Collaborative Cross recombinant inbred inter-crosses (RIX) for the study of the behavioral and structural consequences of chronic antipsychotic treatment
Daniela DeCristo
Abstract
Schizophrenia is an idiopathic disorder that affects approximately 1% of the
global population, and presents with persistent delusions, hallucinations, and
disorganized behaviors. Antipsychotics are the standard treatment of schizophrenia, but
are frequently discontinued by patients due to inefficacy and/or intolerable side effects.
Chronic treatment with the antipsychotic haloperidol causes tardive dyskinesia in about
30% of patients, manifested in humans as involuntary and often permanent orofacial
movements. Tardive dyskinesia can be effectively modeled in rodents by means of
vacuous chewing movements. Recombinant inbred inter-crossed (RIX) mice, derived
from the genetically diverse Collaborative Cross murine population, were treated with
haloperidol and subjected to a panel of behavioral assessments to monitor the
development of side effects. Furthermore, transition electron microscopy (TEM) was
used to assess the impact of chronic haloperidol treatment on synaptic structures in
wild-type C57BL/6 mice. While there were similar neuronal synaptic architectures in the
corpus striata across treatment groups, distinctions were found in phenotypic behavior
between strain genotypes. These findings add to our understanding of the genetic basis
of the side effects of haloperidol with the aim of higher efficacy and lower burden of
haloperidol treatment.
Introduction
Schizophrenia is a highly complex and heritable psychiatric disorder with many
worldwide, schizophrenia is among the top ten leading causes of disability and is
accompanied with high costs for patients, their caretakers, and society as a whole1-4. Although schizophrenia was first described more than 100 years ago, an
underlying cause remains unknown. While genome-wide association studies (GWAS)
and structural variation studies have led to many advancements in our understanding of
the disorder in the past 5-10 years, its pathophysiology is still not well-defined.
Additionally, symptoms are often difficult to identify and can manifest in varying degrees
depending on the patient. As a result, doctors are tasked with correctly diagnosing
patients with schizophrenia using a vague clinical and research definition and an
imprecise therapeutic approach.
Recent innovations in genomics have shed light on the genetic basis of
schizophrenia. The Psychiatric Genomics Consortium, led by Dr. Patrick Sullivan,
applied GWAS using a large sample set of 36,989 schizophrenia patients and 113,075
unaffected individuals in order to determine possible candidate genes. The Consortium
found 108 loci in the human genome that confer an increased risk for the disorder. Within
this set of loci, there is an overrepresentation of genes with neuronal and brain
expression, including DRD25,6. DRD2 codes for the production of brain receptors for dopamine, a neurotransmitter highly involved in complex thought, movement, reward
and other behaviors. The gene is also the target of all clinically-effective antipsychotic
medications currently in use; these drugs antagonistically interact with dopamine
receptors to reduce hallucinations and delusions7.
Unfortunately, antipsychotic drugs frequently lead to serious adverse drug
no exception. Haloperidol is a prototypical antipsychotic and potent antagonist of the
dopamine receptor D2 in the striatum, a brain region that plans and moderates movement.
In addition to other side effects, a subset of patients will experience severe and often
irreversible motor ADRs including uncontrolled and purposeless jaw movements, termed
tardive dyskinesia (TD)8. Prevalent in about 30% of patients and permanent in half of those affected, the reason why haloperidol causes this ADR has not been discovered9-11. Currently, there is no compelling way to predict adverse drug reactions (ADRs) or drug
efficacy using the genetic makeup of a patient, an ability that would make drug treatment
of schizophrenia using antipsychotics safer and more effective.
The laboratory mouse can be used as a proxy to study certain human
pharmacogenetic phenotypes. Collaborative Cross RIX mice recapitulate many of the
features of TD with haloperidol treatment including jaw tremors, tongue protrusions, and
vacuous chewing movements (VCMs), making them an ideal model system. A major
goal of the Center for Integrated Systems Genomics at the University of North Carolina
at Chapel Hill is to harness the genetic diversity of RIX mice to explore the genetic basis
of antipsychotic side effects12. This study is a further exploration of the preliminary data published by Crowley et al. that determined the validity of murine VCMs as a model of
TD induced by haloperidol and the heritability of risk of adverse drug effects10. Previous studies have explored morphological and synaptic changes that are
thought to be associated with haloperidol treatment and TD in the brain using rodents and
transition electron microscopy. Results from these studies are inconclusive and
contradictory at times. Kerns et al. suggested that alterations in synaptic architecture
observing an enhanced number of synaptic boutons and an increase in perforated and
double synapses due to haloperidol treatment13. In contrast, Benes et al. found that changes in synapse numbers were region-specific, and that striatal neuronal density and
relative number of nerve terminals were unchanged between treatment groups14. Additional studies have similarly reported an increase in perforated synapses in the
caudate nucleus and striatum13-17, yet no change was found in the nucleus accumbens or the medial prefrontal cortex in a separate study17. Other studies examining asymmetric synapses, or synapses that exhibit larger postsynaptic densities than presynaptic densities
and are typical of excitatory inputs, found decreases in asymmetric synaptic
densities16,18,19. Roberts and Lapidus suggested that this decrease in excitatory inputs may protect against VCMs in their examination of the relationship between haloperidol,
VCMs, and asymmetric synapses as a result of finding differentially affected asymmetric
synaptic densities in the low and high VCM groups, with a high amount of asymmetric
synaptic densities only in the high VCM group19.
The current problem at hand is that there is no compelling way to predict drug
efficacy or adverse drug reactions such as TD. The goal of this study is to elucidate the
genetic basis of TD by determining sensitivity to haloperidol-induced adverse drug
effects in the genetically diverse RIX mouse population. Synaptic morphology of striatal
tissue was also analyzed using electron microscopy due to the critical role of the striatum
in modulating motor function19. Behavioral phenotypes and synaptic ultrastructure were examined between mice treated with haloperidol and those treated with placebo. This
paper discusses findings from the behavioral characterization and synaptic architecture
TD susceptibility and contributes to the growing predictive power of pharmacogenomics
for improved personalized treatment of patients impacted by schizophrenia.
Methods
Animals
During the course of this study 846 male and female mice from 73 different RIX
lines derived from the Collaborative Cross (The Jackson Laboratory, Bar Harbor, ME)
were phenotyped. Mice were housed on a cycle of 12 hours of light and 12 hours of dark
with lights on at 0700 hours in a room held at a consistent temperature range of 20-24 °C.
Two mice of the same sex were housed per standard ventilated cage with water and
Purina ProLab IsoPro 3000 food available constantly and positioned on the roof of the
cage. For each strain, mice were grouped in three batches so that three replicate mice per
sex and treatment combination were tested by the end of the study. The study strictly
followed the ‘Guide for the Care and Use of Laboratory Animals’ with approval by the
Institutional Animal Care and Use Committee of the University of North Carolina20.
Haloperidol treatment and activity testing
Methods for drug administration were adopted from pilot studies indicating that
the optimal human-like steady-state concentration of haloperidol could be obtained using
a 30-day release tablet that delivered 3.0 mg kg-1 per day to the mouse10,21. Mice were anesthetized for 2 minutes with isoflurane, and haloperidol pellets were implanted into
the mice using a trocar22. The drug was administered after mice were 8 weeks old. The activity of the mice was monitored using open field testing in activity chambers with
infrared beams to measure locomotor and exploratory behavior of mice before treatment
Scoring orofacial movements
Video recording of vacuous chewing movements (VCMs) was carried out after 28
days post-treatment. To this end, mice were briefly anesthetized with isoflurane and
restrained for 25 minutes using a plastic collar. Collars were made from two plastic
semicircular pieces that were adjustable based on neck size and to achieve the most
comfortable position for the mouse. The collar partially immobilized the mice at the neck
but still permitted head movement to allow for video recording of jaw movements by
JVC Everio digital camcorders. Digital videotapes were made using the protocol
developed by Tomiyama et al.23. The first 10 minutes of video were not analyzed in order to allow the mice to adjust to the collar and to relax. The last 15 minutes of the video
were scored for orofacial movement. Videos were randomized and scored by a
single-blinded rater to increase consistency and to reduce any deviation or bias between raters.
The rater was trained by an expert and a set of standard training videos used in the study
by Crowley et al. to align the rater with correct identification of VCMs according to the
scoring from the previous study10. Drift was monitored by re-scoring random videos throughout the course of the study.
The movements that were specifically analyzed were tongue protrusions, jaw
tremors, overt chewing movements, and subtle chewing movements. Individual events of
each movement with the exception of tremors were counted; tremors were measured by
duration in seconds. Subtle chewing movements were defined as instances of vertical jaw
movement in which the inside cavity of the mouth could not be seen and the jaw was not
open for a long period of time. Overt chewing movements occurred when a larger vertical
extended length of time. The videos were scored using The Observer XT (Noldus Inc.,
Wageningen, Netherlands) observational data analysis program.
Behavioral analysis
Overall analysis of activity was performed using R (The R Foundation, Vienna,
Austria). Statistical analyses of scored observations were carried out using JMP software
(version 12.0.1, SAS Institute Inc., Cary, NC). Data from 846 RIX mice were collected
from 73 strains in 51 batches. Due to the very minor distinctions between subtle and overt
VCMs, the counts of these two phenotypes were combined for analysis.
The data for VCMs between treatment groups was fit following Two-Way
ANOVA with interaction:
Yijk = μ+αi+βj+γij+εijk
εijk ~ N(0,σ2) i = 1,…,ni
j = 1,…,nj
Where the α are strain effects, β are the treatment effects, and γ are the interactions
thereof. After dropping strain 5080x6750, as there was only a single mouse of this strain,
ni = 72, nj = 2, the model was fit in SAS PROC MIXED version 9.4. The following
hypothesis was then tested:
H0: γij = 0 ∀i,j
In words, the significance of the interactions - that is, whether the treatment effect differs
by strain – was tested. For this, the F-test was employed. Estimates of the treatment effect
for each strain were calculated with combined subtle and overt VCM totals as the
p-values, which have been adjusted to control False Discovery Rate at 0.05.
Tissue preparation
Using the same 30-day treatment protocol previously described, ten C57BL/6
mice were perfused with a solution of 2% paraformaldehyde/2.5% glutaraldehyde in 0.15
M sodium phosphate buffer, pH 7.4. A 2 mm thick coronal section (Bregma coordinates
from +1.0 to -1.0) was dissected and the ventral striatum (~1 mm thickness) was
manually isolated from each mouse. Samples were post-fixed in 1% osmium tetroxide in
0.15 M sodium phosphate buffer, pH 7.4, for one hour. Following gradual dehydration
with ethanol and propylene oxide, samples were infiltrated and embedded in PolyBed
812 epoxy resin (Polysciences, Inc., Warrington, PA). Light microscopy sections (1 µm)
were cut, mounted on slides, and stained with 1% toluidine blue O in 1% sodium borate.
After selecting the region of interest, 70 nm ultrathin sections were cut using a Leica
Ultracut UCT microtome (Leica Microsystems, Inc., Bannockburn, IL) and a diamond
knife. The sections were mounted on 200 mesh copper grids and contrasted with 4%
uranyl acetate and Reynolds' lead citrate stains.
Transition electron microscopy
Samples were observed using a LEO EM 910 transmission electron microscope at
80 kV (Carl Zeiss SMT, Inc., Thornwood, NY) and digital images were acquired with a
Gatan Orius SC1000 CCD camera and Digital Micrograph Software (version 2.3.1,
Gatan, Inc., Pleasanton, CA). With striatal samples from 10 different mice, 10 locations
per section were mapped to separate grids. Images were taken on a single plane to
eliminate any possibility of counting a synapse twice. For each location, one image was
without overlap. Four images per mouse were also acquired at 50,000 × for very close
examination of synapses.
Image analysis
Using Fiji, an open-source ImageJ software, synapses and mitochondria were
counted in each of the 100 electron microscopy images taken at 10,000 × using the cell
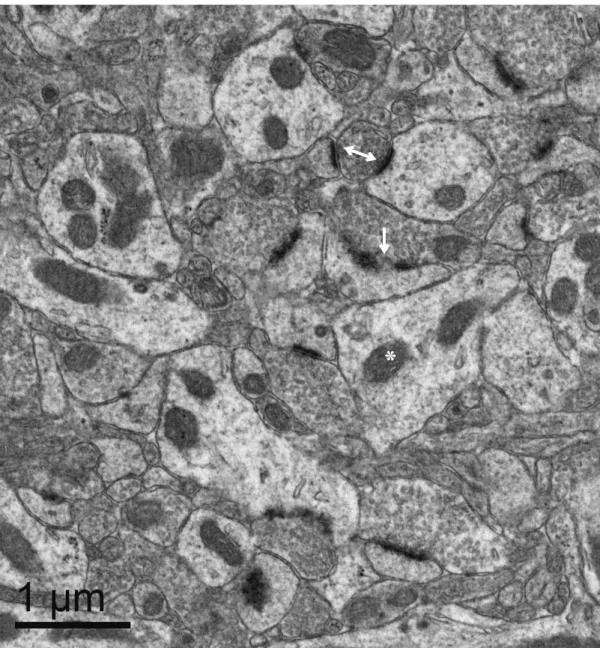
counter tool24. General observations of the amount of symmetric versus asymmetric synapses were made. Perforated synapses were defined as having breaks in synaptic
continuity greater than 0.05 μm and double synapses were those that exhibited active
zones with two different postsynaptic structures in accordance with similar previous
studies (Fig. 2)13,25. Perforated and double synapses were also counted in the 100 images at 10,000 ×. The scaling and measurement tools were used in Fiji to validate distances
between synaptic densities for perforated synapses.
Counts of the four measures of number of synapses, mitochondria, perforated
synapses and double synapses were totaled for each subject from their respective 10
images and evaluated using a Student’s t-test with an alpha of 0.0125 after Bonferroni
correction in JMP (version 12.0.1, SAS Institute Inc., Cary, NC).
Results
Behavior analysis
Overall open field activity analysis between treatment groups indicates reduced
activity in the mice treated with haloperidol in contrast to mice treated with placebo.
Haloperidol-treated mice spent less time exploring the open field both horizontally and
vertically. Levels of anxiety (as measured using time in center) and repetitive behaviors
VCMs analysis
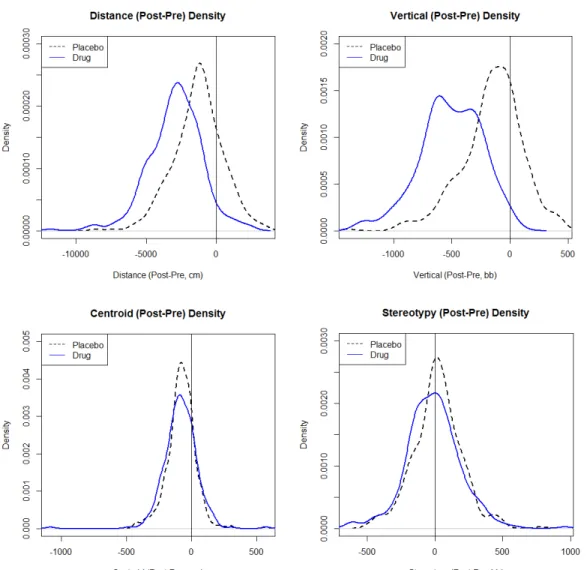
Haloperidol-treated mice showed greater susceptibility to subtle and overt VCMs
than placebo-treated mice with no significant differences between groups in number of
tremors and tongue movements (Fig. 4). Strain-by-strain analysis of the recorded number
of subtle, overt, tremors, and tongue protrusion VCMs indicate trends similar to the
overall trend observed between treatment groups (Fig. 5-8). From the Two-Way ANOVA
of 72 strains, the null hypothesis that there was no difference in treatment effect between
strains was rejected at the α = 0.05 level of significance with a calculated p-value of
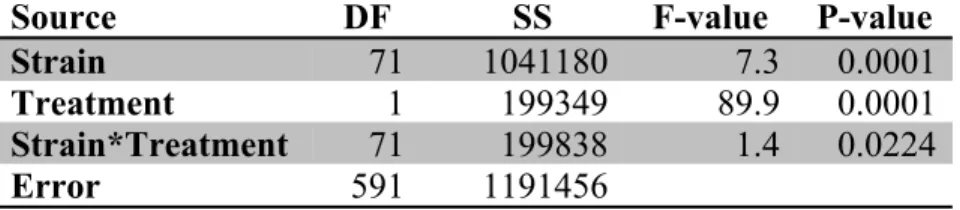
0.0224. Thus, the effect of treatment differs by strain (Table 1).
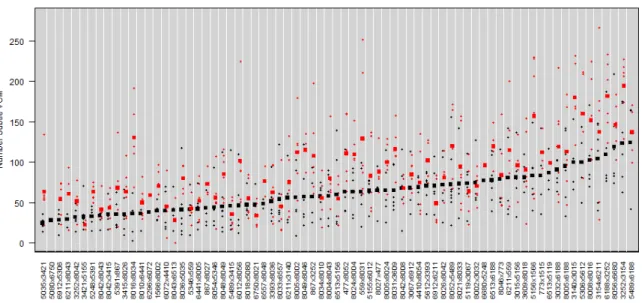
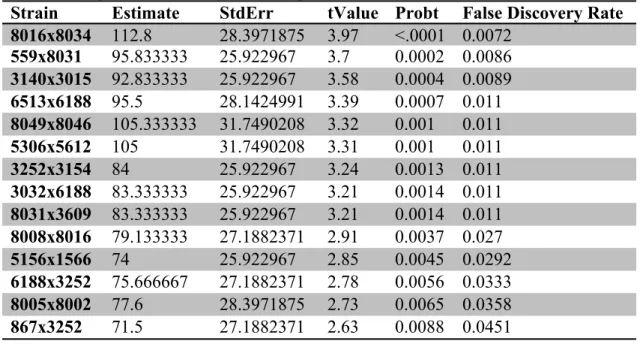
Individual statistical analysis of each strain allowed for clear identification of
strains significantly affected by haloperidol. In fourteen strains (8016x8034, 559x8031,
3140x3015, 6513x6188, 8049x8046, 5306x5612, 3252x3154, 3032x6188, 8031x3609,
8008x8016, 5156x1566, 6188x3252, 8005x8002, 867x3252), the number of combined
subtle and overt VCMs in mice treated with haloperidol were significantly different from
the number of VCMs recorded in mice administered placebo after controlling FDR at
0.05 (Table 2).
Striatal ultrastructure analysis
Most of the synapses observed in every image were asymmetric and had larger
post-synaptic densities than pre-synaptic densities. No differences were found between
30-day C57BL/6 treatment groups for the four measured ultrastructural components:
number of synapses, mitochondria, perforated synapses, and double synapses per sample.
Calculated p-values do not pass the Bonferroni-corrected threshold of significance of
Discussion
The Collaborative Cross-derived RIX lines have proven to be a vital means for
exploring heterogeneity. The results of this study validate the use of RIX mice for the
study of VCMs due to the differential haloperidol sensitivities observed in the strains
tested. The discovery of fourteen RIX strains that are highly susceptible to
haloperidol-induced TD contributes to the aim to identify genes regulating this susceptibility. Using
these lines diminishes the need for human studies with very large sample sizes required to
effectively parallel the diversity found in the entire population. Genetic mapping of
candidate regions identified in mice will allow precious human samples to be used only
for highly probable regions of interest as part of future research.
While significant differences were not found in the synaptic architecture between
drug and placebo treatment groups, increased power with a larger sample size or more
electron microscopy imaging may be necessary to reveal any differences or confirm
results. Additionally, images of another location in the striatum or at a different
time-point may also illustrate a more significant impact of the drug than what was captured in
this study. The results of this study suggest that the underlying mechanism of TD by
chronic haloperidol treatment does not involve alterations in striatal ultrastructure in
congruence with findings by Kessas et al.26. Further investigations of neuroplasticity, mitochondrial amounts, and synaptic counts will shed more light on this area of research.
The genetic variation found in the human population is difficult to model using
animals that have been bred to be genetically similar for many generations. These
difficulties arise when trying to solve problems and answer questions related to human
some of the smallest variances like single nucleotide polymorphisms and variable number
tandem repeats have been associated with significant phenotypic differences between
individuals. The Collaborative Cross murine population may be a valuable tool to assess
these genetic features.
Due to the genetic diversity and intricate breeding system of these Collaborative
Cross RIX lines, complex modeling of sex effects, effect of parental lines, within group
variability and between group variability is being carried out to examine the results in a
larger context and elucidate relationships between strains for genetic candidate mapping.
These efforts aim to better understand the relationship between gene expression and
chronic haloperidol treatment and to correlate phenotypes with strain genotypes and
RNA-sequencing data. Genome analysis will be done to try and identify regions of the
genome that correlate with those mice that reacted most strongly to the drug.
The recent popularity and intrigue of personalized medicine has generated much
discussion and research in health care. Evaluation of an individual patient’s genome,
familial genetic history, and pertinent environmental factors all to arrive at a precise
approach to treatment may lead to greater therapeutic success as this approach is refined.
The identification of genes involved in higher risk of TD development due to chronic
haloperidol treatment in RIX lines allows for focused application in human treatment to
increase physicians’ predictive power to prevent harmful side effects and maximize
benefit in the treatment of patients with schizophrenia.
Conclusions
RIX strains had wide-ranging sensitivities to haloperidol-induced VCMs, with
haloperidol does not influence synaptic amount or morphology. Further explorations of
changes in striatal neuroplasticity will clarify these results. This study will aid in the
discovery of the genetic determinants of susceptibility to tardive dyskinesia with more
in-depth analysis underway.
References
1 World Health Organization. The Global Burden of Disease: 2004 Update. Geneva:
WHO Press, 2008.
2 Murray CJL and Lopez AD. Alternative projections of mortality and disability by cause
1990-2020: Global Burden of Disease Study. Lancet. 1997;349(9064):1498-504.
3 Knapp M, Mangalore R, Simon J. The global costs of schizophrenia. Schizophrenia
bulletin 2004; 30: 279-293.
4 Saha S, Chant D, McGrath J. A systematic review of mortality in schizophrenia: is the
differential mortality gap worsening over time? Archives Gen Psych 2007; 64: 1123-31.
5 Sullivan P, Daly M, O’Donovan M. Genetic architectures of psychiatric disorders: the
emerging picture and its implications. Nat Reviews 2012; 13: 537-551.
6 Ripke, S. et al. Genome-wide association analysis identifies 13 new risk loci for
schizophrenia. Nature Genetics 2013; 45:1150-9.
7 Roth B, Sheffler D, Kroeze W. Magic shotguns versus magic bullets: selectively
non-selective for mood disorders and schizophrenia. Nature Reviews 2004; 3: 353-359.
8 Kinon B, Lieberman J. Mechanisms of action of atypical antipsychotic drugs: a critical
analysis. Psychopharmacology 1996; 124: 2-34.
9 Dayalu P, Chou KL. Antipsychotic-induced extrapyramidal symptoms and their
10 Crowley J, Adkins D, Pratt A, Quackenbush C, van den Oord E, Moy S, Wilhelmsen
K, Cooper T, Bogue M, McLeod H, Sullivan P. Antipsychotic-induced vacuous chewing
movements and extrapyramidal side effects are highly heritable in mice. Pharmacogen J
2010; 12: 147-155.
11 Soares-Weiser K, Fernandez H. Tardive dyskinesia. Semin Neurol 2007; 27: 159–
169.
12 Threadgill D, Miller D, Churchill G, Pardo-Manuel de Villena F. The Collaborative
Cross: a recombinant inbred mouse population for the systems genetics era. ILAR Journal
2001; 52: 24-31.
13 Kerns JM, Sierens DK, Koa LC, Klawans HL, Carvey PM. Synaptic plasticity in the
rat striatum following chronic haloperidol treatment. Clin Neuropharm. 1992;
15(6):488-500.
14 Benes FM, Paskevich PA, Davidson J, Domesick VB. The effects of haloperidol on
synaptic pattern in the rat striatum. Brain Res. 1985; 329:265-174.
15 Meshul CK, Stallbaumer RK, Taylor B, Janowsky A. Haloperidol-induced
morphological changes in striatum are associated with glutamate synapses. Brain Res.
1994; 648(2):181-95.
16 Andreassen O, Meshul CK, Moore C, and Jorgensen HA. Oral dyskinesias and
morphological changes in rat striatum during long-term haloperidol administration.
Psychopharm. 2001; 157: 11-19.
17 Meshul CK, Janowsky A, Casey DE, Stallbaumer RK, Taylor B. Effect of haloperidol
and clozapine on the density of "perforated" synapses in caudate, nucleus accumbens, and
18 Benes FM, Paskevich PA, Davidson J, Domesick VB. Synaptic rearrangements in
medial prefrontal cortex of haloperidol-treated rats. Brain Res. 1985; 348: 15-20.
19 Roberts RC and Lapidus B. Ultrastructural correlates of haloperidol-induced oral
dyskinesias in rats: a study of unlabeled and enkephalin-labeled striatal terminals. J
Neural Transm. 2003; 110: 961–975.
20 National Research Council (US) Committee for the Update of the Guide for the Care
and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals. 8th
edition. Washington (DC): National Academies Press (US); 2011.
21 Fleischmann N, Christ G, Sclafani T, Melman A. The effect of ovariectomy and
long-term estrogen replacement on bladder structure and function in the rat. J Urol 2002; 168:
1265–1268.
22 Hsin-Tung E, Simpson G. Medication-induced movement disorders. In: Kaplan HI,
Sadock BJ (eds). Comprehensive Textbook of Psychiatry. Lippincott Williams and
Wilkins: Philadephia, PA, 2000; 2265–2270.
23 Tomiyama K, McNamara FN, Clifford JJ, Kinsella A, Koshikawa N, Waddington JL.
Topographical assessment and pharmacological char- acterization of orofacial
movements in mice: dopamine D(1)-like vs. D(2)-like receptor regulation. Eur J
Pharmacol 2001; 418: 47–54.
24 Schindelin J, Arganda-Carreras I, Frise E et al. Fiji: an open-source platform for
biological-image analysis. Nature methods 2012; 9(7): 676-682.
25 Meshul CK, Casey DE. Regional, reversible ultrastructural changes in rat brain with
26 Kessas M, Creed M, Nobrega JN. An examination of synaptic proteins following
chronic haloperidol in a rat model of tardive dyskinesia. Psychology & Neuroscience
Supplementary Tables and Figures
!!!!a g e d %7 %w e e k s %
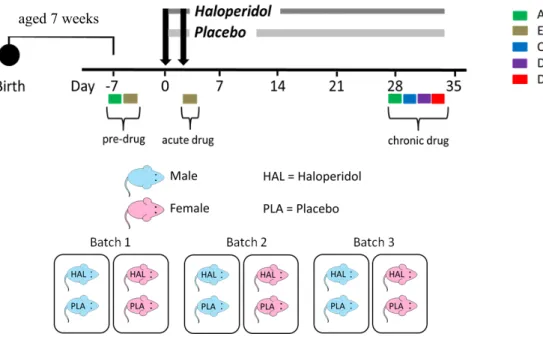
Figure 1. RIX phenotyping pipeline. 8-week old mice (846 male and female mice) from 73 RIX strains were phenotyped in 51 batches using this phenotyping pipeline to identify strain differences in response to haloperidol treatment. Open field activity (Activity) was assessed before and after treatment. Fecal boli will be stored at -80 °C for potential microbiomics work (*). Extrapyramidal symptoms (EPS), or acute side effects, were tested soon after treatment. Vacuous chewing movements (VCMs; i.e. Orofacial) were recorded after 28 days of drug treatment.
Figure 2. Transmission electron microscopy (TEM) was used to assess changes in synaptic density after chronic haloperidol treatment. Representative electron
microscopy image of striatal tissue with a perforated synapse (single arrow) defined by a break between synaptic densities of greater than 0.05 μm, a double synapse (double arrow) where active zones are entering two different synaptic structures and a mitochondria (*). Many mitochondria and synapses are clearly seen in this image.
Figure 7. Differential susceptibility to tremors between RIX strains. Strain-by-strain analysis of the recorded number of tremors VCMs organized by increasing mean of tremors VCMs counts for placebo groups (black squares). Means of counts for
Figure 8. RIX strains do not exhibit differential susceptibility to tongue movements. Strain-by-strain analysis of the recorded number of tongue VCMs organized by
Table 1. Treatment effect on subtle and overt VCMs significantly differs between strains. We fit the data to a Two-Way ANOVA with interaction (after dropping strain 5080x6750 as there only a single mouse of this strain). The null hypothesis that there was no difference in treatment effect between strains was rejected at alpha=0.05 with a calculated p-value of 0.0224 for the combined effect of strain and treatment. Thus, the effect of haloperidol and placebo on combined subtle and overt VCM amount differs between strains.
Source DF SS F-value P-value
Strain 71 1041180 7.3 0.0001
Treatment 1 199349 89.9 0.0001
Strain*Treatment 71 199838 1.4 0.0224
Table 2. Haloperidol-treated and placebo-treated groups within 14 strains are significantly different. Significant differences in the combined subtle and overt VCMs phenotype were found between groups within fourteen strains using a two-way ANOVA and controlling False Discovery Rate at alpha=0.05.
Strain Estimate StdErr tValue Probt False Discovery Rate 8016x8034 112.8 28.3971875 3.97 <.0001 0.0072
559x8031 95.833333 25.922967 3.7 0.0002 0.0086 3140x3015 92.833333 25.922967 3.58 0.0004 0.0089
6513x6188 95.5 28.1424991 3.39 0.0007 0.011
8049x8046 105.333333 31.7490208 3.32 0.001 0.011
5306x5612 105 31.7490208 3.31 0.001 0.011
3252x3154 84 25.922967 3.24 0.0013 0.011
3032x6188 83.333333 25.922967 3.21 0.0014 0.011 8031x3609 83.333333 25.922967 3.21 0.0014 0.011 8008x8016 79.133333 27.1882371 2.91 0.0037 0.027
5156x1566 74 25.922967 2.85 0.0045 0.0292
6188x3252 75.666667 27.1882371 2.78 0.0056 0.0333
8005x8002 77.6 28.3971875 2.73 0.0065 0.0358
Figure 9. Chronic haloperidol treatment does not appear to impact synaptic
Table 3. No significant differences in synaptic architecture and mitochondria amount in 30-day treated C57BL/6 mice. Values after t-test analyses of the sum of synapses, mitochondria, perforated synapses, and double synapses per sample indicate that there is no difference between haloperidol-treated and placebo-treated groups because p-values do not pass the set threshold of 0.0125 after Bonferroni correction.
Observations DF t Ratio P-value (prob > |t|, α = 0.0125)
Sum(synapses) 10 8 0.911597 0.3886
Sum(mitochondria) 10 8 -0.91259 0.3881
Sum(perforated synapses)
10 8 0.452438 0.663