HLA-E–restricted regulatory CD8
T cells are
involved in development and control of human
autoimmune type 1 diabetes
Hong Jiang, … , Zongyu Zheng, Leonard Chess
J Clin Invest.
2010;
120(10)
:3641-3650.
https://doi.org/10.1172/JCI43522
.
A key feature of the immune system is its ability to discriminate self from nonself.
Breakdown in any of the mechanisms that maintain unresponsiveness to self (a state known
as self-tolerance) contributes to the development of autoimmune conditions. Recent studies
in mice show that CD8
+T cells specific for the unconventional MHC class I molecule Qa-1
bound to peptides derived from the signal sequence of Hsp60 (Hsp60sp) contribute to
self/nonself discrimination. However, it is unclear whether they exist in humans and play a
role in human autoimmune diseases. Here we have shown that CD8
+T cells specific for
Hsp60sp bound to HLA-E (the human homolog of Qa-1) exist and play an important role in
maintaining peripheral self-tolerance by discriminating self from nonself in humans.
Furthermore, in the majority of type 1 diabetes (T1D) patients tested, there was a specific
defect in CD8
+T cell recognition of HLA-E/Hsp60sp, which was associated with failure of
self/nonself discrimination. However, the defect in the CD8
+T cells from most of the T1D
patients tested could be corrected in vitro by exposure to autologous immature DCs loaded
with the Hsp60sp peptide. These data suggest that HLA-E–restricted CD8
+T cells may play
an important role in keeping self-reactive T cells in check. Thus, correction of this defect
could be a potentially effective and safe approach in […]
Research Article
Immunology
Find the latest version:
Research article
HLA-E–restricted regulatory CD8
+
T cells
are involved in development and control
of human autoimmune type 1 diabetes
Hong Jiang,1 Steve M. Canfield,1 Mary P. Gallagher,2 Hong H. Jiang,1
Yihua Jiang,1 Zongyu Zheng,1 and Leonard Chess1
1Department of Medicine and 2Department of Pediatrics, Naomi Berrie Diabetes Center (NBDC), Columbia University,
College of Physicians and Surgeons, New York, New York, USA.
A key feature of the immune system is its ability to discriminate self from nonself. Breakdown in any of the
mechanisms that maintain unresponsiveness to self (a state known as self-tolerance) contributes to the
devel-opment of autoimmune conditions. Recent studies in mice show that CD8
+T cells specific for the
unconven-tional MHC class I molecule Qa-1 bound to peptides derived from the signal sequence of Hsp60 (Hsp60sp)
contribute to self/nonself discrimination. However, it is unclear whether they exist in humans and play a role
in human autoimmune diseases. Here we have shown that CD8
+T cells specific for Hsp60sp bound to HLA-E
(the human homolog of Qa-1) exist and play an important role in maintaining peripheral self-tolerance by
dis-criminating self from nonself in humans. Furthermore, in the majority of type 1 diabetes (T1D) patients tested,
there was a specific defect in CD8
+T cell recognition of HLA-E/Hsp60sp, which was associated with failure of
self/nonself discrimination. However, the defect in the CD8
+T cells from most of the T1D patients tested could
be corrected in vitro by exposure to autologous immature DCs loaded with the Hsp60sp peptide. These data
suggest that HLA-E–restricted CD8
+T cells may play an important role in keeping self-reactive T cells in check.
Thus, correction of this defect could be a potentially effective and safe approach in the therapy of T1D.
Introduction
The fundamental question of what is “self” and what is “foreign,” as seen by the immune system, determines how the immune sys- tem discriminates self from nonself. In this regard, the pioneer-ing work of Burnet and Medawar suggested that the definition of self versus nonself is arbitrary because foreign antigens presented during fetal life are thereafter considered self (1). Moreover, it is known that all T cells are self-referential in the sense that they are positively selected for survival on self-peptide(s) bound to MHC molecules during thymic positive selection (2) before thymic nega-tive selection, in which thymocytes expressing TCR of high avidity to self-antigens are deleted (3–5).
We have previously proposed and tested an “avidity model” of peripheral T cell regulation that postulates that, like in the thymus, the immune system discriminates self from nonself during adaptive immunity in the periphery not by recognizing the structural differ-ences between self versus foreign antigens, but rather by perceiving the avidity of T cell activation (6–9). It is generally accepted that thymic negative selection, in which, high-avidity self-reactive thy-mocytes are deleted, eliminates the imminent danger of pathogenic autoimmunity in the periphery and is the major mechanism of cen- tral self-tolerance (3–5). However, while releasing the “innocent” self-reactive T cells with low avidity, thymic negative selection also allows a large fraction of self-reactive T cells of intermediate avidity to be released into the periphery under normal circumstances (10–12), and functional activation of this population of cells has the poten-tial to elicit pathogenic autoimmunity (12–15). The potential to develop autoimmune disease is therefore inherent in every indi-vidual and must be specifically dealt with by peripheral regulatory
mechanisms (6–9). In this regard, we have demonstrated in murine studies that self/nonself discrimination is accomplished by thymic negative selection followed by peripheral T cell regulation in which Qa-1–restricted CD8+
T cells selectively downregulate intermediate-avidity T cells activated by any antigens (6–8). Since the peripheral self-reactive T cell repertoire is devoid of high-avidity self-reactive cells due to thymic negative selection, the selective downregulation of intermediate-avidity T cells simultaneously enables the suppres- sion of autoimmunity and the preservation of the functional anti-infection immunity, which is dominated by high-avidity T cells.
The concept that perceiving the avidity of T cell activation can be translated into peripheral T cell regulation is the essence of the avidity model. The cellular mechanism that defines how perceiv-ing the avidity of T cell activation is translated into peripheral T cell regulation and the molecular structures recognized by regu-latory T cells that enable them to discriminate self from nonself in the periphery are the key issues in regulatory T cell biology. In this regard, we have recently demonstrated that the heat shock peptide Hsp60sp, coupled with the MHC class Ib molecule Qa-1, is a common surrogate target structure preferentially expressed on the intermediate-avidity T cells and is specifically recognized by a subset of Qa-1–restricted CD8+ T cells (7). Thus, by a unified and
simple cognitive mechanism — specific recognition of a common target structure, preferentially expressed on the intermediate-avid-ity T cells — the Qa-1–restricted CD8+ T cells are able to selectively
target and downregulate intermediate- but not high-avidity T cells to accomplish self/nonself discrimination in the periphery (8).
The translation of the murine Qa-1–restricted CD8+
T cell–medi-ated pathway to humans is based on evidence that the human homolog of Qa-1, HLA-E, can function as a restricting element for human regulatory CD8+ T cells (16). Here we show that humans
have a cognitive mechanism similar to that discovered in mice, in
Conflict of interest: The authors have declared that no conflict of interest exists.
Citation for this article: J Clin Invest. 2010;120(10):3641–3650. doi:10.1172/JCI43522.
research article
which HLA-E–restricted CD8+ T cells, the human counterpart of
murine Qa-1–restricted CD8+
T cells, function to maintain self-tol-erance in healthy humans by discriminating self from nonself in the periphery. In addition, we provide evidence that the HLA-E– restricted regulatory CD8+ T cell–mediated pathway plays a role in
the immunopathogenesis of the human autoimmune disease type 1 diabetes (T1D). T1D is an autoimmune disorder in which T cells reactive with major antigenic components of pancreatic β cells have a central role. While these self-reactive T cells are under the control of peripheral regulatory mechanisms in healthy individuals, failure of the control leads to the destruction of the β cells and consequent T1D (17, 18). To directly test this idea, we studied patients with T1D and found that, in the majority of the patients tested, HLA-E– restricted CD8+ T cells have a cognitive defect in their capacity to
recognize the specific common target structure, HLA-E/Hsp60sp, expressed on target cells, and functionally fail to mediate self/non-self discrimination. Moreover, this cognitive and functional defect in the CD8+ T cells in most patients can be corrected by an in vitro
boost of CD8+ T cells with autologous immature DCs loaded with
Hsp60sp peptide. Taken together, these studies provide evidence that the CD8+
T cell recognition of HLA-E/Hsp60sp may be a cen-tral cognitive mechanism enabling self/nonself discrimination in the periphery and that the defect of this pathway may be involved in the development and control of T1D.
Results
Generation of human HLA-E–restricted CD8+ T cell lines that
specifically recognize HLA-E/Hsp60sp expressed on the target cells We have shown previously that murine immature DCs loaded with the Qa-1–binding peptide Hsp60sp, but not Qdm, can be used as a vaccine to induce CD8+ T cell–dependent protection from EAE (7).
This observation suggested that functional HLA-E/Qa-1–restricted CD8+ T cells could be induced by Hsp60sp-loaded immature DCs
in vivo. It is well known that Qdm is the leader sequence derived from conventional MHC class Ia proteins in mice that is capable of
binding to Qa-1 (19). Importantly, the resultant Qa-1/Qdm com-plex is the specific ligand for NKG2A receptor on NK cells, and the recognition of this ligand by NK cells can inhibit NK killing (20–22). Thus, B7sp, the human equivalent of Qdm (23), was used as an ideal control HLA-E–binding peptide for Hsp60sp in our current human studies. Employing this approach, we established a standard protocol and successfully generated two types of human CD8+ T cell lines in vitro from healthy individuals by stimulating
purified peripheral CD8+ T cells with autologous immature DCs
loaded either with Hsp60sp, designated CD8(H) cells, or loaded with HLA-B7sp, designated CD8(B) cells.
The CD8+ T cell lines generated from the healthy donors were
first tested to determine whether they recognize HLA-E/Hsp60sp expressed on the target T cells. We thus established human trans-fectant cell lines that express HLA-E on their surface to identify the HLA-E–binding peptide(s) that could be recognized by the regulatory CD8+ T cells. An HLA-E expression construct was
established and transfected into a human HLA-A/B/C–deficient B cell line, B721 (24). HLA-E expression clones were generated by limiting dilution. The cloned B721/HLA-E transfectants (B721/E) were first tested for their surface expression of HLA-E by staining the peptide-loaded B721/E cells with HLA-E–specific mAb 3D-12 (25). Consistent with other reports (7, 23), both B7sp and Hsp60sp bound to HLA-E and, therefore, stabilized the expression of HLA-E on the cell surface (Supplemental Figure 1; supplemental material available online with this article; doi:10.1172/JCI43522DS1), and thus could be used as HLA-E–binding peptide–presenting cells to test the specific recognition of the target structure by the HLA-E–restricted CD8+ T cells in a CD8+ T cell inhibition assay (7, 8).
Hsp60sp-specific CD8+ T cells inhibit HLA-E–expressing cells
loaded with Hsp60sp but not control peptide B7sp
We then investigated the specificity of the CD8(H) lines generated from the healthy individuals by testing the inhibitory effect of the CD8(H) lines on the HLA-E–expressing transfectant B721/E cells loaded with Hsp60sp, B7sp, or control non–HLA-E–bind-ing peptide (CD8+ T cell inhibition assay). The CD8(H) lines
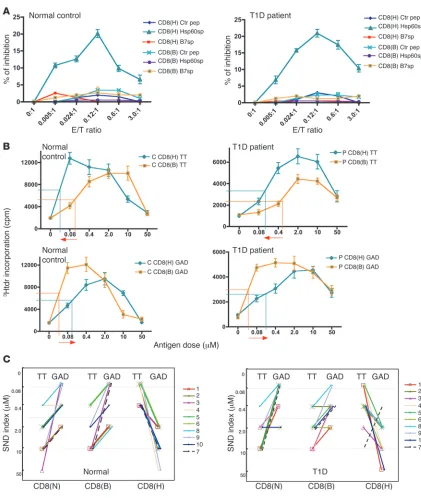
consistently showed in all individuals tested a potent suppres-sion of B721/E cells loaded with Hsp60sp but not B721/E cells loaded with B7sp or control peptide (see representative results in Figure 1A). Moreover, the control CD8(B) lines did not suppress the B721/E targets loaded with any peptides. The specific cogni-tive recognition of HLA-E/Hsp60sp expressed on the target cells by the HLA-E–restricted CD8+ T cells was further confirmed by
the detection of cytolytic enzymes (CEs) secreted by the HLA-E– restricted CD8+ T cells. Thus, increased secretion of CEs, such as
perforin, granzyme A, and granzyme B, by the CD8+ T cells was
observed when the CD8(H) lines were cocultured with B721/E cells loaded with Hsp60sp but not with B721/E cells loaded with control peptide B7sp, or when CD8(B) lines were cocultured with B721/E cells loaded with Hsp60sp or B7sp, as shown by repre-sentative results in Figure 1B. These observations were precisely correlated with the specific killing of targets by the CD8(H) lines that were loaded with Hsp60sp but not B7sp, as detected in the CD8+ T cell inhibition assay above.
We concluded from these experiments that the ability of CD8(H) but not CD8(B) cells to specifically recognize Hsp60sp presented by HLA-E strongly suggested that the CD8(H) and CD(B) cells may represent functionally distinct subsets of CD8+ T cells with
respect to specificity and function.
Figure 1
Generation of human HLA-E–restricted CD8+ T cell lines that function
to discriminate self from nonself by specifically recognizing HLA-E/Hsp-60sp expressed on the target cells. (A) CD8(H) lines from healthy indi-viduals specifically inhibit HLA-E–expressing cells loaded with Hsp60sp. CD8(H) and control lines were generated and tested in a CD8+ T cell
inhibition assay as described in Methods. Data represent mean ± SEM and were from 3 healthy normal individuals. Ctr pep, control peptide. (B) Increased CE expression by the HLA-E–restricted CD8+ T cells
triggered by the specific target structure HLA-E/Hsp60sp. CD8(H) and CD(B) lines from healthy individuals were cocultured with B721/E cells loaded with peptide Hsp60sp or control peptide B7sp. The CE expression by the CD8+ T cells was detected by 3-color intracellular
staining with perforin (Perf), granzyme A (GA), and granzyme B (GB), followed by FACS analysis as described in Methods. Shown are rep-resentative data from 1 of 3 healthy normal individuals. Top row: CE expression indexes of 3 different combinations of CE expression; bot-tom row: intracellular staining patterns of the CEs on the CD8(H) lines. (C) CD8(H) lines from healthy individuals suppress the overall immune responses to self-antigens MBP and GAD but enhance the immune responses to foreign antigens TT and PPD. CD8(H) and control CD8(B) lines were generated and tested in a self/nonself discrimina-tion assay as described in Methods. Data are shown as mean ± SEM and were from 3 healthy normal individuals. 3Hdtr, [3H]thymidine. YRK,
HLA-E–restricted CD8+ T cells function to discriminate
self from nonself in the periphery
We next directly tested the function of self/nonself discrimination by systematically testing the effect of the CD8(H) lines on the over- all avidity of immune responses to self-antigens myelin basic pro-tein (MBP) (or glutamic acid decarboxylase [GAD]) versus those to foreign antigens tetanus toxoid (TT) (or purified protein derivative [PPD]) in a standard T cell proliferation assay as described previ-ously (8). Thus, purified autologous CD4+ T cells were activated by
serial doses of either MBP (and GAD) or TT (and PPD), and CD8(H) cells were added to the CD4+ T cell cultures. CD8(B) and CD8(N)
(primed with DCs without being loaded with peptides) served as controls. In the studies using GAD as a self-antigen, the donors were selected to be DR4+. As shown in Figure 1C, in the same individuals
shown in Figure 1A, the CD8(H) cells suppressed the responses to self-antigens MBP and GAD but enhanced the responses to foreign antigens TT and PPD, compared with control CD8(B) or CD8(N) cells (see data from all controls and T1D patients presented below). The typical pattern of an inhibited immune response was shown by a decreased overall avidity, reflected by an increased median effective dose (ED50), in immune responses to self-antigen MBP and GAD in
the presence of CD8(H) compared with the responses to the same antigens in the presence of control CD8(B) and CD8(N) cells. In contrast, the typical pattern of an enhanced immune response was shown by an increased overall avidity, reflected by a decreased ED50,
[image:5.585.46.542.80.454.2]in immune responses to foreign antigen TT and PPD in the pres-ence of CD8(H) compared with the responses to the same antigens in the presence of control CD8(B) and CD8(N) cells.
Figure 2
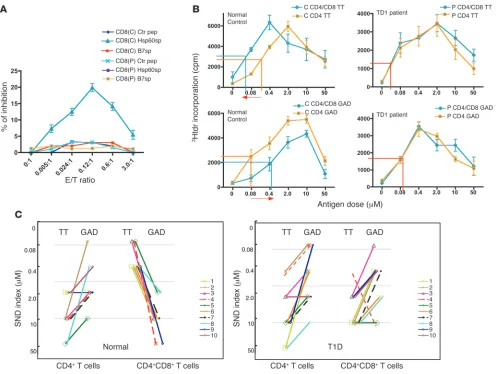
A defect in the HLA-E–restricted CD8+ T cell–mediated pathway is detected in a majority of the T1D patients tested. (A) Freshly isolated CD8+ T
cells from a T1D patient lost the capacity to specifically recognize HLA-E/Hsp60sp target structure. CD8+ T cells were purified from T1D patients
and tested in a CD8+ T cell inhibition assay as described in Methods. Data show mean ± SEM and are representative of 9 of 10 T1D patients
tested (P) paired with healthy normal controls (C). (B) Freshly isolated CD8+ T cells from a T1D patient lost the capacity to discriminate self from
nonself. CD8+ and CD4+ T cells were purified from T1D patients, and responses of CD4+ T cells were tested and compared in the presence
and absence of CD8+ T cells in a standard self/nonself discrimination assay as described in Methods. Data are shown as mean ± SEM and are
representative of 9 of 10 T1D patients tested paired with healthy normal controls. (C) Freshly isolated CD8+ T cells from the majority of the T1D
patients tested lost the capacity to discriminate self from nonself in the periphery. Immune responses of purified CD4+ T cells to self-antigen GAD
versus to foreign antigen TT were compared with those of CD4+ T cells plus CD8+ T cells in each T1D patient, paired with normal control. Data
research article
Taken together, the results suggest that combination of the two functional assays provides a unique and reliable assay system that enables the precise identification and detection of human HLA-E– restricted CD8+
T cell populations. Moreover, it links the specific-ity of HLA-E–restricted CD8+ T cells to their biological function of
self/nonself discrimination. These cognitive and functional prop-erties together make a clear distinction between the unique CD8+
T cells that recognize Hsp60sp bound by HLA-E [CD8(H)] and the CD8+ T cells recognizing HLA-E binding to other peptides, such
as the HLA-E/B7sp complex [CD8(B)]. This observation indicates that a major consequence of regulation by the HLA-E–restricted, Hsp60sp-specific CD8+
T cells is its differential effect on the over- all immune response to self versus to foreign antigens. These stud-ies demonstrate that HLA-E–restricted, Hsp60sp-specific CD8+ T
cells, which are capable of discriminating self from nonself, exist and function in healthy human subjects.
Do HLA-E–restricted CD8+ T cells that function
to discriminate self from nonself play a role in the development and control of human T1D?
To determine whether HLA-E–restricted CD8+ T cells also play a
role in the control of human T1D, we tested the function of CD8+
T cells from patients at the NBDC with recent-onset T1Ds. We demonstrate below that via the same mechanism identified in mice, HLA-E–restricted CD8+ T cells exist and play an important
role in maintaining self-tolerance in healthy humans by discrimi-nating self from nonself in the periphery and that this functional property was defective in a majority of the T1D patients tested.
A defect in the HLA-E–restricted CD8+ T cell–mediated pathway
is detected in a majority of the T1D patients tested
We previously demonstrated that Qa-1–restricted CD8+ T cells control
the spontaneous development of T1D in NOD mice, in part, by means of self/nonself discrimination (8). To determine whether HLA-E– restricted CD8+ T cells also play a role in the control of human
T1D, we tested the function of freshly isolated CD8+ T cells from
10 patients with recent onset of T1D. The standard antigens cho-sen were TT for foreign antigen and GAD and MBP for self-antigens,
and each patient tested was selected as DR4+ and
paired with a DR4+ healthy normal control. CD8+ T cells freshly isolated from most of the T1D patients tested fail to recognize specific target structure HLA-E/Hsp60sp expressed on target cells. We first identified the specificity of the CD8+ T cells as
described above. Representative results presented in Figure 2A show that while CD8+ T cells from a
normal control specifically inhibited the B721/E cells loaded with Hsp60sp, but not control pep-tides, CD8+ T cells from a T1D patient failed to
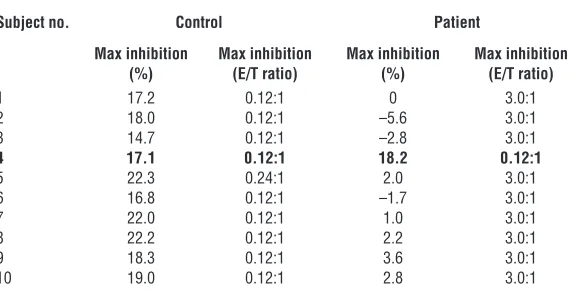
inhibit B721/E cells loaded with Hsp60sp. More-over, as summarized in Table 1, freshly isolated CD8+ T cells from 9 of 10 T1D patients tested
failed to specifically inhibit B721/E cells loaded with Hsp60sp, compared with normal controls (P < 0.001), precisely correlating with the failure of self/nonself discrimination in each patient as summarized in Figure 2C (see below).
CD8+ T cells freshly isolated from most of the T1D patients tested have a defect in the capacity to discrimi-nate self from nonself . We then compared the over-all immune responses of purified CD4+ T cells to self versus foreign
antigens in the presence or absence of freshly isolated CD8+ T cells.
Representative results in 9 of 10 T1D patients tested compared with normal controls are shown in Figure 2B. Thus, the CD8+ T
cells from normal controls significantly depressed the overall CD4+
T cell response to self-antigen GAD, reflected by an increased ED50,
while enhancing the overall response to foreign antigen TT, reflect-ed by a decreased ED50, compared with CD4+
T cell cultures with-out CD8+ T cells. However, when the same tests were performed
in the T1D patients, there was no significant difference in ED50
between purified CD4+ T cells with and those without CD8+ T cells
responding to either self-antigen GAD or to foreign antigen TT, indicating that CD8+ T cells from T1D patients have a defect in or
have lost the capacity to discriminate self from nonself.
In addition to ED50, a new parameter was designed to evaluate
the data from the self/nonself discrimination assay: the self/nonself discrimination index (SND index). The SND index represents the antigen dose that elicits the highest T cell proliferation of immune response to a particular antigen and can be used to directly reveal the relationship of T cell responsiveness to self versus to foreign antigens in each individual. This can be graphically shown by a line connecting SND index of TT versus GAD or MBP in each test. The direction of the connecting line in each individual indicates the relationship between immune responses elicited by self versus by foreign antigens. A high SND reflects a reduced antigen-specific responsiveness, and conversely, falling SND indicates increasing antigen responsiveness. When data from more than 200 tests per-formed in both normal controls and T1D patients were analyzed side-by-side using SND index and ED50 as parameters to evaluate
the responsiveness of T cells, SND indexes were always consistent with ED50 in each test. Thus, SND index was used as an additional
simple and straightforward parameter to present the summarized data from self/nonself discrimination assay throughout this study.
Results from 10 T1D patients with corresponding controls are summarized in Figure 2C and Supplemental Table 1. Notably, in the absence of CD8+ T cells, the responses to self-antigen GAD
[image:6.585.56.343.123.272.2]were higher than to foreign antigen TT, reflected by lower SND indexes for GAD than for TT, in both T1D and normal controls.
Table 1
Comparison of the specificity of freshly isolated CD8+ T cells between T1D patients and normal controls
Subject no. Control Patient
Max inhibition Max inhibition Max inhibition Max inhibition
(%) (E/T ratio) (%) (E/T ratio)
1 17.2 0.12:1 0 3.0:1 2 18.0 0.12:1 –5.6 3.0:1 3 14.7 0.12:1 –2.8 3.0:1
4 17.1 0.12:1 18.2 0.12:1
5 22.3 0.24:1 2.0 3.0:1 6 16.8 0.12:1 –1.7 3.0:1 7 22.0 0.12:1 1.0 3.0:1 8 22.2 0.12:1 2.2 3.0:1 9 18.3 0.12:1 3.6 3.0:1 10 19.0 0.12:1 2.8 3.0:1
Freshly isolated CD8+ T cells from the majority of the T1D patients tested fail to recognize
spe-cific target structure HLA-E/Hsp60sp expressed on target cells compared with normal controls.
In the presence of CD8+ T cells, the normal controls exhibited the
reverse phenomenon: the GAD responses were inhibited (reflect-ed by increased SND indexes) among all individuals tested, while responsiveness to foreign antigen (TT) was increased. In contrast, 9 of 10 T1D patients tested showed the same distribution pattern of SND indexes, regardless of the presence or absence of CD8+ T
cells, suggesting that the presence of CD8+ T cells in the culture
had no significant impact on antigen-driven proliferation in those T1D patients. This observation was further confirmed in cultures examining reactivity to another (T1D-unrelated) self-antigen, MBP (Supplemental Figure 2 and Supplemental Table 2).
The CD8+ T cells from most T1D patients tested could be
boosted in vitro to restore their function
We further tested whether the specific cognitive defect of CD8+ T
cells from T1D patients could be corrected by boosting the CD8+
T cells. Thus, the CD8+ T cells from the T1D patients were boosted
in vitro by priming with autologous immature DCs loaded with Hsp60sp [CD8(H)] or B7sp [CD8(B)] or not loaded with peptide [CD8(N)], as described above.
The function of the boosted CD8+ T lines was first tested for
their specificity in a CD8+ T cell inhibition assay. As shown in
Figure 3A, CD8(H) cells from a T1D patient specifically inhib-ited B721/E cells loaded with Hsp60sp, but not B7sp or control peptide, as effectively as those form a healthy individual. CD8(B) cells from the same patient did not inhibit at all. As summarized in Table 2, this phenomenon was also observed in 8 of 9 T1D patients originally found to have a defect in their CD8+ T cells. However,
the function of the CD8+ T cells in 1 of 9 T1D patients (no. 7) was
not restored by just a simple boost in vitro. After restoration of the function of the CD8+ T cells in the 8 subjects who responded,
statistical analysis showed no significant difference between the CD8+ T cells of the T1D group and a normal control group with
regard to their ability to specifically inhibit HLA-E–expressing tar-gets loaded with Hsp60sp (P = 0.98 without T1D subject no. 7).
Representative results on restored function of self/nonself dis-crimination are shown in Figure 3B. CD8(H) lines, generated from the same 8 of 9 T1D patients by an in vitro boost with the autologous
DCs loaded with Hsp60sp, regained the function of self/nonself discrimination compared with the normal controls. As summarized in Figure 3C and Supplemental Table 3, in the presence of the control CD8(N) and CD8(B) lines, the SND indexes were significantly lower in CD4+ T cells
responding to GAD than to TT (P < 0.05) in both T1D and normal control groups, reflecting an uncontrolled reactivity to self-antigen GAD. However, when CD8(H) cells were added into the CD4+ T cell cultures, in 9 of 10 patients (except
patient no. 7) and 10 normal controls, the SND indexes became significantly higher in GAD responses than in TT responses (P < 0.05). Thus, under the control of the CD8(H) lines, an inhib- ited immune response to self-antigen GAD asso- ciated with an increased immune response to for-eign antigen TT was detected in most of the T1D patients compared with the normal controls.
Thus, CD8+ T cells from most T1D patients
tested could regain their specific cognitive function, at a molecular/cellular level, by an in vitro boost with autologous immature DCs loaded with Hsp60sp, leading to a restoration of their ability to discriminate self from nonself at a biological system level.
Discussion
Our current studies provide evidence that via the same cognitive mechanism that we have identified in mice, the HLA-E–restricted CD8+ T cells that function to discriminate self from nonself do
exist and operate in humans. Furthermore, we found that the HLA-E–restricted CD8+ T cells have a specific cognitive defect in a
large fraction of T1D patients, suggesting that this defect may be involved in the development and control of T1D.
The specificity of HLA-E–restricted CD8+ T cells determines their biological function of self/nonself discrimination: the molecular/cellular mechanism of self/nonself discrimination. In a 1990 editorial in Science , Daniel E. Kosh- land Jr. wrote, “Of all the mysteries of modern science, the mecha-nism of self versus nonself recognition in the immune system ranks at or near the top” (26). Clearly, self-tolerance must be achieved by mechanisms of self/nonself discrimination. However, in the field of immune regulation, this concept seems to be ignored when the peripheral mechanisms of self-tolerance are studied. Despite the numerous research articles written during the last several decades on the peripheral regulation of the immune responses in both human autoimmune disease and murine models of autoimmunity, few, if any, directly dealt with the cognitive mechanisms of peripheral self/nonself discrimination (9). In this regard, discrimination of self from nonself and control of the magnitude and class of the immune response are two equally important but distinct peripheral regula-tory mechanisms that operate in concert to ensure optimal function of the immune system (9, 27, 28). In general, while mechanisms of self/nonself discrimination evolve to establish and maintain self-tol-erance, control of the magnitude and class of the immune response serves to ensure an optimal response to foreign pathogens by avoiding collateral damage from excessive reactions or an improper response due to an inadequate class of immune response (9, 27, 28). However, currently, almost all the identified peripheral regulatory mechanisms including those employed by the various types of regulatory CD4+
T cells function by controlling the magnitude and class of immune
Table 2
Comparison of the specificity of CD8+ T cell lines between T1D patients and normal controls
Subject no. Control Patient
Max inhibition Max inhibition Max inhibition Max inhibition
(%) (E/T ratio) (%) (E/T ratio)
1 17.0 0.12:1 14.8 0.12:1 2 22.6 0.12:1 23.6 0.12:1 3 17.9 0.12:1 19.4 0.12:1 4 17.1 0.12:1 18.2 0.12:1 5 17.1 0.24:1 18.7 0.12:1 6 16.9 0.12:1 17.8 0.12:1
7 19.8 0.12:1 2.1 3.0:1
8 18.4 0.12:1 20.2 0.12:1 9 28.7 0.12:1 26.2 0.24:1 10 21.8 0.24:1 18.9 0.12:1
CD8+ T cells from most T1D patients tested regain the capacity to recognize specific target
research article
Figure 3
The CD8+ T cells from most T1D patients tested could be boosted in vitro to restore their function. (A) CD8+ T cells from a T1D patient restored
the capacity to specifically recognize the target structure HLA-E/Hsp60sp, after being boosted in vitro with autologous DCs loaded with Hsp60sp peptide. CD8(H) and control CD8+ lines were generated from each T1D patient and corresponding normal control and were tested and compared
in a CD8+ T cell inhibition assay. Data are shown as mean ± SEM and are representative of 8 of 9 T1D patients who originally tested with a defect
of the HLA-E–restricted CD8+ T cells. (B) CD8+ T cells from a T1D patient restored the capacity to discriminate self from nonself after an in vitro
boost with autologous DCs loaded with Hsp60sp peptide. CD8(H) and control CD8(B) lines were generated from each T1D patient and corre-sponding normal control and tested and compared in a self/nonself discrimination assay. Data are shown as mean ± SEM and are representative of 8 of 9 T1D patients who originally tested with defect of the HLA-E–restricted CD8+ T cells. (C) CD8+ T cells from most T1D patients tested
responses (27, 28). While this is important, it does not provide a bio- logical basis for the specificity required to precisely, safely, and effec-tively prevent and treat human organ-specific autoimmune diseases caused by failure of peripheral self/nonself discrimination (9, 16, 28, 29). In this regard, we have demonstrated in mice that one of the cen-tral characteristics of the HLA-E/Qa-1–restricted CD8+
T cell–medi-ated pathway is its unique specificity of regulation. The specificity of the regulation is not at the level of antigens that activate the target T cells but at the level of the particular common surrogate target structure, such as Qa-1/Hsp60sp, preferentially expressed on the target T cells as a function of intermediate-avidity activation (6, 7), which could be initiated by any antigens (8). This feature of the HLA-E/Qa-1–restricted CD8+ T cells enables the immune system to
employ a unified and simple cognitive molecular/cellular mecha-nism, at a biological system level, to discriminate self from nonself, in order to maintain self-tolerance. The observation in our current studies that defects in this cognitive recognition of HLA-E/Hsp60sp by the HLA-E–restricted CD8+ T cells were associated with failure of
self/nonself discrimination in the majority T1D patients tested pro-vides evidence from human disease that supports this notion. This is also supported by independent studies in Qa-1–KO mice (30) and, more precisely, in Qa-1 mutant knock-in mice possessing impaired Qa-1 binding to the TCR/CD8 coreceptor (31), providing direct and unequivocal molecular evidence that the cognitive recognition of Qa-1/peptide on the target T cells by the Qa-1–restricted CD8+ T cells
is crucial in maintaining self-tolerance.
The significance of HLA-E–restricted CD8+
T cells in maintain- ing peripheral self-tolerance has been demonstrated by self/non-self discrimination assays in our current studies. In all the tests performed with different settings in normal controls, the immune responses of CD4+ T cells to self-antigens were always significantly
higher than (P < 0.05) or the same as immune responses to for-eign antigens, reflected by the lower or similar SND indexes, in the absence of the HLA-E–restricted CD8+ T cells. In contrast, in
all the tests containing the HLA-E–restricted CD8+ T cells, the
immune responses of CD4+ T cells to self-antigens were always
significantly lower (P < 0.05), reflected by the higher SND indexes, compared with immune responses to foreign antigens, without any exceptions (Supplemental Tables 1–3, Figure 2C, Figure 3C, and Supplemental Figure 2). This notion is further supported by the fact that the majority of the T1D patients tested have lost the function of their HLA-E–restricted CD8+ T cells.
Given the observations in mice that intermediate-avidity T cells preferentially express a common target structure on their surface (7–9), we have established two assays based on the central biology of this regulatory pathway. These two assays, together, transformed the basic science into a direct and practical approach in humans to identify HLA-E–restricted CD8+ T cells. Thus, detection of the
specificity of the HLA-E–restricted CD8+ T cells recognizing HLAE/
Hsp60sp preferentially expressed on intermediate-avidity target T cells can be directly linked to the biological function of self/non-self discrimination at a system level without the need for indirectly identifying the intermediate-avidity target T cells at a clonal level. As importantly, self/nonself discrimination can be experimentally detected only at a system level but not at a clonal level, to reveal its biological and functional significance. Taken together, the results suggest that definitive specificity of the HLA-E–restricted CD8+ T
cells in combination with their biological function of self/nonself discrimination have led to the conclusion that the molecular/cel-lular mechanism of self/nonself discrimination in our system is
mediated by the HLA-E–restricted CD8+ T cells that selectively
downregulate the target T cells expressing HLA-E/Hsp60sp. These two assays together provide a unique and reliable assay system enabling the identification of the specificity (7) and the determination of the biological function of self/nonself discrimi-nation (8) of human HLA-E–restricted CD8+ T cells in both basic
and clinical studies. The data obtained by these assays are statis-tically significant (Tables 1 and 2 and Supplemental Tables 1–3) and consistent with the murine observations that the degree and magnitude of the regulation detected are sufficient to effectively protect animals from T1D and EAE (7, 8).
It is of interest that Qa-1/HLA-E is a central molecule that links the NK pathway to the T cell regulation. We demonstrated in our cur-rent studies that via a specific cognitive recognition of the HLA-E/ Hsp60sp expressed on the target T cells, the HLA-E–restricted CD8+
T cells discriminate self from nonself in the periphery. It could be postulated that the target T cells expressing higher levels of HLA-E/ Hsp60sp would express less HLA-E/B7sp on their surface and, there- fore, be more susceptible to NK killing as well. This may link the reg-ulation of adaptive immunity to innate immunity in the context of self/nonself discrimination, which should be further explored (9).
The in vivo relevance of the HLA-E–restricted CD8+ T cell–mediated path-way suggested by the current studies. The relevance of the assay system to in vivo activities of the HLA-E–restricted CD8+ T cell population was
shown by two major sets of results. First, in samples from healthy individuals, there were no functional differences between freshly iso-lated CD8+ T cells (Figure 2 and Table 1) and in vitro boosted HLA-E–
restricted CD8+ T cells (Figure 3 and Table 2), as revealed by the two
assays. These data indicated that HLA-E–restricted CD8+ T cells are
constantly activated and function in vivo in healthy people. The in vivo relevance of in vitro generation of CD8(H) and CD8(B) lines is also supported by the observation that vaccinating animals with Hsp60sp-loaded, but not Qdm-loaded, DCs can induce a CD8+ T
cell–dependent protection from EAE (7). Second, Figure 2, A–C, pres-ents the data comparing the function of freshly isolated CD8+ T cells
from blood of T1D patients and normal controls, revealed by the specificity and self/nonself discrimination assays. These cells were not manipulated in vitro. The cognitive defects of the HLA-E–restricted CD8+ T cells in most of T1D patients tested, compared with normal
individuals, represent the direct relevance of this regulatory pathway in vivo. In addition, since the in vivo relevance of the in vitro gener-ated CD8+ T cell lines have been demonstrated in normal controls,
these cellular reagents could be used for providing extremely useful information if the defect of the CD8+ T cells from T1D patients can
be corrected by an in vitro boost with autologous DCs loaded with Hsp60sp in clinical settings for potential prognosis.
In the current studies, among the 10 T1D patients tested, 9 revealed evidence of a specific cognitive defect of HLA-E–restricted CD8+ T
cells, with one exception, patient no. 4. The high incidence of the defect of the HLA-E–restricted CD8+
T cells in the T1D patients test-ed makes it possible to postulate that HLA-E–restricted CD8+ T cell–
mediated pathway may play a major role in the development and con-trol of human T1D. However, CD8+ T cells from patient no. 7 could
not be corrected. This observation indicates that the specific cogni-tive defect of the CD8+ T cells in this patient may not be at the level
of the CD8+ T cell itself, but at the level of either DCs, which may not
be capable of inducing the CD8+ T cells, or CD4+ T cells, which may
not be susceptible to the CD8+
research article
elevated ability to respond to self-antigen independent of HLA-E– restricted CD8+ T cells. The ability to identify the subset of T1D
patients with a defect of HLA-E–restricted CD8+ T cells will enable
future natural history studies in T1D to determine whether the func-tion of these regulatory cells correlates with clinical course.
Identification of the peripheral regulatory mechanisms that func-tion to discriminate self from nonself would enable novel clinical interventions to prevent and treat autoimmune diseases without damaging anti-infection and antitumor immunity, which is a major side effect of the immunotherapeutic trials currently under inves- tigation for the treatment of T1D. Our findings suggest a poten-tial treatment of T1D via vaccination of patients with autologous immature DCs loaded with Hsp60sp to specifically activate the HLA-E–restricted CD8+ T cells in vivo. This potential therapeutic
approach is supported by the fact that vaccinating B10PL mice with immature DCs loaded with Hsp60sp, but not control peptide Qdm, effectively protected animals from subsequently induced EAE and that the protection was CD8+ T cell dependent (7). If proven correct
in vivo in humans, this approach would specifically prevent continu-ous destruction of “regenerated” β cells that may arise spontaneous-ly or by any possible advanced modern therapeutic technology, such as stem cell transplantation, in established T1D patients. Further-more, the unique assay system may improve our ability to accurately predict the development of T1D and may allow earlier diagnosis of patients. β Cell mass decreases during the asymptomatic prodromal period of pre-T1D (32, 33), so diagnosis and intervention at the ear-liest stages could potentially provide an opportunity to intervene before any significant destruction of β cells occurs.
Methods
Reagents and human samples. Anti–HLA-E mAb 3D-12 and control mAb 4D-12 were a gift from Daniel Geraghty (Fred Hutchinson Cancer Research Center, Seattle, Washington, USA). The staining reagents — fluorescein-conjugated (Fl-conjugated) anti–human CD8, phycoerythrin-conjugated (PE-conjugated) anti–human CD4, Fl-conjugated goat anti-mouse, PE–anti–human perforin, Fl–anti–human granzyme A, and Cy-Streptavidin — were purchased from BD. Biotin-conjugated (Bio-conjugated) anti–human granzyme B was purchased from R&D Systems. Peptides — hHSP60sp (QMRPVSRVL); hB7sp (VMAPRT- VLL); TTp830-843 (QYIKANSKFIGITE); hGAD65p555-567 (NFFRMVISN-PAAT); hMBPp84-102 (NPVVHFFKNIVTPRTPPP) — were synthesized by GeneScript Corp. PPD was purchased from Sanofi Pasteur Ltd.
Blood samples from T1D patients and controls were obtained under a protocol approved by the institutional review board of Columbia Univer-sity College of Physicians and Surgeons. All subjects (or their guardians) provided written informed consent.
Preparation of human PBMCs and purified CD8+ and CD4+ T cells. PBMCs were prepared from heparinized blood, which was diluted 1:1 with HBSS and lay-ered over Ficoll-Hypaque, and centrifuged at 1,000 g for 30 minutes in a Sor-vall RC-3 centrifuge (Ivan Sorvall Inc.). The lymphocytes at the serum-Ficoll interface were removed and washed 3 times in preparation for further puri-fication of subsets. Both CD8+ and CD4+ T cells in all experiments presented
in this study were positively selected by MACS magnetic beads (Miltenyi Biotec) as described previously (6). Briefly, the PBMCs were incubated with anti–human CD4- or CD8–conjugated magnetic beads at 10 × 106 cells/10 μl
of beads, and the CD+ and CD– populations were isolated using a separation
column exposed to a magnetic field according to the manufacturer’s proto-col. The purity of the CD4+ or CD8+ T cells was greater than 95%.
Generation of HLA-E expression transfectants. HLA-E fusion construct (pDsRed) was engineered by RT-PCR from the human B cell line B721 using the following primers: CCAAGCTTATGGTAGATGGAACCCTCCTTT
(forward) and GGGGATCCAACAAGCTGTGAGACTCAGACCC (reverse). Amplified clones in pCR2.1 were fully sequenced. Six independent full-length clones representing the HLA-E 101 haplotype but lacking the 3′ termination codon were subcloned into the mammalian expression vector pDsRed-Express-N1 (Clontech Laboratories Inc.), yielding a single open reading frame encoding a fusion protein consisting of HLA-E joined to a variant of the Discosoma species red fluorescent protein (Clontech Labora-tories Inc.). The pDsRed–HLA-E construct was introduced into the HLA class I–deficient B cell line B721 (24) by electroporation, and stable clones were selected by subcloning in Geneticin (G418).
Testing HLA-E surface expression of HLA-E transfectants . We assessed the sur- face expression of HLA-E on B721 cells transfected with HLA-E by exog-enously loading the cells with hHsp60sp and B7sp or control non–HLA-E– binding peptides at 26°C for 18 hours. Cells were then washed, stained with anti–HLA-E mAb 3D-12 (25) followed by Fl–goat anti-mouse Ig, and analyzed on a FACScan flow cytometer and by CellQuest software (BD) as previously described (6). mAb 4D-12 served as control (25).
Generation of HLA-E–restricted Hsp60sp-specific CD8+ T cell lines. DCs were derived from PBMCs depleted of CD4+ and CD8+ T cells and were cultured in
6-well plates in serum-free Click’s medium at 37°C, 5% CO2, for 1–1.5 hours.
The wells were gently washed. The nonadherent cells were washed away, and the adherent cells were then cultured in medium containing GM-CSF and IL-4 at final concentrations of 80 ng/ml and 20 ng/ml, respectively. DCs were harvested on D6 and loaded with Hsp60sp, B7sp, or no peptide at 50 μM, 37°C, for 2 hours. The purified CD8+ T cells (1.5 × 106 to 2 × 106)
were then cocultured with 0.5 × 106 peptide-loaded DCs in 1 ml in a 48-well
plate to set up the lines of CD8(H), CD8(B), and CD8(N) cells. IL-2 was added on the second day. These 3 types of lines were established for each T1D patient and control through out this study.
CD8+ T cell inhibition assay. Freshly isolated CD8+ T cells were purified from
PBMCs, and CD8(H) and CD8(B) lines were generated as described. HLA-E– transfected cells (B721/E) served as targets and were passively loaded with testing peptides — Hsp60sp, B7sp, and control non–HLA-E–binding peptide — overnight at 26°C. Equal numbers of unlabeled B721/E cells loaded with peptides and CFSE-labeled parental B721 cells that were not loaded with peptide were mixed, and testing CD8+ T cells were added to the targets at
graded E/T ratios, from 3:1 to 0.005:1. We studied the specificity of freshly isolated CD8+ T cells by comparing their inhibition of target B721/E cells
loaded with Hsp60sp versus those loaded with B7sp. In addition, the speci-ficity of CD8(H) lines was compared with that of control CD8(B) lines. In this regard, we established that CD8(H) cells tested had no effect on B721 cells alone or B721 cells pulsed with Hsp60sp or B7sp. On day 5–6, the cell mixtures were assessed by FACS analysis, in which the CD8+ T cells were gated
out during the analysis. The ratio between the two types of targets was calcu-lated and compared in the presence or absence of the CD8+ T cells to evaluate
the effect of testing CD8+
T cells on the targets. The percent specific inhibi-tion was calculated as: {[ratio of loaded B721/E versus unloaded B721 cells in control cultures (without CD8+ T cells) – ratio in experimental cultures (with
CD8+ T cells)]/ratio in control cultures} × 100 (7, 8). Statistical analysis by
2-tailed Student’s t test of the highest percentage of the inhibition was used to evaluate significant differences among different groups (P < 0.05).
Detection of intracellular CEs secreted by the CD8+ T cells. CD8(H) and CD8(B) lines were generated from healthy individuals as described. The established HLA-E–transfected cells (B721/E) served as targets to trigger the CD8+ T