Copyright @ 1969 AmericanSocietyforMicrobiology Printed in U.S.A.
Simian Virus 40
Deoxyribonucleic
Acid
Replication
I.
Effect
of
Cycloheximide
onthe
Replication
of
SV40 Deoxyribonucleic
Acid
in
Monkey
Kidney Cells and in Heterokaryons of
SV40-transformed
and
Susceptible
Cells
SAULKIT, TAKASHI KURIMURA, RAMON A. DE TORRES, AND DEL R. DUBBS
Department of Virology andEpidemiology, Division of Biochemical Virology, Baylor University College of
Medicine, Houston,
Texas77025Received for publication 23 September 1968
Infectious deoxyribonucleic acid (DNA) wasextracted from greenmonkey kid-ney (CV-1) cultures atvarious times after the cultures wereinfected with simian
virus 40(SV40)atinputmultiplicities of 0.01 and 0.1plaque-forming unit (PFU)per
cell. Apronounced decrease in infectious DNAwasobserved from 3 to16hrafter virus infection, suggesting that structurally altered intracellular forms may have
beengenerated early in infection. Evidence is also presented thatSV40DNA
syn-thesis requires concurrent protein synthesis. DNA replication was studied in the presenceand absence ofcycloheximidein: (i)SV40-infected and uninfected cultures ofCV-1cells;(ii) culturessynchronized with1-3-D-arabinofuranosylcytosine(ara-C) for 24to30hrpriortotheaddition ofcycloheximide; and (iii) in heterokaryons of SV40-transformed hamster andsusceptible monkey kidney cells. DNA synthesiswas
determined bypulse-labeling the cultures with 3H-thymidineatvarious times from 24to46hr after infection. In addition, thetotal infectious SV40 DNAwasmeasured.
Addition of cycloheximide, even after early proteins had been induced, grossly
inhibited both SV40 and cellular DNA syntheses. The activities of thymidine kinase, DNApolymerase, deoxycytidylate deaminase, and thymidylate kinasewere
measured; theseenzymeactivitiesremained high foratleast9hrinthepresenceof
cycloheximide. SV40 DNA prelabeled with 3H-thymidine before the addition of cycloheximidewasalso relativelystable during the time required for cycloheximide
to inhibit further DNAreplication.
Thekinetics of simian virus 40 (SV40)
deoxy-ribonucleic acid
(DNA) replication
have been studiedby
anumberof methods:plaque
assayofinfectious
DNA,
radioautography,
and bandcentrifugation
or nitrocellulose chromatography ofpulse-labeled
DNA(16, 17, 19).
Thesestudies haveshown that SV40 DNAreplication begins
atabout 12 to 16hrand continues at a
rapid
rateuntil about44 hrafter virus infection.Incontrast, infectious virus
production
commences after a20-to24-hreclipseperiod and virus titers increase untilabout60 to 72 hrafter infection.Also, infec-tious SV40 DNA was first detected in hetero-karyons of transformed andsusceptible cells at19
hraftercellfusion,butwasnotdetected in trans-formedcells beforefusionorduringthe first 11 hr after fusion (19). Although the timing of SV40 DNAsynthesishasbeenelucidated,themolecular events whichoccur duringreplicationarepoorly understood. Presumably, these eventsinclude: (i) conversion of closed circular, double-stranded SV40 DNA (form I) to a "nicked" circle (form
II);
(ii) formation of complementary copies ofeach of the parental DNA strands; and (iii)
joining
ofends ofpolynucleotide chainsto form superhelical (form I) daughter molecules. It is possible thatsome ofthese processes take place atspecific cellular sites, perhapsthenuclear mem-brane (4, 22, 27).Two facets of SV40 DNA relication are de-scribedin thepresentstudy. First,it is shown that infectious SV40 DNA undergoes an
"eclipse"
soonaftervirusinfection ofgreenmonkeykidney (CV-1) cells. Secondly, we have asked whether synthesisofessentialproteins earlyininfection is a sufficientcondition for the
replication
of SV40 DNA late in infection, or whether there is an additional requirement for thesynthesis
of pro-teins concurrentwithreplication
ofSV40 DNA. Ourdata show that theadditionofcycloheximide eitherto cultures pretreated with1-f3-D-arabino-furanosylcytosine (ara-C) or to culturesnot pre-treated with ara-C, at timesafter early enzymes have been induced
by
virusinfection,
strongly inhibitsSV40 DNAreplication.
25
on November 11, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
Cellcultures and SV40 stocks. The cultivation of CV-1 and SV40-transformed hamster (TSV-5) cells hasbeendescribed (6, 17). SV40 virus was propagated and assayed in monolayer cultures of CV-1 cells.
Heterokaryonsof transformed TSV-5 andsusceptible
(CV-1) cells were produced by treating mixtures of cellswithultraviolet light-inactivatedSendai virus, as
described previously (6).
Extractionand assay of infectious SV40 DNA.
In-fectious SV40 DNA was extracted from cultures of infected cells andfromheterokaryonsoftransformed andsusceptible cellswithp-aminosalicylateandphenol (16). The infectivityof theDNA extracts was
deter-mined by plaque titration onmonolayer cultures of
CV-1cells (19).
Incorporation of 3H-thymidine (3H-dT) into DNA and separation of labeled SV40 DNA from cellular DNA.At various times after infection, cultures were
inoculated with0.1or0.2mlof3H-dT(NewEngland Nuclear Corp., Boston, Mass.). The cultures were
incubatedat37Cfor the times indicated in thelegends
to the tables and figures. Noninfected cultures were
treatedsimilarly,except that eithergrowthmediumor
spent medium (medium removed from 4- to 7-day-old cultures) was substituted for the virus prepara-tions. After the 3H-dT labeling period, the cultures wereharvestedas previously described (16), and the total DNAcontentandspecific activity oftheDNA
were determined on portions of the cellsuspensions
(18). Theremainder ofthe cellsweresedimented by centrifugation, washed with saline-glucose solution,
andusedforextraction ofcellular and viral DNA. To determine therelativeamountsof 3H-dTincorporated
intocellularand viralDNA, the nitrocellulose
chro-matography procedure of Kit et al. (16) and band
centrifugation in CsCl gradients (16, 31) were
em-ployed.
Enzyme assays. Assaysfor thymidine kinase, thy-midylatekinase,deoxycytidylate deaminase,andDNA polymerase were performed by standard methods employedinthislaboratory(15, 17, 20, 21).
RESULTS
Decrease in DNA
infectivity
early after SV40 infectionof CV-1 cells.When CV-1 cellswerepro-ductively
infectedwithSV40virus,
anincreasein infectiousSV40 DNAwasobserved,
beginning
atabout 12 to 16 hr after
infection,
and maximal titers ofDNAinfectivity
were attained at about 44 hr after infection.Theinitial increase ininfec-tious SV40 DNA took
place
at 12 to 16hr,
whether cells were infected at
high input
multi-plicities[10
to50plaque-forming
units(PFU)
per cell] orat lowinput
multiplicities (0.01
to 0.1 PFU/cell). Itapeared
fromprevious
experi-ments, however, that at low
input
multiplicities,
but not at
input
multiplicities
of 1PFU/cell
orgreater, theinfectious DNAcontentof cells was
less at 7.5 hr thanat2.5 hrorat 20 hr after infec-tion (19).
Theobserved decrease ininfectiousDNA early after SV40 infection suggested that structurally altered intracellular forms might begenerated in virus-infectedcells. As a first step in testing this idea, theearlyinfectionperiod was studied more closely. At low inputmultiplicities, a decreasein SV40 DNAinfectivity was observed by 6 hr after virus infection (Fig. 1). When the input multi-plicity was 0.1 PFU per cell, DNA infectivity diminishedby15.5 hr toalmost one-tenththat at 3 hr after infection. At the input multiplicity of 0.01 PFU/cell, the low levels of DNA infectivity from 6 to 15.5 hr after infection represented the limits of detectableSV40DNA. The data in Fig. 1 demonstrate that, early after virus infection, most of the parental SV40 DNA either is
con-105
2104
>\D 102
-_j
Z)
UL_ 102
0L
10~ ~ ~ M-0
lo
0 5 10 15 20 25 30 35
HOURS AFTER SV40 INFECTION
FIG. 1. Assay ofinfectious SV40 DNA at various times after infection ofCV-1 cells with intact virus.
Confluent monolayer cultures (107 cells per culture)
wereinfectedwith Imlof SV40atinputmultiplicities
of0.1and 0.01 PFU/cell. Afteran adsorptionperiod of2hr at 37 C, unadsorbedvirus wasremoved,20ml ofnew growth medium was added, and the cultures werefurtherincubatedat37C.Atthetimesindicated,
the cultures were trypsinizedand DNA extracts were preparedfrom the resulting cell suspensions (16, 19).
on November 11, 2019 by guest
http://jvi.asm.org/
[image:2.487.269.459.248.565.2]vertedto anoninfectious form or to aform not
extractable
by
thep-aminosalicylate
procedure.
[We
also considered the hypothesis that the loss of SV40DNAinfectivity
may haveresulted from the progressive loss of virus adsorbed at the cell surface which never penetrated andwas pro-gressively detached from the cells. To test this hypothesis, CV-1 cultures were infected with SV40 atinputmultiplicities
of about 20 and 0.2 PFU/cell, respectively. After a 2-hr adsorption period at 37C, the unadsorbed virus was re-moved, the cultures were rinsed four times, and 20mlof growth mediumwasadded. At2, 6, 10, and 16 hr. samples were removed from the me-dium and assayed for SV40. Contrary to the foregoinghypothesis,
the virus concentration in the supernatant media decreased progressively from 2 to 16 hr after infection. Thetotal PFU/ culture in thesupernatantmediawasineach case less than1O-3
that extracted from the cells in the culture at 16 hr after infection.]Effect of cycloheximide on the incorporation of 3H-dTinto DNA of noninfected andSV40-infected CV-1 cells. Tolearn whether concurrentprotein synthesis was required for
incorporation
of 3H-dT into DNA, the effect of cycloheximide was studied in nonsynchronized cultures and in cul-turesin which DNA synthesis was synchronized by pretreatment of cellswith
ara-C. A pulse-labeling experiment involvingara-C pretreatment isshown inFig. 2.SV40-infectedCV-1cells were incubated;with15,ugofara-Cper mlfrom2to24H3-dT INCORPORATION FOR 2 HRS.
'7r% r% INTO DNA n
z
a
c@
=L
hr after
infection.
"Mock-infected" control cul-tures weresimilarly
treated. It has been shown previously (14, 17,21)
that ara-C inhibits DNA synthesis and SV40capsid
protein formation,
but not theinduction ofearlyenzymes or SV40-tumorantigen.
Indeed,
the activities ofthymidine
kinase and DNA polymerase were greater in
either uninfected or
SV40-infected
monkey
kidney cultures treated with ara-C than in
un-treated cultures.
Theara-Cinhibition ofDNA
synthesis
canbe reversed byremoving
ara-C andadding
deoxycy-tidine. After the addition of
deoxycytidine,
a rapidsynthesis
of DNA was initiated(Fig. 2).
The
synthesis
ofDNA,
asmeasured eitherby
theincorporation
of tritium label for2hr intoDNA or by colorimetric measurement of the total DNA content per culture, was greater in SV40-infected cultures than in noninfected cultures. Whencycloheximide
was added to the cultures atthe time of thedeoxycytidine
reversal of the ara-Cblock,
3H-dTwasincorporated
intoDNA,
butat a
grossly
inhibitedrate.Similarresultswere obtained aftercycloheximide
additiontocultures not treated with ara-C.ThedatainFig.2suggestedthat
cycloheximide,
an inhibitor of
protein synthesis
(17), severely
curtails DNA
synthesis
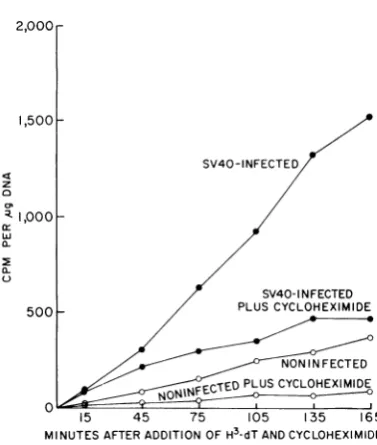
within 2 to 4 hr after addition ofthedrug.
Tolearnmoreprecisely
the time required to produce the cycloheximide in-hibition,theexperimentshowninFig.
3 was car-ried out. SV40-infected cultures of CV-1 cellsTOTAL ,ug DNA SV40- INFECTED
CV-I CELLS
200_
UNINFECTED _
CV-I CELLS t6_
-J
U120_
\ o _
<
80-TED z
L__---v D40
TED+CH
w
I
+1
u
SV40 -INFECTED CV- CELLS
UN NFECTED+CH INFECTED+CH
Ara-C
28 32 36 40 0 2 24 28 32 36
HOURS AFTER SV40 INFECTION
40
FIG. 2. Effectof cycloheximide (3,g/ml) ontheincorporation,for2hr,of3H-dTintoDNA ofSV40-infected
anduninfectedgreen monkeykidneycultures.Seven-day-old CV-J cells (9.3 X 10 cells/culture) were infected
with SV40at an inputmultiplicity of90PFU/cell. At 2 hrafterinjection, the unadsorbed virus wasremoved, andara-C(15
,g/mI)
wasadded. The cultureswereincubatedat37Cfor24 hrandwashed:50 ,ug of deoxycytidine perml, withorwithout3 ,ugof cycloheximideperml, wasadded. Cultureswereharvestedat the times indicatedin thefigure. At22, 24,29,and 35 hr after infection,0.1 mlof3H-dT(0.5,uc and2,ug/ml) was added to each
bottle.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.487.89.380.407.589.2]z
0
z 1,000
w
a-m
SV40-NFECTED
500
NONINFECTED
NOINEC-T
PLUSCYCLOHEXIMIDE15 45 75 105 135 165
MINUTES AFTER ADDITION OFH3-dT AND CYCLOHEXIMIDE
FIG. 3. Effect of'cycloheximide (3 jig/ml) on the kinetics of 3H-dT incorporation i)ito DNA of SV40-infected and uninfected CV-1 cell cultutres. Replicate CV-1 cell cultures (11.4 X JQ6 cells/culture) were
infected with SV40 at an inpuit multiplicity of' 50
PFU/cell. At 36 hrafter intkctio,i, 0.2 ml of3H-dT
(2,uc and2 jg/ml) wasadded. Some of the cultures were also treated with cycloheximide. Cultures were
harvestedatthetimesshownonthefigure.
wereincubated for 36 hrto
permit
theinitiationof viralDNA synthesis. 3H-dT wasadded torepli-cate cultures, either with or without cyclohexi-mide. Cultureswereharvestedatthetimesshown in Fig. 3.
Incorporation
of 3H-dT was severely inhibited within 1 hr ofcycloheximide
treatmentand only limited
incorporation
of 3H-dT into DNA occurred thereafter. The results suggested thataproteinsynthesized
denovoisratelimiting
for DNA
synthesis
in both uninfected and SV-40-infectedcultures.Separation oflabeled cellular and SV40 DNA. The
experiments
inFig.
3 show thatSV40 stimu-lated DNA synthesis in CV-1 cell cultures and that the synthesis of DNA in either infected oruninfected cultures was inhibited
by
cyclohexi-mide.Todeterminewhether bothSV40 and cellu-lar DNA syntheses were inhibited
by
cyclohexi-mide treatment, nitrocellulose
chromatography
and bandcentrifugation
experiments
werecarried out(Fig. 4,5).
Inthenitrocellulosechromatogra-phy experiment
(Fig. 4),
double-stranded DNArepresentsSV40DNAand
single-stranded
DNAis cellular DNA. As shown
previously
(16),
SV 40infection stimulatescellularDNAsynthesis
in CV-1 cells 36 to 44 hr after infection.Cyclo-heximide treatment prevents this stimulation of
cellular DNA synthesis and also markedly re-duces SV40 DNA synthesis. The band centrifuga-tion experiment(Fig. 5) further demonstrated the inhibitoryeffect of cycloheximide on both cellular DNA synthesis (fractions 1 to 22) and on SV40 DNA synthesis (fractions 23 to 30). Similar results were obtained when the 3H-dT labeling periods were34 to 38 hr, 40 to 44 hr, or 42 to 46 hr afterinfection.
Stability of "pulse-labeled" SV40 DNA during
[image:4.487.67.256.64.287.2]a "chase" period with or without cycloheximide. We considered the possibility that SV40 DNA synthesized during cycloheximide inhibition might beunstable. This instability could, perhaps, result from afailure to synthesize essential "maturation" proteins or capsid proteins in the presence of the drug. The "pulse-chase" experiment depicted in Table 1 contradicts this hypothesis. Infected and uninfected cultures of CV-1 cells were labeled with :H-dTfrom 36 to 40 hr after infection. The cultures were then washed, and excess "cold" thymidinewasaddedeither with or without cyclo-heximide. Cultures were further incubated at 37 C and harvested at 44 and 48 hr. The per cent tritium counts in cellular and viral DNA were
800r 700
O 600
w 500
z, 400
Ir-I X. 300
a. z
o o 200 Z Z)
-J 40
H
H3dT CH H3dT+CH
FIG.4. Estimationby nitrocellulose column
chroma-tography ofthe relative amounts ofSV40 DNA and cellular DNA synthesized in uninfected and
SV40-in,fected CV-I cellcultures.DNA samples wereheated
at100Cfor10mm in1 X SSCbeforechromatography. Single-stranded (SS) DNA is the denatured DNA
which requires 0.1 X SSC and 0.01 N NaOH,for
elution. Double-stranded (DS) DNA is that DNA which is not denatured by the heating and is eluted from the columns with 2 X SSC (circular SV40 DNA). The intervalsof3H-dTincorporationintoDNA
are shown below each column. Replicate cultures of
7-day-old CV-1 cells (107 cells/culture) were infected
with SV40 at an input multiplicity of100PFU/cell.
iuuI
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.487.269.469.338.527.2]EFFECT OF CYCLOHEXIMIDE ON SV40 DNA
0
P
400-tz. NONINFECTED, NONINFECTED, SV40-INFECTED, 300 WITHCH NOCH WITHCH
200 36-40hrs.PI 6-40hrs. PI
100
5 9 13 17 21 25 29 33 37 41 43
FRACTION NUMBER
FIG. 5. Sedimentation-velocity analysis (band centrifugation with CsCJ, density 1.503 gcm-3,
dis-solved in 0.01 M tris(hydroxymethyl)aminomethane
buffer, pH 8) ofDNA from uninfected or
SV40-in-fectedCV-1 cellslabeledwith3HdTfrom36to40 hr after infection. DNA samples were from the same
experimentshown inFig. 4. Alamellaof 0.1 ml,
con-taining 3 gg of DNA, was added to each tube. Replicate cultures were treated with 3 jg of cyclo-heximideper mlat the time of 3H-dT addition. The centrifugation was for 150 min at 20Cand100,000 X g in the SW39 rotor of the Spinco L2
cen-trifuge.
determined by nitrocellulose column chroma-tographyand bandcentrifugationinCsCl density gradients. Thetotal 3H-dTin theDNA of both the uninfected and the infected cultures was
about the sameafter the 8-hr chaseperiodasat
40hr.Cycloheximidetreatmentdidnotaccelerate breakdown of either cellularorSV40 DNA. Thus,
the inhibition of 3H-dTincorporation into DNA causedbycycloheximidecannotbe ascribedtoan
enhancement of DNA turnover.
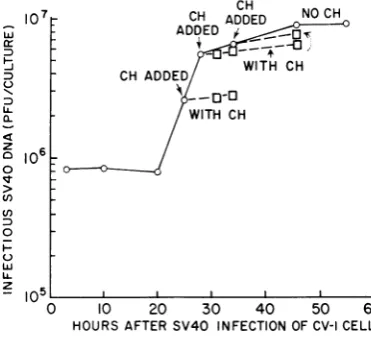
Effect of cycloheximide on the replication of
infectiousSV40 DNA. The experimentsin Fig. 2
through 5 depict the effect of cycloheximide on
the "pulse-labeling" of SV40 DNA. The total
SV40 DNA accumulated can be determined by
measuring the infectious SV40 DNA at various times after infection. Figure 6 shows the effect of cycloheximide treatment on this process. Cultureswerefirst incubated for 25 hrormoreto
permitthe formationof"earlyproteins"andthe initiation of DNA replication. Cycloheximide
was added toreplicate cultures 25, 28, or 34 hr after infection. Cycloheximide treatment in-hibited the formation of infectious SV40 DNA, but did not induce loss of DNA infectivity. Ex-perimentswerealsocarriedoutinwhichCV-1 cell cultures were first treated with ara-C to permit formationof "earlyproteins" andtosynchronize
w
tL3
0L.
z
0
0
c)
Uf)
0
0
w
z
io7I
6
In5
CH ADDED NO CH
ADDEDDE
CH ADDED WITH OH
WITH OH
:~~~~~ 0
0 10 20 30 40 50 60
[image:5.487.34.227.66.230.2]HOURS AFTERSV40 INFECTIONOFCV-I CELLS
FIG. 6.Effectofcycloheximide (CH)on the
forma-tionof infectious SV40 DNAin CV-1cultures. Seven-day-old CV-1 cell cultures (11 X 106 cells/culture)
were infected with SV40 at an input multiplicity of 20 PFU/cell. Cycloheximide (3 ,ug/ml) was added
at 25, 28, and 34 hr, and cultures were harvestedat
thetimesindicatedinthe figure.
TABLE1.Stability ofSV40 DNA labeledfrom36 to 40 hrpostinfection (PI) duringsubsequentincubation
of infectedcellsfrom40 to48 hr PI with and withoutcycloheximide (CH)a
Totalcountsper min perculture
Total counts/min for SV40 DNA Time of cell) Noninfected SV40-infected
NoCH With CH NoCH WithCH NoCH WithCH
40 5 X 105 1.59 X 106 8.6 X 105
44 5.7 X 105 5 X 105 1.66 X 106 1.48 X 106 8.6 X 105 8.6 X 105
48 5.3 X 105 4.8 X 105' 1.27 X 106 1.15 X 106 7.6 X 105 6 X
10'
aSeven-day-oldCV-1 cells (9.4 X 106cells/culture) were infected with SV40 at an input multiplicity of100PFU/cell. Replicatecultures wereincubated with 3H-dT (2 ,uc and 0.25,g/ml) from 36 to 40 hr PI. At 40 hrPI, cultureswerewashed,20,ugof nonradioactive thymidine per ml was added, and cultures
werefurtherincubated at 37C.Cycloheximide (3,ug/ml) was added at 40 hr PI to the infected and un-infected cultures.
VOL. 3, 1969
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.487.242.428.248.417.2]In the preceding experiments, cycloheximidewas added at a time when enzymes in the DNA bio-synthetic pathway had already attained high levels. We considered the hypothesis that one of
NO CH the enzymesmightundergo rapidturnoverduring
the arrest of
protein
synthesis
and that thisen-- CH ADDED WITH CH zyme deficiency might restrict DNA synthesis.
+C______- - ( - Four ofthe enzymes catalyzing nucleotide inter-conversions and DNA replication were therefore studied, namely, thymidine kinase, thymidylate WITH CH> - kinase, deoxycytidylate deaminase, and DNA
/ _-~ polymerase. In some instances, cultures were
pre-treatedfor 30 to 32 hr with ara-C before the addi-tion of cycloheximide. The results of these
experi--
--
ments failed to substantiate the hypothesis that- l
cycloheximide
initiated arapid
loss of enzymeCH ADDED activities. Thus, in
SV40-infected
cultures treated-3,ug/ml) with cycloheximide at 36 hr, the activities of
(3,ug/ml)Ithymidine kinase, deoxycytidylate deaminase, and
thymidylate kinase were as high at 42 hr as in culturesnottreatedwith the
drug. Similarly,
when!O 30 40 50 60 70 80 cycloheximide was added at 38 hr, the activities of
IOURS AFTER FUSION OF TSV-5 a CV-I CELLS thymidine kinase and DNA polymerase were
* 7. Effectofcyclohexinmicle (CH) on thetorma- about as high with cycloheximide as without drug )J infectious SV40 DNA in heterokaryons of treatment at 45 hr after infection. An experiment
9rmed TSV-5 and susceptible CV-1 cells. Mix- in which uninfected and SV40-infected cultures
)f CV-1 (107 cells) antd TSV-5 (5.8 X 106 cells) werepretreated with ara-C for 30 hr before the
treated
wit/i
14,000hemagglutinlatinig untits
of addition of cycloheximide is shown in Table 2.olet-inactivated Sendai virus to promote fusion The cultures exhibited high levels of thymidine 4mixture
equivalenit
to2X 101 CV-1 and 1.2 X kinase activity for at least 9 hr aftercyclohexi-V-5 cells was theni seeded inprescriptioni bottles mide was added. anid inzcubatedat 37 Cfbrthe times inidicated.
Cyclo-heximide (3 ,ug/ml) wasadded at 29 or at48hrafter
fusioni.
subsequent SV40 DNA
replication.
Ara-C wasremoved and
deoxycytidine
wasadded24 hrafter infection.Cycloheximide
was added toreplicate
cultures at 28 and 38 hr after infection. In this case, also,
cycloheximide
treatment inhibited infectious SV40 DNAsynthesis.
However, for 8 hr aftercycloheximide addition,
there was noappreciable
decrease of infectious SV40 DNA. These experiments further demonstrated that SV40 DNA isrelatively
stable for several hours after protein synthesis is inhibited.Experimentswerealsoperformedonthe forma-tion ofinfectious SV40DNAinheterokaryonsof susceptible CV-1 and transformed TSV-5 cells. Addition ofcycloheximide to the cultures at 29 or 48 hr aftercell fusion also retarded the
rapid
formation of infectious SV40 DNA in these cul-tures (Fig. 7).
Effect of cycloheximide treatment on enzyme activities. The induction of enzymes by SV40 is
first detected at about 12 to 16 hr after virus infection, and maximal enzyme activities are
[image:6.487.61.250.54.310.2]usuallyobserved
by
about30 hr(14,
17,19,
21).
TABLE 2. Thymidinie kiniase activities ofuninlfected
anidSV40-inifectedcultures ofCV-1cells treatedwith ara-Cfrom2 to30 hr PI
anidsubsequenitly treated with
cycloheximide (CH)a
Time Uninfected SV40-infected enzyme
assayed
(hrPI) NoCH withCII No CII W\ithCH
30 11.0 14.6
32 10.2 9.6 13.5 16.1
35 12.2 11.5 18.0 14.3
39 13.2 12.7 19.0 16.0
42 12.3 9.1 19.2 12.6
aEnzyme activity is expressed as picomoles of
3H-deoxyuridinemonophosphate formed from 3H-deoxyuridine per microgram ofprotein in 10 min
at 38 C. Seven-day-old cultures of CV-1 cells (11 X 106 cells/culture) were infected with SV40
at an input multiplicity of83 PFU/cell. Cultures
wereincubated with20,ugofara-C perml from2to
30 hrafterinfection,themediawerechanged,and
50 ug ofdeoxycytidine per ml, and in some cases
3,ug ofcycloheximide per ml,wasadded. Cultures
wereharvestedatthetimes indicated.
104
7
5
3 2
Q 103
J 5
cr
H
a-- 2
z
02
0 T 7
n 5 3 2
lo'l
2 H
FIG tion o
tranisfc
turesc
were I
ultravi( (19).A
106TS
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.487.264.458.412.553.2]DISCUSSION
The present experimentshave shown thatSV40 DNA infectivity undergoesaneclipse early after
virus infection of CV-1 cells. This finding is
reminiscent of the observation that phage X
DNA extracted from cells soon after infection
retainsverylittleinfectivityintheKaiser-Hogness transformation assay due to the formation of
structurallyalteredintracellularforms(3, 5, 32). It ispossiblethat the loss ofSV40DNAinfectivity also results from the formation of structurally altered intracellular forms. The DNA of
(pX174
RF, which resemblesSV40DNA in conformation
and molecularweight,is known toundergo
struc-tural changes during replication (22). Parental
spX174-RF molecules are attached to a
fast-sedimenting cell component; the "attached"
RF is foundinpartasRFIIand inpartasRFI.
Only attached RF molecules in the RFIH form
(nicked-circle) replicate, anddaughterRF
mole-culesarelater found free in thecytoplasmofthe
bacterialcell.
We recognize, however, that the loss ofSV40 DNA infectivity may havea less interesting
ex-planation. Perhaps this loss results from
intra-cellular nuclease action which occurs after virus
particles are "uncoated" and while the SV40 DNA is "in transit" to replication sites.
Addi-tional studies, including experiments utilizing labeled virus, arerequired to ascertain the
mol-ecular conformation of the noninfectious SV40 DNA and its intracellular localization, and to
learn whether it is, indeed, an obligatory
inter-mediate inSV40DNAreplication.
Thepresentexperimentshave also shownthat
protein synthesis must occur concurrently with
SV40 DNA replication in productively infected
CV-1 cellsand in heterokaryonsof transformed andsusceptiblecells. It isnot knownwhetherthe
requisite proteins are viral-specific or cellular
proteins. This studyand otherinvestigations (25, 28, 30) have shown that drugs which prevent
protein synthesisinhibitthe acceleration of
cellu-larDNAsynthesis insynchronized cellcultures;
it is conceivablethata specificclass ofhost pro-teins isrequiredfor bothcellular andviral DNA
replication.
From thetemporalpoint ofview, the proteins requiredduring SV40DNAreplicationare"late"
proteins. Thatis, ininfected culturesnot treated withara-C, they arerequired severalhours after
progenySV40DNAandcapsid protein syntheses
have beeninitiated.Also,inculturessynchronized with ara-C, the new class ofproteins is needed after the "early" class of proteins has
accumu-lated. Data which suggest a stoichiometric
rela-tionshipbetween DNA andprotein synthesis have
been obtained in several microbial and mam-maliansystems(23-25, 28). Invirus-infected cells, it has been shown that protein synthesis late in phage development is necessary
for
the produc-tion ofinfectiousX DNA (5). Thesynthesis of a histidine-containing protein is necessary for the replication of the parental replicative form ofspX
DNA(9).
Evidence
hasbeenpresented
that poxvirus DNA synthesis requires concurrent protein synthesis (1, 13). Moreover, although the enzymescatalyzing
theincorporation
of thy-midine intoDNA in cellularextractsfrom pseu-dorabies-infected cellsarestable after addition of puromycin, DNA synthesis in vivo is rapidly inhibitedby
addition of the drug (2).The nature and functions of the proteins re-quired for SV40DNAreplication havenot as yet been elucidated. Some possibilities that may be considered arethattheproteins are: (i) enzymes
exhibiting
markedinstability
during inhibition of protein synthesis; (ii) proteins which stabilize progenySV40DNA;
or(iii)
proteins correspond-ing to the structural and initiator proteins (24, 26) postulated by Jacob et al. (11). Studies on the activities of the thymidine kinase, DNA polymerase,deoxycytidylate
deaminase, andthy-midylate
kinase of infectedcells
indicate that these enzymeactivities
are relatively stable for several hours after cycloheximide treatment. It is,therefore, unlikely thatone of these enzymes is the essential protein. However, these results do notnecessarily
excludearoleforanother enzymefunctioning
in DNA biosynthesis. The data in Table 1 and those in Fig. 6 and 7 also suggest that the required protein is not one needed to stabilize SV40 DNA,since
progeny DNA is relatively stable for several hours aftercyclo-heximide
addition. The third possibility, that protein synthesis is required eitheras astructural componentofanSV40 DNAreplicationmachine oras aninitiator protein, merits furtherstudy (11, 22, 24, 26). Data supportingthe hypothesisthat DNAreplication
mightoccur in direct association withmembranestructureshavebeen obtainedfor the (pX174RF (22) and for microbial(8, 12) and mammaliancell systems (4,27).Therateofcellu-lar DNA
replication
is about2pm/min
at 37 C for mammalian cells(10,
29) and variesfrom 6 to40,tm/min
for microbialcells,
dependingupon the growth media (7). Since SV40 DNA is less than 2,umlong,it isprobablethatonce,initiated, around ofreplication requiresonly afew minutes. The time intervals studied in the experiments shown inFig.
2and 3aremuchlongerthan this. Thus, the observed inhibitions may reflect the failure to fabricate new viral DNAreplication
complexes, while existing replication complexes
on November 11, 2019 by guest
http://jvi.asm.org/
may continuetofunction. This may permit con-tinued synthesis of SV40 DNA atreduced rates.
ACKNOWLEDGMENTS
Thisinvestigationwassupportedby National Science
Founda-tion grant GB5917, American Cancer Society grant E-291E,
Robert A. Welch Foundation grant Q-163, and Public Health ServicegrantsCA-06656-06,1-K6-AI-2352,and5-K3-CA-25,797. WethankMarjorie Johnson, CarolynSmith,JudithRotbein,
and Sun Ock Yim for able technical assistance. LITERATURE CITED
1. Bedson,H.S., and J.G.Cruickshank. 1968.Multiple protein
functions in the replication ofpox virus DNA. J. Gen. Virol. 3:147-151.
2. Ben-Porat,T.,and A.S.Kaplan.1966. Thereplicationofthe double-stranded DNA ofananimalvirusduring intracellu-larmultiplication.Intern.Congr.Microbiol.,9th, Moscow, p.463-482.
3. Bode, V.C., andA. D.Kaiser.1965.Changesin thestructure andactivityofXDNA inasuperinfectedimmunebacterium. J. Mol. Biol. 14:399-417.
4. Comings, D. E., andT.Kakefuda. 1968. Initiation of deoxy-ribonucleicacid replication at the nuclear membrane in human cells.J.Mol.Biol. 33:225-229.
5. Dove, W.F., andJ. Weigle. 1965. Intracellularstate of the chromosome ofbacteriophage lambda. I. The eclipse of
infectivityof thebacteriophageDNA.J.Mol.Biol. 12:620-629.
6. Dubbs,D.R., andS.Kit. 1968.Isolation of defectivelysogens
from simian virus 40-transformed mousekidney cultures. J. Virol.2:1272-1282.
7. Eberle, H.,andK.G.Lark.1967. Chromosomereplicationin Bacillus subtilis culturesgrowing at different rates. Proc. Natl. Acad.Sci. U.S. 57:95-101.
8. Ganesan, A.T., and J. Lederberg. 1965. A cell membrane boundfraction of bacterialDNA.Biochem.Biophys.Res. Commun.18:824-835.
9. Greenlee,L.L., andR. L.Sinsheimer. 1968. Theprocessof infection withbacteriophage(pX174.XVII.Effects ofspecific
metabolicinterruptions.J.Mol.Biol. 32:303-332. 10. Huberman, J.A., andA. D.Riggs. 1968. On the mechanism
ofDNAreplication in mammalian chromosome. J. Mol. Biol.32:327-341.
11. Jacob,F.,S.Brenner,andF.Cuzin. 1963. On theregulation
ofDNAreplicationinbacteria.ColdSpringHarborSymp.
Quant. Biol. 28:329-348.
12.Jacob, F.,A.Ryter, andF.Cuzin. 1966.Ontheassociation betweenDNA and membranein bacteria.Proc.Roy.Soc.
(London)Ser. B164:267-278.
13. Kates,J.R., andB. R.McAuslan.1967.Relationshipbetween
proteinsynthesisandviraldeoxyribonucleicacidsynthesis.
J.Virol.1:110-114.
14. Kit,S. 1967. Induction ofenzymesof DNA metabolismby
simianvirus40,p.l16-143.InY.Ito(ed.), subviral carcino-genesis, First Intemational SymposiumonTumorViruses.
Chikusa-ku,Nagoya,Japan.
15. Kit, S., R. A. de Torres, and D. R.Dubbs. 1966. Arabino-furanosyl-cytosine-inducedstimulation of thymidine kinase and deoxycytidylate deaminase activities of mammalian cultures. CancerRes.26:1859-1866.
16. Kit, S., R. A. de Torres, D. R.Dubbs, and M. L. Salvi. 1967. Induction of cellular deoxyribonucleic acid synthesis by simian virus40. J. Virol. 1:738-746.
17. Kit, S., D. R. Dubbs, P. M.Frearson,and J. L.Meinick.1966. Enzyme induction inSV40-infected green monkey kidney cultures. Virology29:69-83.
18. Kit, S., D. R. Dubbs, and T. C.Hsu. 1963.Biochemistlyof vaccinia-infected mouse fibroblasts (strain L-M). HI. Radioautographicandbiochemical studies on thymidine Hs uptakeintoDNA of L-M and rabbit cells inprimaryculture. Virology19:13-22.
19. Kit, S., T. Kurimura, M. L. Salvi, and D. R. Dubbs. 1968. Activation of infectiousSV40DNAsynthesis in transformed cells. Proc.Natl. Acad.Sci.U.S. 60:1239-1246.
20. Kit,S., J. L. Melnick, M. Anken, D.R. Dubbs, R. A. de Tor-res, and T. Kitahara. 1967. Nonidentity of some simian virus 40-induced enzymes with tumor antigen. J. Virol.
1:684-692.
21.Kit, S., L. J. Piekarski, and D. R. Dubbs. 1967 DNA polym-eraseinducedbysimianvirus 40. J. Gen. Virol. 1:163-173. 22. Knippers, R., and R. L. Sinsheimer.1968.Process of infection ofbacteriophageq,Xl74. XX. Attachment of the parental DNA of bacteriophage gX174 to a fast sedimenting cell
component. J. Mol.Biol. 34:17-29.
23. Lark, K. G. 1966. Regulation ofchromosome replication and segregation inbacteria. Bacteriol.Rev. 30:3-32. 24. Lark, K. G., and C. Lark. 1966. Regulationof chromosome
replication in Escherichia coli: acomparison of the effects ofphenethyl alcohol treatmient with thoseof amino acid starvation. J.Mol.Biol. 20:9-19.
25. Mueller,G. C., K.Kajiwara,E.Stubblefield. and R. R. Rueck-ert. 1962. Molecular eventsin the reproductionof animal cells.I.Theeffectof puromycinontheduplicationof DNA. CancerRes.22:1084-1090.
26. Porter, B., and D. K. Fraser. 1968. Involvement of protein andribonucleic acid in the association ofdeoxyribonucleic
acid with other cell components of Escherichia coli. J. Bacteriol. 96:98-104.
27. Sved, J. A. 1966. Telomere attachment ofchromosomes. Some genetical and cytological consequences. Genetics 53:747-756.
28. Taylor,E.W.1965.ControlofDNAsynthesisinmammalian cells inculture. Exptl. CellRes.40:316-332.
29.Taylor, J. H. 1968. Ratesofchaingrowthandunits of repli-cation in DNA of mammalian chromosomes. J. Mol. Biol.31:579-594.
30.Terasima, T., and M.Yasukawa. 1967. SynthesisofGI pro-teinprecedingDNAsynthesisinculturedmammalian cells. Exptl. Cell Res. 44:669-672.
31. Weil, R., M. R. Michel, and G. K. Ruschmann. 1965. In-duction of cellular DNAsynthesis by polyomavirus. Proc. Natl.Acad.Sci. U.S.53:1468-1475.
32.Young,E.T.,and R. L.Sinsheimer. 1964.Novelintra-cellular forms oflambda DNA. J. Mol.Biol. 10:562-564.