to initially infected cells in culture). We propose that the basal structure is the readthrough-inactive form of the RSE, which, when present, results in the ribosome terminating transla-tion at the p28 amber codon. A major issue that remains
unre-solved is the pairing location(s) for residues on the 3=side of the RSE in the basal structure. Although a number of possible pairing sites both upstream and downstream of the RSE were examined, none of these locations were structurally altered by mutations in the 3=side of the RSE (and elsewhere) that would have been ruptive (data not shown). m27 mutations, which extensively dis-rupt the pairing partner for the 5=side of RSE S1⫹S2, did not lead to widespread formation of the phylogenetically conserved RSE S1⫹S2 stem (Fig. 8B). This suggests that it may also be necessary to free the 3=side of the RSE S1⫹S2 stem from its paired location before the S1⫹S2 stem can form. Silent m28 mutations that dis-rupted four of the pairing partners for the 5=side of RSE S1⫹S2 were not detrimentalin vivo(Fig. 10E), suggesting that the basal structure locations for both sides of the RSE may need to be mod-ified before an effect on virus accumulation is discernible.
Although no structural evidence for the phylogenetically con-served active form of the TCV RSE currently exists, genetic evi-dence clearly demonstrated that an RSE containing the S2 stem is critical for recodingin vitro(Fig. 3). The phylogenetically con-served structure that includes stem S1 is also not compatible with FIG 9Proposed structure of the alternative, prominent RSE conformation. (A) WT SHAPE profile for gRNA synthesizedin vitro. (Left) The phylogenetically conserved conformation is suggested to be the “active” form of the RSE, which also requires engagement in the long-distance interaction with the 3=end and the formation of the pseudoknot for activity. (Right) The most prominent conformation of the RSE is proposed to be the “basal” form, which must convert to the active form for readthrough. (B) (Left) Phylogenetically conserved conformation with thein vivogRNA SHAPE profile. (Right) Proposed basal structure with thein vivoSHAPE profile.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:13.585.67.519.66.430.2]FIG 10Extended SLA contains higher-order structure and is important for gRNA accumulationin vivo. (A) Locations of the m28, m29, and m30 mutations in the basal and active structures of the RSE. Each set of mutations has a different background color for its residues: blue for m28, green for m29, and pink for m30. (B) Effects of SLA mutations on the SHAPE profiles of SLA and the RSE. Dark blue and light blue circles designate residues that were more or less reactive, respectively, than WT residues. Note that m28 and m29 cause nearly reciprocal structural changes in the adjacent stems. (C) Effects of SLA mutations on the structure of the RSE. Note that m28 and m29 cause alterations in residue flexibility in lower L2 and adjacent upper S2 (3=) similar to those caused by m14, m25⫹m26, and m27 (Fig. 6and7). (D)In vitrotranslation of full-length WT and mutant gRNAs in WGE. (E) RNA gel blot of WT and mutant gRNAs accumulating at 40 hpi. The line indicates that a lane was removed from the blot.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:14.585.47.544.69.622.2]the required pseudoknot between S1 and L3 residues. This suggests that a series of conformational changes are likely needed for the re-coding event. One possible scenario would have ribosomes unwind-ing both sides of the lower stem of the extended SLA basal structure during translation, which causes the signature structural changes in RSE L2 that might lead to the establishment of the long-distance interaction with the 3=end, possibly helping to stabilizing the RSE active structure, as was found for PEMV2 (21). When the RSE 3=-side S1 cytidylates form the pseudoknot, this might further stabilize the
active structure, leading to ribosome pausing and insertion of a tRNA that decodes the amber stop codon.
Evidence for additional RSE conformations in unrelated vi-ruses.Since viral RNAs are known to adopt alternative conforma-tions of overlapping sequences to facilitate transiconforma-tions between reversible processes (46), conformational switches modulating ri-bosome recoding might be widespread. In addition to West Nile virus (24), the presence of alternative structures has been sug-gested by SHAPE structure modeling of the RSE region in full-FIG 11Sequence conservation and conformational similarities between the RSEs of TCV and unrelated viruses. (A) The conserved sequence (cons) at the PRT site in tombusvirids is conserved in gammaretroviruses engaged in PRT of amber codons. The conserved sequence is highlighted. MuLV,Murine leukemia virus; RSV,Rous sarcoma virus; PERV,porcine endogenous retrovirus; OHEV,Odocoileus hemionus endogenous virus; FLV,Feline leukemia virus; BEV,Baboon endoge-nous retrovirus; MDEV,Mus dunni endogenous virus; DIAV,Duck infectious anemia virus; RfRV,Rhinolophus ferrumequinum retrovirus. (B) Proposed basal structure for the MuLV RSE, obtained using sequences upstream of the RSE. (Left) Active structure. (Right) A possible basal structure proposed previously (38, 39). (Center) An alternative possibility for the basal structure involving additional 5=sequences not considered in the earlier study. Residues are shown in blue to assist with orientation. All substitutions in the 24 viruses examined (shown in red) maintain both active and proposed basal structures. The stop codon is boxed, with a yellow background. (C) Proposed basal SLA structure for SARS-CoV. The upstream SLA hairpin serves as a repressor of PRF (33–35). Sequences shown in blue are capable of pairing. One hundred SARS-CoV isolates were examined for sequence differences, and all unique variants are shown (with GenBank accession numbers in parentheses). All sequence variations maintain the SLA and the proposed basal structure pairing.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:15.585.44.543.59.509.2]length HIVin vitro, which indicated that the previously proposed two-helix model (9, 47, 48) was missing upstream and down-stream sequences that together generate a more complex structure (49). Ribosomes translating through this larger frameshifting do-main were proposed to remodel portions of the extended struc-ture into the conventionally accepted two-helix active strucstruc-ture. The RSE of MuLV, in addition to having the same conserved gua-nylates proximal to the termination codon as the TCV RSE (38, 39) (Fig. 11B), can also adopt an alternative hairpin conformation (50,51). Mutations specifically affecting this alternative hairpin structure (but not the pseudoknotted structure) were highly det-rimental to virus viability (51). These data are consistent with MuLV also having basal and active conformations of its RSE.
Since the severe acute respiratory syndrome coronavirus (SARS-CoV) SLA hairpin was also repressive for recoding (33), we examined the sequence of the SARS-CoV SLA/RSE region to de-termine whether an alternative conformation that extends the length of the SLA, as in TCV, might be possible. As shown inFig. 11C, an alternative structure that extends the length of SLA and contains a 10-bp stem is predicted. This SLA extension is formed by pairing sequence upstream of the SLA with sequence from the 5=side of the lower RSE stem, similarly to TCV. Variants of the sequence found in a minority of SARS-CoV isolates maintain the structure of both SLA and the extended stem. Thus, for TCV and possibly SARS-CoV, SLA may serve as a scaffold for extension of its lower stem by base-pairing residues upstream of SLA with RSE residues, thereby disrupting the structure of the active RSE.
In conclusion, current models proposed to explain how RSEs mediate either⫺1PFR or PFT at the precise ratios found in in-fected cells require the assumption that all sequences necessary for regulating recoding are encompassed within the recoding site and downstream RSE (13, 20, 25). However, the original strategies used to identify active RSEs would have eliminated repressive/regu-latory upstream hairpins that might be key components in the for-mation of alternative basal forms of RSEs. Our results support the necessity of examining recoding within full-length viral gRNA in or-der to account for the full spectrum of RNA elements necessary to precisely regulate this critical event in the life cycle of RNA viruses.
ACKNOWLEDGMENTS
We thank all members of the Simon lab for helpful discussions and Pam-ela Katali for technical assistance. We also thank J. Dinman for providing the 1M7.
This work was supported by the National Science Foundation (MCB-1411836) and National Institutes of Health (R21AI117882-01) (to A.E.S.). M.M.K. and M.C. were partly supported by NIH Institutional Training Grant 2T32AI051967-06A1.
FUNDING INFORMATION
This work, including the efforts of Anne E. Simon, was funded by HHS | National Institutes of Health (NIH) (R21AI117882-01). This work, in-cluding the efforts of Micki M. Kuhlmann and Maitreyi Chattopadhyay, was funded by HHS | National Institutes of Health (NIH) (2T32AI051967-06A1). This work, including the efforts of Anne E. Simon, was funded by National Science Foundation (NSF) (MCB-1411836).
REFERENCES
1.Dreher TW, Miller WA.2006. Translational control in positive strand RNA plant viruses. Virology 344:185–197. http://dx.doi.org/10.1016/j .virol.2005.09.031.
2.Firth AE, Brierley I.2012. Non-canonical translation in RNA viruses. J Gen Virol93:1385–1409.http://dx.doi.org/10.1099/vir.0.042499-0. 3.Bidou L, Rousset J-P, Namy O.2010. Translational errors: from yeast to
new therapeutic targets. FEMS Yeast Res10:1070 –1082.http://dx.doi.org /10.1111/j.1567-1364.2010.00684.x.
4.Caliskan N, Peske F, Rodnina MV.2015. Changed in translation: mRNA recoding by⫺1 programmed ribosomal frameshifting. Trends Biochem Sci40:265–274.http://dx.doi.org/10.1016/j.tibs.2015.03.006.
5.Beier H, Grimm M. 2001. Misreading of termination codons in eu-karyotes by natural nonsense suppressor tRNAs. Nucleic Acids Res29: 4767– 4782.http://dx.doi.org/10.1093/nar/29.23.4767.
6.Csibra E, Brierley I, Irigoyen N.2014. Modulation of stop codon read-through efficiency and its effect on the replication of murine leukemia virus. J Virol88:10364 –10376.http://dx.doi.org/10.1128/JVI.00898-14. 7.Shehu-Xhilaga M, Crowe SM, Mak J.2001. Maintenance of the Gag/
Gag-Pol ratio is important for human immunodeficiency virus type 1 RNA dimerization and viral infectivity. J Virol75:1834 –1841.http://dx .doi.org/10.1128/JVI.75.4.1834-1841.2001.
8.Wills NM, Gesteland RF, Atkins JF.1991. Evidence that a downstream pseudoknot is required for translational read-through of the Moloney murine leukemia virusgagstop codon. Proc Natl Acad Sci U S A88:6991– 6995.http://dx.doi.org/10.1073/pnas.88.16.6991.
9.Dulude D, Berchiche YA, Gendron K, Brakier-Gingras L, Heveker N. 2006. Decreasing the frameshift efficiency translates into an equivalent reduction of the replication of the human immunodeficiency virus type 1. Virology345:127–136.http://dx.doi.org/10.1016/j.virol.2005.08.048. 10. Plant EP, Rakauskaite R, Taylor DR, Dinman JD.2010. Achieving a
golden mean: mechanisms by which coronaviruses ensure synthesis of the correct stoichiometric ratios of viral proteins. J Virol84:4330 – 4340.http: //dx.doi.org/10.1128/JVI.02480-09.
11. Jacks T, Madhani HD, Masiarz FR, Varmus HE. 1988. Signals for ribosomal frameshifting in the Rous sarcoma virusgag-polregion. Cell 55:447– 458.http://dx.doi.org/10.1016/0092-8674(88)90031-1. 12. Brierley I, Digard P, Inglis SC.1989. Characterization of an efficient
coro-navirus ribosomal frameshifting signal: requirement for an RNA pseudoknot. Cell57:537–547.http://dx.doi.org/10.1016/0092-8674(89)90124-4. 13. Baranov PV, Henderson CM, Anderson CB, Gesteland RF, Atkins JF,
Howard MT.2005. Programmed ribosomal frameshifting in decoding the SARS-CoV genome. Virology332:498 –510.http://dx.doi.org/10.1016/j .virol.2004.11.038.
14. Grentzmann G, Ingram JA, Kelly PJ, Gesteland RF, Atkins JF.1998. A dual-luciferase reporter system for studying recoding signals. RNA4:479 – 486.
15. Loughran G, Firth AE, Atkins JF.2011. Ribosomal frameshifting into an overlapping gene in the 2B-encoding region of the cardiovirus genome. Proc Natl Acad Sci U S A108:E1111–E1119.http://dx.doi.org/10.1073 /pnas.1102932108.
16. Harrell L, Melcher U, Atkins JF.2002. Predominance of six different hexanucleotide recoding signals 3=of read-through stop codons. Nucleic Acids Res30:2011–2017.http://dx.doi.org/10.1093/nar/30.9.2011. 17. Napthine S, Yek C, Powell ML, Brown TDK, Brierley I.2012.
Charac-terization of the stop codon readthrough signal of Colorado tick fever virus segment 9 RNA. RNA18:241–252.http://dx.doi.org/10.1261/rna .030338.111.
18. Giedroc DP, Cornish PV.2009. Frameshifting RNA pseudoknots: struc-ture and mechanism. Virus Res139:193–208.http://dx.doi.org/10.1016/j .virusres.2008.06.008.
19. Firth AE, Wills NM, Gesteland RF, Atkins JF.2011. Stimulation of stop codon readthrough: frequent presence of an extended 3=RNA structural element. Nucleic Acids Res39:6679 – 6691.http://dx.doi.org/10.1093/nar /gkr224.
20. Namy O, Moran SJ, Stuart DI, Gilbert RJC, Brierley I. 2006. A mechanical explanation of RNA pseudoknot function in programmed ribosomal frameshifting. Nature 441:244 –247. http://dx.doi.org/10 .1038/nature04735.
21. Gao F, Simon AE.2016. Multiplecis-acting elements modulate pro-grammed⫺1 ribosomal frameshifting in Pea enation mosaic virus. Nu-cleic Acids Res44:878 – 895.http://dx.doi.org/10.1093/nar/gkv1241. 22. Ritchie DB, Foster DAN, Woodside MT.2012. Programmed⫺1
frame-shifting efficiency correlates with RNA pseudoknot conformational plas-ticity, not resistance to mechanical unfolding. Proc Natl Acad Sci U S A 109:16167–16172.http://dx.doi.org/10.1073/pnas.1204114109. 23. Green L, Kim C-H, Bustamante C, Tinoco I, Jr.2008. Characterization
on November 7, 2019 by guest
http://jvi.asm.org/
of the mechanical unfolding of RNA pseudoknots. J Mol Biol375:511– 528.http://dx.doi.org/10.1016/j.jmb.2007.05.058.
24. Moomau C, Musalgaonkar S, Khan Y, Jones J, Dinman J.2016. Struc-tural and functional characterization of programmed ribosomal frame-shift signals in West Nile virus strains reveals high structural plasticity amongcis-acting RNA elements. J Biol Chem291:15788 –15795.http://dx .doi.org/10.1074/jbc.M116.735613.
25. Houck-Loomis B, Durney MA, Salguero C, Shankar N, Nagle JM, Goff SP, D’Souza VM. 2011. An equilibrium-dependent retroviral mRNA switch regulates translational recoding. Nature480:561–564.http://dx.doi .org/10.1038/nature10657.
26. Barry JK, Miller WA.2002. A⫺1 ribosomal frameshift element that requires base pairing across four kilobases suggests a mechanism of regu-lating ribosome and replicase traffic on a viral RNA. Proc Natl Acad Sci U S A99:11133–11138.http://dx.doi.org/10.1073/pnas.162223099. 27. Paul CP, Barry JK, Dinesh-Kumar SP, Brault V, Miller WA.2001. A
sequence required for⫺1 ribosomal frameshifting located four kilobases downstream of the frameshift site. J Mol Biol310:987–999.http://dx.doi .org/10.1006/jmbi.2001.4801.
28. Tajima Y, Iwakawa H, Kaido M, Mise K, Okuno T. 2011. A long-distance RNA-RNA interaction plays an important role in programmed ⫺1 ribosomal frameshifting in the translation of p88 replicase protein of Red clover necrotic mosaic virus. Virology417:169 –178.http://dx.doi.org /10.1016/j.virol.2011.05.012.
29. Brown CM, Dinesh-Kumar SP, Miller WA. 1996. Local and distant sequences are required for efficient readthrough of the barley yellow dwarf virus PAV coat protein gene stop codon. J Virol70:5884 –5892. 30. Cimino PA, Nicholson BL, Wu B, Xu W, White KA.2011. Multifaceted
regulation of translational readthrough by RNA replication elements in a tombusvirus. PLoS Pathog7:e1002423.http://dx.doi.org/10.1371/journal .ppat.1002423.
31. Newburn LR, Nicholson BL, Yosefi M, Cimino PA, White KA.2014. Translational readthrough in Tobacco necrosis virus-D. Virology450: 258 –265.http://dx.doi.org/10.1016/j.virol.2013.12.006.
32. Cornish PV, Hennig M, Giedroc DP.2005. A loop 2 cytidine-stem 1 minor groove interaction as a positive determinant for pseudoknot-stimulated⫺1 ribosomal frameshifting. Proc Natl Acad Sci U S A102: 12694 –12699.http://dx.doi.org/10.1073/pnas.0506166102.
33. Su MC, Chang CT, Chu CH, Tsai CH, Chang KY.2005. An atypical RNA pseudoknot stimulator and an upstream attenuation signal for⫺1 ribosomal frameshifting of SARS coronavirus. Nucleic Acids Res33:4265– 4275.http://dx.doi.org/10.1093/nar/gki731.
34. Hu H-T, Cho C-P, Lin Y-H, Chang K-Y.2016. A general strategy to inhibiting viral⫺1 frameshifting based on upstream attenuation duplex formation. Nucleic Acids Res44:256 –266.http://dx.doi.org/10.1093/nar /gkv1307.
35. Cho C-P, Lin S-C, Chou M-Y, Hsu H-T, Chang K-Y.2013. Regulation of programmed ribosomal frameshifting by co-translational refolding RNA hairpins. PLoS One 8:e62283. http://dx.doi.org/10.1371/journal .pone.0062283.
36. Carrington JC, Heaton LA, Zuidema D, Hillman BI, Morris TJ.1989. The genome structure of Turnip crinkle virus. Virology170:219 –226. http://dx.doi.org/10.1016/0042-6822(89)90369-3.
37. Hacker DL, Petty ITD, Wei N, Morris TJ.1992. Turnip crinkle virus
genes required for RNA replication and virus movement. Virology186: 1– 8.http://dx.doi.org/10.1016/0042-6822(92)90055-T.
38. Honigman A, Wolf D, Yaish S, Falk H, Panet A.1991.cis-acting RNA sequences control thegag-poltranslation readthrough in murine leu-kemia virus. Virology 183:313–319. http://dx.doi.org/10.1016/0042 -6822(91)90144-Z.
39. Feng YX, Yuan H, Rein A, Levin JG.1992. Bipartite signal for read-through suppression in murine leukemia virus mRNA: an eight-nucleotide purine-rich sequence immediately downstream of thegag ter-mination codon followed by an RNA pseudoknot. J Virol66:5127–5132. 40. Liu H, Naismith JH.2008. An efficient one-step site-directed deletion, insertion, single and multiple-site plasmid mutagenesis protocol. BMC Biotechnol8:91.http://dx.doi.org/10.1186/1472-6750-8-91.
41. McCormack JC, Simon AE.2006. Callus cultures ofArabidopsis. Curr Protoc Microbiol Chapter 16:Unit 16D.1. http://dx.doi.org/10.1002 /9780471729259.mc16d01s00.
42. Chattopadhyay M, Stupina VA, Gao F, Szarko CR, Kuhlmann MM, Yuan X, Shi K, Simon AE.2015. Requirement for host RNA-silencing components and the virus silencing suppressor when second-site muta-tions compensate for structural defects in the 3=untranslated region. J Virol89:11603–11618.http://dx.doi.org/10.1128/JVI.01566-15. 43. Wilkinson KA, Merino EJ, Weeks KM.2006. Selective 2=-hydroxyl
acy-lation analyzed by primer extension (SHAPE): quantitative RNA structure analysis at single nucleotide resolution. Nat Protoc1:1610 –1616.http://dx .doi.org/10.1038/nprot.2006.249.
44. McGinnis JL, Weeks KM.2014. Ribosome RNA assembly intermediates visualized in living cells. Biochemistry53:3237–3247.http://dx.doi.org/10 .1021/bi500198b.
45. Spitale RC, Crisalli P, Flynn RA, Torre EA, Kool ET, Chang HY.2013. RNA SHAPE analysis in living cells. Nat Chem Biol9:18 –20.http://dx.doi .org/10.1038/nchembio.1131.
46. Simon AE, Gehrke L.2009. RNA conformational changes in the life cycles of RNA viruses, viroids, and virus-associated RNAs. Biochim Biophys Acta1789:571–583.http://dx.doi.org/10.1016/j.bbagrm.2009.05.005. 47. Gaudin C, Mazauric MH, Traikia M, Guittet E, Yoshizawa S, Fourmy
D.2005. Structure of the RNA signal essential for translational frameshift-ing in HIV-1. J Mol Biol349:1024 –1035.http://dx.doi.org/10.1016/j.jmb .2005.04.045.
48. Staple DW, Butcher SE.2005. Solution structure and thermodynamic investigation of the HIV-1 frameshift inducing element. J Mol Biol349: 1011–1023.http://dx.doi.org/10.1016/j.jmb.2005.03.038.
49. Low JT, Garcia-Miranda P, Mouzakis KD, Gorelick RJ, Butcher SE, Weeks KM.2014. Structure and dynamics of the HIV-1 frameshift element RNA. Biochemistry53:4282– 4291.http://dx.doi.org/10.1021 /bi5004926.
50. Alam SL, Wills NM, Ingram JA, Atkins JF, Gesteland RF.1999. Struc-tural studies of the RNA pseudoknot required for readthrough of the
gag-termination codon of murine leukemia virus. J Mol Biol288:837– 852.http://dx.doi.org/10.1006/jmbi.1999.2713.
51. Jones DS, Nemoto F, Kuchino Y, Masuda M, Yoshikura H, Nishimura S.1989. The effect of specific mutations at and around thegag-polgene junction of Moloney murine leukemia virus. Nucleic Acids Res17:5933– 5945.http://dx.doi.org/10.1093/nar/17.15.5933.