Copyright2001 by the Genetics Society of America
Complex Organization of Promoter and Enhancer Elements Regulate
the Tissue- and Developmental Stage-Specific Expression of the
Drosophila melanogaster Gld
Gene
Brian L. Keplinger, Xinmei Guo, Jonathan Quine, Yue Feng and Douglas R. Cavener
Department of Molecular Biology, Vanderbilt University, Nashville, Tennessee 37205 Manuscript received May 23, 2000
Accepted for publication November 2, 2000
ABSTRACT
TheDrosophila melanogaster Gldgene has multiple and diverse developmental and physiological functions. We report herein that interactions among proximal promoter elements and a cluster of intronically located enhancers and silencers specify the complex regulation ofGld that underlies its diverse functions.Gld expression in nonreproductive tissues is largely determined by proximal promoter elements with the exception of the embryonic labium whereGldis activated by an enhancer within the first intron. A nuclear protein, GPAL, has been identified that binds the Gpal elements in the proximal promoter region. Regulation ofGldin the reproductive organs is particularly complex, involving interactions among the Gpal proximal promoter elements, a unique TATA box, three distinct enhancer types, and one or more silencer elements. The three somatic reproductive organ enhancers each activate expression in male and female pairs of reproductive organs. One of these pairs, the male ejaculatory duct and female oviduct, are known to be developmentally homologous. We report evidence that the other two pairs of organs are developmentally homologous as well. A comprehensive model to explain the full developmental regulation ofGldand its evolution is presented.
G
LUCOSE dehydrogenase (GLD), a rare flavo-sele- regulatory pathways: sex determination (Feng et al. noenzyme that generates reactive oxygen species, 1991) and ecdysone, an insect steroid hormone (Mur-participates in an eclectic set of seemingly unrelated thaand Cavener1989). Temporal expression ofGld functions in Drosophila (Cavener and MacIntyre mRNA during embryogenesis, larval development, and 1983; Cavener 1992; Cox-Foster and Stehr 1994). metamorphosis is controlled by ecdysone via the Gld DuringDrosophila melanogasterdevelopment GLD is ex- proximal promoter region. The sex-determination path-pressed in specific sensory organs including the anten- way controls the tissue-specific expression ofGldmRNA no-maxillary complex as well as several ectodermally in the somatic reproductive organs of the adult by re-derived tissues including the somatic reproductive or- pressingGldexpression in specific reproductive organs. gans (Cox-Fosteret al.1990). GLD is critically required The expression of GLD is particularly intriguing in the to modify the puparial operculum so that the newly reproductive organs. InD. melanogaster, as well as most developed adult fly can escape from its puparium case other Drosophila species, Gldis expressed in a specific (Cavener and MacIntyre1983). At the adult stage, subset of the male and female somatic reproductive GLD participates in two unrelated functions: female organs during their development including the male fertility (our unpublished data) and immunity (Cox- ejaculatory duct and ejaculatory bulb and the female
Foster and Stehr 1994; Lovallo and Cox-Foster oviduct, seminal receptacle, spermathecae, parovaria,
1999). A singleGldgene (Caveneret al.1986) is respon- and vaginal plate (Schiffet al.1992;Rosset al.1994). sible for all of these diverse expression patterns and Shortly after eclosionGldis expressed in a more limited functions; consequently the transcriptional regulation subset of the mature adult somatic reproductive organs. ofGldis exceedingly complex. Various Drosophila spe- In contrast to the conservation of Gld expression pat-cies and other insects appear to share these patterns terns during development, the patterns of expression of expression during preadult development but display at the adult stage vary dramatically among the 54 species different patterns in the adult somatic reproductive or- characterized in the genus Drosophila (Schiff et al. gans (Krasney et al. 1990; Schiff et al.1992; Ross et 1992). Among these species, five distinct male expres-al.1994). sion pattern types and six distinct female expression Gld transcription is regulated in part by two major pattern types are observed in the adult reproductive tract. Although GLD is abundantly expressed in the male ejaculatory duct ofD. melanogasterand a few other
Corresponding author:Douglas R. Cavener, Department of Biology,
closely related species, it is not expressed in any
repro-208 Mueller Laboratory, The Pennsylvania State University, University
Park, PA 16802. E-mail: [email protected] ductive organ in adult males in ⵑ20% of the other
The ⫺425dGpal⫺Gld promoter was PCR amplified from species. Expression of GLD in the adult female
sperma-genomic DNA isolated from pWZ170 transgenic flies, which thecae is highly conserved among species but expression
carry the⫺425dGpal⫺/LacZtransgene. The⫺425dGpal⫺PCR in other female organs (e.g., seminal receptacle, parova- product was digested withEcoRI andKpnI and inserted into ria, and oviduct) is highly species specific. the unique EcoRI/KpnI sites in SREC/LacZ, creating SREC/
⫺425dGpal⫺/LacZ. Previously we showed that a somatic reproductive
or-To construct theSREC/dGpal/hsp70/LacZ transgene, the gan enhancer complex (SREC) activates the Gld
pro-3.3-kbXbaI-EcoRI SREC fragment fromGldintron I was isolated moter fused to alacZreporter gene in the developing
as anEcoRIfragment subsequently blunt ended with T4 DNA and mature reproductive tracts similar to the pattern polymerase. It was then inserted in the reverse orientation observed for the normal Gld gene in D. melanogaster into the unique blunt-endedKpnI site ofdGpal/hsp70/LacZ (Gunaratneet al.1994) to createSREC/dGpal/hsp70/LacZ. (Keplingeret al.1996). However, in combination with
To construct theSREC/⫺425mTATA/LacZ transgene, the 3.3-a heterologous promoter, SREC 3.3-activ3.3-ates expression in
kbXbaI/EcoRI SREC fragment fromGldintron I was isolat-a much smisolat-aller subset of the reproductive orgisolat-ans,
sug-ed as anEcoRI fragment from pEG25 and inserted in the re-gesting that theGldpromoter in combination with SREC verse orientation into⫺425mTATA/LacZ(pWAG183).⫺425m
plays an important role in reproductive tract expression. TATA/LacZ contains the TATA mutant PstI/BclI ⫺425 Gld promoter fragment (a single T→A transversion at the second Herein, we show that dispersed proximal promoter
ele-position of the TATA element). ments, denoted Gpal, interact synergistically with the
SREC 2/⫺425/lacZ, SREC 2.1/⫺425/lacZ, SREC 2.2/ SREC to activate transcription in several somatic
repro-⫺425/lacZ, and SREC 2.3/⫺425/lacZ were constructed by ductive organs. A nuclear protein, denoted GPAL, was PCR amplification of the appropriate intronic regions ofGld identified that binds specifically to the Gpal elements and inserted in the⫺425/lacZP-element vector.
Site-directed mutagenesis: Site-directed mutagenesis was and is present in tissues known to require the Gpal
performed using the mega-primer PCR mutagenesis method elements for Gld expression. Further mapping of the
(Picardet al.1994).SREC/⫺425dGpal⫺/LacZ, which already 3.3-kb region containing SREC revealed three distinct
contained four single nucleotide substitutions in the dGpal reproductive organ enhancers and an embryonic labial element, was used as a template. The LacZrev oligonucleotide enhancer unrelated to reproductive organ expression. (AAAGGGGGATGTGCTGCAAG), corresponding toLacZ re-The three reproductive organ enhancers activate ex- porter gene sequences, was used as the downstream (3⬘) primer. The PD6 oligonucleotide (TTGAGTTCGAtCgaTTCG pression in three different pairs of male and female
TCAGTT) served as the mutagenic primer to incorporate organs that are developmentally homologous. In
addi-three single nucleotide substitutions into Gpal-1 (lowercase tion we present evidence that the male ejaculatory bulb letters). The PD5 oligonucleotide (TAGGATCCGACCAAG medulla is homologous to the female spermathecae and TTTCACAGAGCGCAtCgaTGCGGCCA) served as the up-the ejaculatory bulb cortex is homologous to up-the female stream (5⬘) primer. PD5 also incorporated three single-nucleo-tide substitutions into Gpal-4 and adds a convenientBamHI uterus.
restriction site (underlined). The resulting PCR product, Although some details remain to be elucidated, we
⫺104 Gpal⫺was digested withBamHI andKpnI and inserted show herein a comprehensive explanation for the
com-into the unique BamHI/KpnI sites of SREC/LacZ, creating plex tissue-specific expression ofGldmRNA throughout SREC/⫺104Gpal⫺/LacZ.The⫺104Gpal⫺PCR product
frag-development. In addition these data provide an evolu- ment was subsequently sequenced to confirm mutation of the Gpal elements 1–4 and to confirm the integrity of the flanking tionary model for the sequential acquisition of
regula-Gldsequences. tory elements and their subsequent diversification with
Linker-scanning mutagenesis:Linker-scanning mutagenesis respect to controlling the expression of Gld in the
so-was performed using the PCR method developed byGustin matic reproductive organs among the species in the andBurk(1993). Oligonucleotide primers used were as fol-genus Drosophila. lows: For PCR product 1, Casper Gld.1 (CTAGAATTCCTGCA GCCGTTCG) and either LS1A (taggatccgacctcaAATCGGGA GCTTTTGAGTG), LS2A (taggatccgacctcaCTGCGCTCTCTG AAAC), or LS3A (taggatccgacctcaACAGCTTAAAGCTGGCC MATERIALS AND METHODS
GCAAA). For PCR product 2, Droadh.1 (GGTCAAAGTAAAC GACATG) and either LS1B (taggatccgacctcaGCAGCTTTGC
Plasmid construction:AP-element vector containing unique
GGCCAG), LS2B (taggatccgacctcaTGTCTTTCGTTGAGTTC EcoRI, BamHI, andKpnI sites between the SREC and a lacZ
GAG), or LS3B (taggatccgacctcaTTCGTCAGTTTAAAAAG reporter gene was constructed to facilitate insertion of various
ACT). For each of the LS oligonucleotides, the BamHI se-Gld promoter fragments examined herein. The 3.3-kbXbaI
-quence is underlined and Gld homologous sequence is in EcoRI SREC fragment fromGldintron I was blunt ended with
T4 DNA polymerase (Promega, Madison, WI) and inserted uppercase. The PCR products were digested withEcoRI and KpnI and inserted into the uniqueEcoRI/KpnI sites ofSREC/ in the reverse orientation into the blunt endedEcoRI site of
pCaSpeR-AUG-Gal (Thummelet al.1988), creatingSREC/ LacZ, creatingSREC/LS1/LacZ,SREC/LS2/LacZ, andSREC/ LS3/LacZ.Each PCR product was sequenced to confirm the LacZ.
The⫺104 to⫹84 and⫺39 to⫹84Gldpromoter fragments presence of the LS mutations and the integrity of the flanking Gldpromoter sequences.
were PCR amplified from SREC/⫺425/LacZ template. The
PCR products were digested withBamHI and KpnI and in- P-element-mediated transformation and isolation of trans-genic strains:P-element-mediated gene transformation of Dro-serted in the uniqueBamHI/KpnI sites ofSREC/LacZ, creating
SREC/⫺104/lacZandSREC/⫺39/lacZ.The entire PCR prod- sophila was performed essentially as described byRubinand Spradling(1982). Recombinant plasmids in the pW5-hsp70-ucts were sequenced in the final constrpW5-hsp70-ucts forSREC/⫺104/
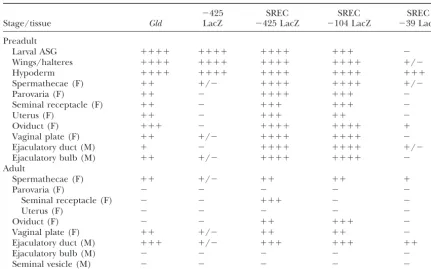
TABLE 1
5ⴕdeletion analysis of proximal promoter elements required for interaction with SREC in the reproductive tract
⫺425 SREC SREC SREC
Stage/tissue Gld LacZ ⫺425 LacZ ⫺104 LacZ ⫺39 LacZ
Preadult
Larval ASG ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹ ⫺
Wings/halteres ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹/⫺
Hypoderm ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹
Spermathecae (F) ⫹⫹ ⫹/⫺ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹/⫺
Parovaria (F) ⫹⫹ ⫺ ⫹⫹⫹⫹ ⫹⫹⫹ ⫺
Seminal receptacle (F) ⫹⫹ ⫺ ⫹⫹⫹ ⫹⫹⫹ ⫺
Uterus (F) ⫹⫹ ⫺ ⫹⫹⫹ ⫹⫹ ⫺
Oviduct (F) ⫹⫹⫹ ⫺ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹
Vaginal plate (F) ⫹⫹ ⫹/⫺ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫺
Ejaculatory duct (M) ⫹ ⫺ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹/⫺
Ejaculatory bulb (M) ⫹⫹ ⫹/⫺ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫺
Adult
Spermathecae (F) ⫹⫹ ⫹/⫺ ⫹⫹ ⫹⫹ ⫹
Parovaria (F) ⫺ ⫺ ⫺ ⫺ ⫺
Seminal receptacle (F) ⫺ ⫺ ⫹⫹⫹ ⫺ ⫺
Uterus (F) ⫺ ⫺ ⫺ ⫺ ⫺
Oviduct (F) ⫺ ⫺ ⫹⫹ ⫹⫹⫹ ⫺
Vaginal plate (F) ⫹⫹ ⫹/⫺ ⫹⫹ ⫹⫹ ⫺
Ejaculatory duct (M) ⫹⫹⫹ ⫹/⫺ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹
Ejaculatory bulb (M) ⫺ ⫺ ⫺ ⫺ ⫺
Seminal vesicle (M) ⫺ ⫺ ⫺ ⫺ ⫺
GAL expression levels for each transgene were visually scored on a⫹to⫹⫹⫹⫹⫹scale, with⫹representing very low level expression and⫹⫹⫹⫹⫹representing very high level expression. Minus denotes no expression in a particular tissue;⫹/⫺ denotes highly variable expression among strains or among individuals within a strain. F, female; M, male. At least three independent transgenic strains were analyzed for each construct and the tabulated results are the consensus of all strains. See Figure 1 for details of each transgene. GLD enzyme expression is included for reference.
were injected into preblastula stage embryos of theD. melano- To extract GPAL-1 from dissected tissues, the tissues of interest were dissected from the appropriate individuals in phosphate-gaster⌬2-3(99B) strain (Robertsonet al.1988). Crosses were
buffered saline solution, pH 7, and nuclear proteins were conducted to localize each transgene to a particular
chromo-isolated using the same methods for whole animal homoge-some, to replace the genetic background with a gal null
nates. mutant, to remove the endogenous transposase activity, and
Nuclear extracts were added to the assay buffer to give 30l to make the transgenes homozygous.
total volume and allowed to incubate at room temperature
-Galactosidase histochemical assays:Dissected Drosophila
for 30 min. The assay buffer was composed of 20 mmHepes tissues were assayed for-galactosidase (GAL) activity using
(pH 8), 100 mmKCl, 1 mmEDTA, 1 mmEGTA, 1 mm benza-a modified histochemicbenza-al stbenza-ain developed by Glaser et al.
midine, 1 mmphenylmethylsulfonyl fluoride, 20% glycerol, (1986). Three to five individuals for each transgenic strain
1 mg/ml BSA, 1g poly(dI)poly(dC)/1g GPAL-1 extract, were assayed at room temperature in the dark for 15 min to
1 ng gel-purified 32P-end-labeled oligonucleotide probe per
overnight as necessary. In addition, each strain was analyzed
reaction. The binding reactions were electrophoresed in a 5% at least twice in separate experiments.GAL expression levels
nondenaturing Tris-glycine polyacrylamide gel. Best results for each strain were visually quantitated on a⫹to⫹⫹⫹⫹⫹
were obtained with 0.5–1.5g GPAL-1 extract. Fold differ-scale, with ⫹ representing just detectable expression and
ences among lanes were estimated by scanning laser
densitom-⫹⫹⫹⫹⫹representing very high level expression.
etry (LKB Ultroscan XL).
Detection of sequence specific binding proteins:To charac-terize nuclear proteins that bind to Gpal four complementary oligonucleotide pairs were synthesized. The two
oligonucleo-tide pairs, dPalGld and mdPalGld, correspond to the sequence RESULTS of theGld promoter from ⫺82 to ⫺46. mdPalGld has four
mutations in the dPal, which destroys the palindromic nature Interaction between the Gld promoter and somatic of this sequence. The oligonucleotide pair dPalN contains the reproductive organ enhancer: We previously defined Gpal sequence flanked by 19 nucleotides unrelated to theGld an SREC within a 3.3-kbXbaI-EcoRI enhancer fragment promoter. The oligonucleotide pair mdPalN is identical to
from DrosophilaGld intron I (Keplingeret al.1996). dPalN except for four mutations in the dGpal sequence.
SREC can activate the⫺425- to⫹84-nucleotide (nt)Gld Protein extracts were isolated from pelleted nuclei of
Gldis normally expressed, both in the developing repro- SREC/⫺39/LacZ.Expression patterns were analyzed by histochemical analysis ofGAL expression in flies car-ductive tract (during metamorphosis) and in the mature
adult reproductive tract (Table 1). In addition, SREC rying the transgenes in a genetic background deficient for the endogenous -galactosidase gene. Three or activates expression of the⫺425/⫹84Gldpromoter in
the adult oviduct and seminal receptacle whereGld is more transgenic strains were analyzed for each trans-gene to rule out chromosomal background effects of not normally expressed at the adult stage (but where
Gld is expressed in preadult development). SREC can the random insertion site of the transgenes.
Removal of the ⫺425- to ⫺104-nt region in SREC/ also activate the heterologous Drosophila hsp70
pro-moter, but only in the adult male ejaculatory duct and ⫺104/LacZresults in the loss of expression in the adult female seminal receptacle but otherwise its pattern was to a lesser extent in the developing male ejaculatory
duct and female spermathecae. These observations sug- identical to theSREC/⫺425/LacZtransgene (Table 1). Deletion of the⫺104 to⫺39 region (SREC⫺39/LacZ) gested thatGldexpression in most of the reproductive
tract requires the SREC as well as proximal promoter resulted in a dramatic and broad attenuation of repro-ductive tract expression of the lacZ reporter gene. None-elements.
Delimiting theGldpromoter region necessary for re- theless, this transgene is expressed at moderate levels in the adult male ejaculatory duct. Much lower levels productive tract expression:To identify the promoter
region required in combination with the SREC for re- are observed in the developing and adult spermathecae and developing oviduct. Expression ofSREC/⫺39/LacZ productive tract expression, we constructed a 5⬘deletion
series of theGldpromoter with a common 3⬘terminus in the reproductive tract is similar to that of theSREC/ hsp70/LacZ transgene (Table 2) where only the adult located at⫹84 nt relative to the transcription start site
(Figure 1). The deletion series was designed to separate ejaculatory duct shows substantial expression (Figure 2b). In summary, the proximal promoter region from putative regulatory elements previously identified by us
(Krasney et al. 1990). Truncation of the region from ⫺104 to⫺39 immediately upstream of the TATA box is crucial for activation of theGldpromoter in the major-⫺425 to⫺104 nt removes the TTAGA elements, a
dis-persed repetitive family of elements. The TTAGA ele- ity of tissues in whichGld is normally expressed. How-ever, for some tissues this region is neither necessary ments, when tested independent of theGld, are capable
of activating a heterologoushsp70promoter at very low nor sufficient for Gldexpression.
The unique Gld TATA and Gpal elements are re-levels in the reproductive tract (Quineet al.1993), but
their requirement for Gld promoter activity had not quired for expression in a subset of reproductive or-gans:The TATA and dGpal elements are the most highly been tested. Truncation of the region from ⫺104 to
⫺39 nt removes a highly conserved 13-nt element (CAG conserved elements within several hundred nucleotides upstream of theGld transcription start site (Rosset al. CTTTAAGCTG) at ⫺68 nt exhibiting dyad symmetry
(Figure 1). This element is sufficient to activate expres- 1994). TheGldTATA element is positioned within the 28- to 31-nt range from the transcription start site as sion of a heterologoushsp70promoter in the anterior
spiracular glands of third instar larvae, in the wings and typically seen for Drosophila promoters (Cherbaset al. 1986); however, its sequence (TTTAAAAA) is atypical; halteres of developing adults during metamorphosis,
and in the adult male ejaculatory bulb (Gunaratneet T at the second position is rare in eukaryotic TATA boxes (Breathnach and Chambon 1981). Nonethe-al.1994). This element was previously named Gpal but
will be denoted herein as dGpal (dyad Gpal) and Gpal less, the unusualGldTATA sequence is highly conserved among the 11 Drosophila species examined (Krasney
will denote the AGCTTT core element. Two Gpal
ele-ments compose dGpal whereas other Gpal eleele-ments are et al. 1990; Ross et al. 1994). We speculated that this unusual TATA sequence may provide tissue specificity. dispersed as single elements in the proximal promoter
region ofGld. To examine the potential regulatory function ofGld unusual TATA box, we chose to change the T at the Each 5⬘ deletion promoter fragment was combined
with the SREC and placed upstream of theLacZreport- second position to an A, thereby creating a prototypical TATA consensus sequence (Figure 1). This mutation er gene in the P-element vector pCaSpeR-AUG-gal
(Thummelet al.1988), creatingSREC/⫺104/LacZand (mTATA) was examined in the fusion gene SREC/
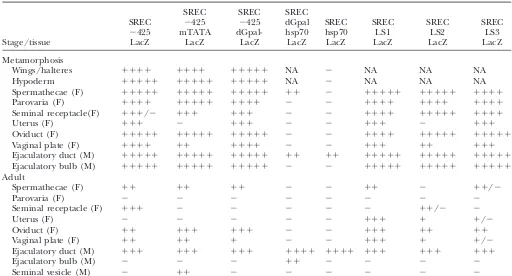
TABLE 2
Mutagenesis of theGldproximal promoter elements and their interactions with SREC
SREC SREC SREC
SREC ⫺425 ⫺425 dGpal SREC SREC SREC SREC
⫺425 mTATA dGpal- hsp70 hsp70 LS1 LS2 LS3
Stage/tissue LacZ LacZ LacZ LacZ LacZ LacZ LacZ LacZ
Metamorphosis
Wings/halteres ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ NA ⫺ NA NA NA
Hypoderm ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ NA ⫺ NA NA NA
Spermathecae (F) ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹ ⫺ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹
Parovaria (F) ⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫺ ⫺ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹⫹
Seminal receptacle(F) ⫹⫹⫹/⫺ ⫹⫹⫹ ⫹⫹⫹ ⫺ ⫺ ⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹
Uterus (F) ⫹⫹⫹ ⫺ ⫹⫹⫹ ⫺ ⫺ ⫹⫹⫹ ⫺ ⫹⫹⫹
Oviduct (F) ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫺ ⫺ ⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹
Vaginal plate (F) ⫹⫹⫹⫹ ⫹⫹ ⫹⫹⫹⫹ ⫺ ⫺ ⫹⫹⫹ ⫹⫹ ⫹⫹⫹
Ejaculatory duct (M) ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹ ⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹
Ejaculatory bulb (M) ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫺ ⫺ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹ ⫹⫹⫹⫹⫹
Adult
Spermathecae (F) ⫹⫹ ⫹⫹ ⫹⫹ ⫺ ⫺ ⫹⫹ ⫺ ⫹⫹/⫺
Parovaria (F) ⫺ ⫺ ⫺ ⫺ ⫺ ⫺ ⫺ ⫺
Seminal receptacle (F) ⫹⫹⫹ ⫺ ⫺ ⫺ ⫺ ⫺ ⫹⫹/⫺ ⫺
Uterus (F) ⫺ ⫺ ⫺ ⫺ ⫺ ⫹⫹⫹ ⫹ ⫹/⫺
Oviduct (F) ⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫺ ⫺ ⫹⫹⫹ ⫹⫹ ⫹⫹
Vaginal plate (F) ⫹⫹ ⫹⫹ ⫹ ⫺ ⫺ ⫹⫹⫹ ⫹ ⫹/⫺
Ejaculatory duct (M) ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹
Ejaculatory bulb (M) ⫺ ⫺ ⫺ ⫹⫹ ⫺ ⫺ ⫺ ⫺
Seminal vesicle (M) ⫺ ⫹⫹ ⫺ ⫺ ⫺ ⫺ ⫺ ⫺
⫺425mTATA/LacZ, otherwise identical toSREC/⫺425/ viously we showed that dGpal is capable of activating the heterologous hsp70 promoter in the male ejacula-LacZ.Analysis of the resultant transgenic strains showed
that the TATA element mutation abrogates GAL ex- tory bulb (Gunaratneet al.1994). However, this expres-sion did not require SREC. The fullGldpromoter (⫺104 pression in the adult female seminal receptacle and
developing female uterus (Table 2). Surprisingly, the to⫹84) containing dGpal does not support expression in the adult ejaculatory bulb. Taken together these re-mutant TATA transgene activates expression in the male
seminal vesicle, an organ for whichGldexpression has sults indicate that other elements in theGld promoter act to repress activation ofGld expression in the adult never been observed in any Drosophila species or in
any other of the numerous Gld transgenic strains we ejaculatory bulb. In addition, elements within the SREC enhancer region also repressGldexpression in this or-have studied. Therefore we speculate that a major
func-tion of the atypical Gld TATA is to repress Gld in the gan at the adult stage (see below).
Multiple segments of the promoter region are re-seminal vesicle.
We performed two experiments to analyze the high- quired for reproductive tract expression:A comparison of the 5⬘deletion analysis and the mutagenesis of the ly conserved dGpal element. Four nucleotide
substitu-tions were introduced into dGpal creating SREC dGpal and TATA elements suggested that other ele-ments must be present in the 65-bp region between ⫺425dGpal⫺/LacZ(Figure 1). The resultant transgenic
strain exhibited a lack of expression in the adult female ⫺104 and⫺39 nt that are required for expression in most of the reproductive organs that normally express seminal receptacle as compared to the nonmutated
SREC/⫺425/LacZ control (Table 2); however, expres- GLD during metamorphosis and in mature adults. A series of linker-scanning mutations in the ⫺104- to sion of GAL was observed in all other reproductive
tissues for which the control transgene is expressed. To ⫺39-nt region were constructed to identify such ele-ments. Each mutation replaces a 20-nt block of Gld test the sufficiency of the dGpal element to interact with
SREC in isolation of other Gld promoter elements, it sequence with random sequence (Figure 1). The LS1 mutation extends from⫺106 to⫺87 nt, LS2 from⫺82 was inserted upstream of thehsp70promoter inSREC/
hsp70/LacZ, creatingSREC/dGpal/hsp70/LacZ (Figure to⫺63 nt and LS3 from⫺59 to⫺40 nt.
In the adult reproductive tract, the LS1 mutation 1). Analysis of the resultant transgenic strains as
com-pared withSREC/hsp70/LacZshowed that the addition abolished GAL expression in the seminal receptacle (Table 2). Expression levels in other adult reproductive of dGpal resulted in activation of the reporter gene in
Figure 2.—Expression of the lacZ reporter controlled byGld transcriptional regulatory ele-ments. All transgenic lines bearing lacZ reporter constructs are homozygous for a null mu-tation of the endogenous Dro-sophila-galactosidase gene and thus the X-GAL staining pat-terns (blue) are solely due to the lacZ reporter. The patterns depicted in this figure and summarized in Tables 1–3 were shown in three or more inde-pendent transgenic lines. (a) SREC/⫺425/lacZtransgenic ex-hibits reporter expression in the embryonic antenno-maxil-lary complex (upper arrow) and labium (lower arrow). The pat-tern is identical to the patpat-tern of Gld mRNA and protein expression. (b) SREC/⫺39/ lacZ transgenic exhibits re-porter expression in the adult male ejaculatory duct (upper arrow) but not in the ejacula-tory bulb. This pattern is identi-cal toGldmRNA and protein. (c)SREC 2.1/⫺425/lacZ trans-gene exhibits reporter expres-sion in the ejaculatory bulb cor-tex (lower arrow) but not the ejaculatory duct. (d) ⫺425/ lacZtransgene exhibits expres-sion in the embryonic antenno-maxillary complex (arrow) but lacks expresexpres-sion in the labium. TheLBE/⫺39/lacZtransgene exhibits expression in the labium but not antenno-maxillary complex (not shown). (e) SREC/⫺425/lacZ transgene exhibits expression in the spermathecal caps (upper arrow) and seminal receptacle (lower arrow). (f )SREC 2.1/⫺425/lacZtransgene exhibits reporter expression in the adult female uterus.
SREC/⫺425/LacZ.The LS2 mutation abolished expres- noted that a dispersed sequence element (AGCTTT) in theGld proximal promoter region was identical to sion in the spermathecae and vaginal plate and resulted
in highly variable expression in the seminal receptacle. the core repeat element of dGpal and denoted these AGCTTT sequences as Gpal elements. We speculated The LS3 mutation abolished expression in the seminal
receptacle and resulted in highly variable expression in that these elements collectively interact with SREC to activate theGldpromoter in the reproductive tract. Four the spermathecae and vaginal plate.GAL expression
in the adult ejaculatory duct and oviduct were unaf- Gpal elements, including the two within dGpal, are pres-ent in the⫺104- to⫺39-nt region (Figure 1). Two addi-fected for all three linker-scanning mutations. The adult
expression patterns of the linker-scanning mutations do tional Gpal elements are located in the⫺425- to⫺104-nt region. To test the role of the Gpal elements in repro-not reveal distinct Gld promoter regions required for
expression in distinct subsets of adult reproductive tract ductive tract expression, we employed a combination of deletion and site directed mutagenesis to sequentially tissues, but rather multiple regions that collectively
con-tribute to expression in several tissues. For example, remove all six Gpal elements in the ⫺425-nt Gld pro-moter (Figure 1). The two upstream Gpal elements were linker-scanning regions 1, 2, and 3 are all required for
adult seminal receptacle expression. Linker-scanning removed in the previously described deletion that re-moves the⫺425- to⫺104-nt region. We performed site regions 2 and 3 are also required for adult spermathecae
and vaginal plate expression. No single sequence within directed mutagenesis on the four remaining Gpal ele-ments in the ⫺104-nt Gld promoter, creating SREC/ the⫺104- to⫺39-nt region is necessary for adult oviduct
expression, suggesting the presence of redundant ele- ⫺104 Gpa l⫺/LacZ.
In the adult reproductive tract, the SREC/⫺104 ments.
Dispersed Gpal elements interact synergistically with Gpal⫺/LacZtransgene expressesGAL only in the male ejaculatory duct, consistent with previous findings that the SREC and among themselves to produce the
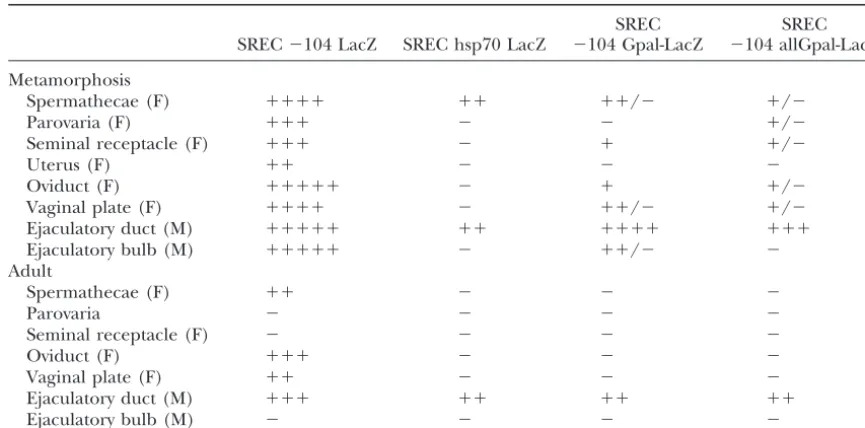
TABLE 3
Site directed mutagenesis analysis of the Gpal elements
SREC SREC
SREC⫺104 LacZ SREC hsp70 LacZ ⫺104 Gpal-LacZ ⫺104 allGpal-LacZ
Metamorphosis
Spermathecae (F) ⫹⫹⫹⫹ ⫹⫹ ⫹⫹/⫺ ⫹/⫺
Parovaria (F) ⫹⫹⫹ ⫺ ⫺ ⫹/⫺
Seminal receptacle (F) ⫹⫹⫹ ⫺ ⫹ ⫹/⫺
Uterus (F) ⫹⫹ ⫺ ⫺ ⫺
Oviduct (F) ⫹⫹⫹⫹⫹ ⫺ ⫹ ⫹/⫺
Vaginal plate (F) ⫹⫹⫹⫹ ⫺ ⫹⫹/⫺ ⫹/⫺
Ejaculatory duct (M) ⫹⫹⫹⫹⫹ ⫹⫹ ⫹⫹⫹⫹ ⫹⫹⫹
Ejaculatory bulb (M) ⫹⫹⫹⫹⫹ ⫺ ⫹⫹/⫺ ⫺
Adult
Spermathecae (F) ⫹⫹ ⫺ ⫺ ⫺
Parovaria ⫺ ⫺ ⫺ ⫺
Seminal receptacle (F) ⫺ ⫺ ⫺ ⫺
Oviduct (F) ⫹⫹⫹ ⫺ ⫺ ⫺
Vaginal plate (F) ⫹⫹ ⫺ ⫺ ⫺
Ejaculatory duct (M) ⫹⫹⫹ ⫹⫹ ⫹⫹ ⫹⫹
Ejaculatory bulb (M) ⫺ ⫺ ⫺ ⫺
promoter elements (Keplinger et al.1996). However, only a small additive effect by eliminating expression in the preadult ejaculatory bulb and by increasing line-as wline-as seen for theSREC/⫺39/lacZtransgene,
oblitera-tion of the Gpal elements reduces the expression in the to-line variability among most of the other remaining tissues in which theSREC/⫺104 Gpal⫺/lacZtransgene male ejaculatory duct indicating that the Gpal elements
are necessary for the normally high level ofGldexpres- was expressed.
Collectively, the three linker-scanning mutations elimi-sion in this organ. Expreselimi-sion of theSREC/⫺104 Gpal⫺/
LacZtransgene is undetectable in other adult reproduc- nate the Gpal sequences in the⫺104- to⫺39-nt region. LS1 is mutated for the Gpal-like elements at ⫺99 nt; tive tract tissues (female spermathecae, oviduct, and
vaginal plate), in contrast to SREC/⫺104/LacZ, with LS2 removes Gpal elements 2, 3, and 4. LS3 removes Gpal-1 and the Gpal-like element at⫺50 nt (Figure 1). four intact Gpal elements, or SREC/⫺425/LacZ, with
six intact Gpal elements. In the developing reproductive Assuming that most, if not all, of the effects of the linker scanning mutations are due to altering the Gpal tract, GAL is expressed in the male ejaculatory duct
(Table 3). However, in comparison toSREC/⫺104/LacZ elements, it is clear that no single Gpal element is re-quired for all tissues. Most tissues are relatively insensi-andSREC/⫺425/LacZ, expression is greatly diminished
in many developing reproductive tract tissues (female tive to loss of Gpal elements if the other Gpal elements in the⫺104- to⫺39-nt region are intact. However some parovaria, seminal receptacle, uterus, and oviduct) and
is both diminished and highly variable in others (male tissues (e.g., adult female seminal receptacle and vaginal plate) are particularly sensitive to mutations of one or ejaculatory bulb and female spermathecae and vaginal
plate). Thus, the repetitive Gpal elements are required more Gpal elements. LS2, which bears mutations in three Gpal elements including the highly conserved for synergistic interactions between the SREC and the
Gldpromoter and for activation in all adult reproductive dGpal elements at⫺72 nt, has the single most potent effect of eliminating expression in the developing tract tissues except the ejaculatory duct. Gpal elements
also play a key role in expression in the developing uterus and adult female spermathecae as well as reduc-ing the expression in vaginal plate (Table 2). Expression reproductive tract, although they are not strictly
re-quired in all tissues. in the developing uterus requires the unusual TATA box sequence as well. However, elimination of the dyad Two additional sequences (AGTTTC at⫺99 nt and
AGTTCG at ⫺50 nt) present in the ⫺104- to ⫺39-nt Gpal elements alone does not abrogate expression in the developing uterus.
region are very similar to the Gpal elements and may
account for the residual expression in the reproductive Proximal promoter regulation of Gld expression in nonreproductive organs: Previously we reported that tract seen in the SREC/⫺104 Gpal⫺/LacZ transgene.
To examine whether these two Gpal-like elements were the dGpal dyad repeat could drive expression of the heterologous hsp70 promoter in the anterior spiracular responsible for the residual expression, these sites were
functions in both reproductive and nonreproductive cleotide complexes were electrophoretically fraction-ated to observe potential mobility shifts in the probe as tissues. To further test this hypothesis we examined the
expression of theGldpromoter deletion transgenes de- a consequence of binding to specific proteins. Nuclear proteins isolated from the 100–300 mm NaCl nuclear scribed above. As with GLD, the⫺425/lacZandSREC/
⫺425/lacZtransgenes both show very high levels of re- fractionation revealed the presence of a prominent pro-tein that binds the 30-bp dPalGld (e.g., Figure 3a, lane porter gene expression in the larval anterior spiracular
glands (ASG) and pupal developing wings, halteres, and 1 and 3b, lane 1). We denoted this protein as GPAL. To determine the specificity of binding GPAL we per-hypoderm (Table 1). Reporter gene expression is
dra-matically reduced in theSREC/⫺39/lacZ transgene in formed a number of competition binding studies where the ability of mutant and nonspecific DNA fragments the ASG, wings, and halteres but is largely unaffected
in the hypoderm. These data are consistent with the were tested for their ability to inhibit the binding of dPalGld to GPAL. A mutated form of dPalGld, denoted hypothesis that the Gpal elements are necessary for
ex-pression in the ASG, wings, and halteres but that only mdPalGld, contains four substitutions in the 13-bp re-gion containing the dPal sequence. These substitutions the⫺39 to⫹84 region of theGldpromoter is required
for hypoderm expression. Hypoderm expression is still destroy the regulatory function of dPal (Gunaratne
maintained in the TATA mutant transgene (Table 2). We speculate that the region between⫹1 and⫹20 con-tains a hypoderm activation element inasmuch as it is
Figure 3.—Identification of GPAL, a Drosophila nuclear the most highly conserved region flanking the
transcrip-protein that specifically binds the dGpal element. Gel mobility tion start of theGld gene among nine Drosophila and shift experiments were conducted using nuclear protein ex-one Scaptodrosophila species analyzed to date (Kras- tract from one or more Drosophila developmental stages or tissues and double-stranded oligonucleotides corresponding neyet al.1990;Rosset al.1994;Merritt1998).
to regions of theGld promoter containing Gpal elements, A nuclear protein binds selectively to the dyad Gpal
isolated Gpal elements, mutant Gpal elements, or nonspecific element:Nuclear extracts from Drosophila embryos or
DNA fragments. The following double-stranded oligonucleo-specific tissues were isolated and probed with a radiola- tides were used in these experiments: dPalGld, 30 bp con-beled double-stranded 30-bp oligonucleotide (dPalGld) taining the 13-bp dGpal and nativeGld flanking sequences; mdPalGld, 30 bp containing four substitutions in dPal but corresponding to the dyad Gpal (dGpal) and flanking
otherwise identical to dPalGld; dPalN, 33 bp containing the sequences. All reactions were done in the excess of the
13-bp dPal sequence flanked by random sequences; mdPalN, unrelated poly dIdC competitor and other specific and
et al. 1992). Unlabeled dPalGld was able to compete GPAL ofD. melanogasterand was also found to be highly enriched in the male ejaculatory bulb (not shown). A effectively with labeled dPalGld when present in only a
20-fold excess (Figure 3a, lane 2) whereas a 450⫻molar high level of GLD is present in the male ejaculatory bulb ofD. virilis(Schiffet al.1992).
excess of mdPalGld (Figure 3a, lane 6) was required to
compete at an equivalent level. Therefore these four A 361-bp fragment of SREC contains two separate enhancers and a silencer: We previously reported the substitutions also ablated GPAL binding as well as dGpal
regulatory function. To further examine the importance presence of redundant enhancer elements dispersed in the 3.3-kb SREC that could activate reproductive organ of the flanking sequences surrounding dGpal two
addi-tional 33-bp oligonucleotides, dPalN and mdPALN, expression (Keplinger et al. 1996). Results from two overlapping fragments suggested that a strong adult were constructed. dPalN contains the native 13-bp dPal
sequence but has a randomized flanking sequence unre- ejaculatory duct-oviduct enhancer mapped in a 361-bp region as well as a more general somatic reproductive lated to theGldpromoter. mdPALN is identical to dPalN
but contains the same four nucleotide substitutions in organ enhancer that activates expression during the metamorphic development of these organs. This 361-bp mdPalGld. Binding affinity was reduced somewhat when
the sequences flanking dGpal were substituted with un- fragment (denoted SREC 2) as well as three small over-lapping subfragments (denoted SREC 2.1, SREC 2.2, related sequences (Figure 3b, lanes 7–12) indicating
that these flanking sequences are important for increas- and SREC 2.3) were inserted upstream of the⫺425 Gld promoter/lacZ P-element expression vector (Figure 4). ing specificity. The binding affinity was further reduced
when four substitutions were introduced in the 13-bp Replicate transgenic lines were obtained for each of the four constructs and the expression of lacZ was deter-region containing the dGpal element (Figure 3b, lanes
13–18). In summary, the dPal sequence is the most mined in adult reproductive tissues (Table 4). As pre-dicted, the 361-bp SREC 2 activated theGld promoter important determinant of binding GPAL to dPalGld,
but the flanking sequences also somewhat influence the in the adult male ejaculatory duct and female oviduct as well as the developing ejaculatory duct, the cortex of binding.
To further examine the specificity of binding of the ejaculatory bulb, oviduct, and spermathecae. Sur-prisingly, the 5⬘ most 110-bp subfragment (SREC 2.1) GPAL, three unrelated and unlabeled competitor DNAs
were tested for their ability to act as competitors with activated theGldpromoter in the adult male ejaculatory bulb cortex and female uterus, two tissues lacking ex-dPalGld for binding GPAL. These competitors,
includ-ing EF-1 (Figure 3b, lanes 19 and 20), calf thymus DNA pression in the SREC 2 transgenic lines. In addition SREC 2.1 activates reporter gene expression in the de-(not shown), and plasmid DNA de-(not shown), were
found to be 50- to 100-fold less efficient competitors as veloping ejaculatory bulb, uterus, and seminal recepta-cle. SREC 2.2 (middle one-third of SREC 2) did not compared to DNA fragments containing the wild-type
sequence of dPal. activate the Gld promoter in any reproductive tissue. SREC 2.3 activated theGld promoter in the adult and On the basis of our Gld promoter mapping
experi-ments (Quineet al.1993;Gunaratneet al.1994) and developing ejaculatory duct. Within SREC 2.3 is a 9-bp element (TTTTGAATG) similar to an ejaculatory duct experiments described herein, we predicted that the
GPAL protein should be present in several tissues. Two enhancer element present in the Drosophila Adh gene (Fanget al. 1991; Fangand Brennan 1992). Similar of these tissues, which can be readily dissected, are the
anterior spiracular glands of third instar larvae and the sequences are present in regulatory regions of two other genes,andropinandEst-6, that are expressed in the ejacu-male ejaculatory bulb. In addition we predicted that
the level of expression of GPAL should be high in em- latory duct (Karotam and Oakshott 1993; Sergeev et al.1995).
bryos as compared to third instar larvae and should be
substantially higher in adult males than in adult females. Overall the 361-bp SREC 2 is composed of two distinct tissue-specific enhancers, one for the cortex of the ejacu-Mobility shift experiments were conducted (as above)
using nuclear extracts of developmentally staged ani- latory bulb and the uterus (BCUE) and one for the oviduct and ejaculatory duct (EOE; Table 4; Figure 4). mals and hand-dissected tissues (Figure 3c). As
pre-dicted GPAL is at least an order of magnitude higher In addition, the SREC 2.3 fragment appears to repress enhancer activity of SREC 2.1 in the uterus (at both in embryos as compared to third instar larvae (Figure
3c, lanes 1–3vs.lanes 4 and 5). However, the anterior stages) and in the ejaculatory bulb cortex at the adult stage. This repressor element may correspond to an spiracular glands of third instar larvae are highly
en-riched for GPAL (Figure 3c, lane 6). Adult male extracts element distinct from the ejaculatory duct-oviduct hancer or the activity of the EOE may block the en-and extracts isolated from adult male ejaculatory bulbs
are also enriched for GPAL (Figure 3c, lane 13). A tissue hancer activity of the BCUE.
The developmental homology of the female and male subtraction experiment also shows that most of the
GPAL protein in the abdomen of adult males is present somatic reproductive organs is correlated with theGld
enhancer tissue specificity: The genital imaginal discs in the ejaculatory bulb (lane 13 vs. lanes 10–12). A
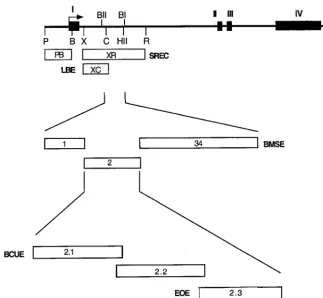
reproduc-Figure4.—Mapping the labial and so-matic reproductive organ enhancers in theGldgene. Transcript map of theGld gene. Solid rectangles represent exons I–IV. Thin line represents 5⬘ flanking sequences, introns, and 3⬘ flanking se-quences. B,BamHI; BII,BglII; BI,BglI; C, ClaI; HII, HincII; P, PstI; R, EcoRI; and X,XbaI. Open rectangles below the transcript map represent fragments ex-amined for enhancer activity. The labial enhancer (LBE) is localized to the XC fragment. The family of SREC en-hancers are located in the XR fragment and are composed of the BMSE (frag-ment 34), the BCUE (frag(frag-ment 2.1), and the EOE (fragment 2.3).
tive organs and the major function of the sex-determina- We show for the first time that a single enhancer (EOE) specifically activates gene expression in these two ho-tion pathway is to repress the opposite sex primordia
(EpperandNothiger1982). A variety of fate mapping mologous organs. Similar fate mapping experiments are somewhat less clear about the developmental homology and transplantation experiments of these primordia
have also shown that specific male and female organs are of the female uterus and spermathecae and male ejacu-latory bulb. We present herein new evidence showing serially homologous (Dobzhansky1930;Littlefield
andBryant1979;DobendorferandNothiger1982; that (1) the female uterus is homologous to the cortex of the male ejaculatory bulb, (2) the paired female
sper-ChenandBaker1997). Evidence for serial homology
is most compelling for the male ejaculatory duct and mathecae are homologous to the two hemispheres of the ejaculatory bulb medulla, and (3) the Gld gene female oviduct. Under appropriate experimental
condi-tions the primordia for each can be redirected to differ- contains two distinct enhancers that correspond to these two newly discovered developmental homologies. entiate into the organ normally found in the opposite.
TABLE 4
Deletion mapping somatic reproductive organ enhancers and silencers in the 361-bp SREC 2
SREC 2 SREC 2.1 SREC 2.2 SREC 2.3
Metamorphosis
Spermathecae ⫹⫹ ⫹/⫺ ⫹/⫺ ⫹/⫺
Uterus ⫹ ⫹⫹ ⫺ ⫺
Oviduct ⫹⫹ ⫺ ⫺ ⫹⫹
Ejaculatory bulb cortex ⫹⫹ ⫹⫹ ⫺ ⫺
Ejaculatory bulb medulla ⫹⫹ ⫺ ⫺ ⫺
Ejaculatory duct ⫹⫹⫹⫹ ⫺ ⫺ ⫹⫹⫹
Adult
Spermathecae ⫺ ⫹/⫺ ⫹/⫺ ⫹/⫺
Uterus ⫺ ⫹⫹⫹⫹ ⫺ ⫺
Oviduct ⫹⫹ ⫺ ⫺ ⫹⫹⫹
Ejaculatory bulb cortex ⫺ ⫹⫹⫹⫹ ⫺ ⫺
Ejaculatory bulb medulla ⫺ ⫺ ⫺ ⫺
BeloteandBaker(1982) isolated and characterized denote this enhancer as the ejaculatory bulb medulla and spermathecae enhancer (BMSE). The BMSE, simi-temperature-sensitive mutants of the transformer (tra)
gene, a gene required for female sexual development. lar to the BCUE and EOE, activates expression in homol-ogous male and female tissues.
They showed that if chromosomal female (XX)trats
mu-tants were reared throughout development at the re- An embryonic labial enhancer is present in the first intron ofGldadjacent to SREC:During embryonic de-strictive temperature (29⬚) they would develop as males
with normal male somatic reproductive organs whereas velopment GLD is expressed in the antenno-maxillary complex and labium. Previously we showed that the if they were reared at the permissive temperature (18⬚)
they would develop as females. We showed that if such ⫺425 to⫹84 promoter region ofGld was sufficient to drive expression in the embryonic antenno-maxillary females were reared at the restrictive temperature for
the first few days of development until the decision to complex immediately anterior to the labium (Figure 2d) as well as all other somatic tissues throughout preadult select the appropriate primordia for differentiation and
then shifted to the permissive temperature,Gldexpres- development. The embryonic labium is the only known nonreproductive organ in which this minimal promoter sion was misregulated in the resultant male reproductive
organs (Feng et al.1991). Specifically,Gld expression region is not sufficient forGldexpression. In the process of deletion mapping the SREC, we discovered an en-was derepressed in the ejaculatory bulb medulla.
Recip-rocal temperature shift experiments resulted in depres- hancer that activates expression in the embryonic la-bium (Figure 2a). The labial enhancer (LBE) was identi-sion of Gld expression in female oviduct. We report
herein that if these temperature shift experiments are fied in a 1.3-kb fragment located 639 bp downstream of theGldpromoter. When present with either theGld performed during the period when the decision to
se-lect the sex-appropriate primordia, elements of both promoter or a heterologoushsp70promoter, LBE acti-vates a highly specific and restricted expression in the male and female reproductive tissues are produced. In
several cases, we observed formation of hybrid repro- embryonic labium. Deleting all of the Gpal elements from theGldpromoter does not affect the ability of this ductive organs composed of fused spermathecae
sur-rounded by uterine tissue yielding a structure very simi- enhancer to activate the Gld promoter in this tissue. Thus despite the coincident expression of Gld in the lar to the male ejaculatory bulb medulla and cortex,
respectively (Figure 5, b–e) In these structures, the sper- antenno-maxillary complex and the labium, their tran-scriptional regulation is quite distinct, the former con-mathecal caps exhibit aberrant lateral extensions similar
to the horns of the ejaculatory bulb medulla. Most in- trolled by the proximal promoter elements and the lat-ter controlled by a downstream enhancer.
tersexual reproductive tracts generated were composed of both male ejaculatory duct and female oviduct that were both attached directly to the spermathecae. The
DISCUSSION spermathecae were partially transformed to ejaculatory
bulb medullar hemispheres. In normal males, the ejacu- Gpal elements drive expression in both reproductive and nonreproductive tissues:We previously showed that latory duct directly connects to the ejaculatory bulb
medullar hemispheres (Figure 5a). We propose that the dGpal element alone is sufficient to drive expression of a heterologoushsp70promoter in the anterior spirac-this hybrid organ is created by simultaneous activation
of the male and female genital disc primordia resulting ular glands of third instar larvae and in the adult male ejaculatory bulb of Drosophila (Gunaratneet al.1994). in the development of pseudoejaculatory bulb
com-posed of the two homologous female tissues, namely the GPAL, a nuclear protein that binds specifically to dGpal, is readily detectable by mobility shift assays in the ASG uterus forming the cortex and the paired spermathecae
forming the two hemispheres of the medulla. Previous and ejaculatory bulb. GLD expressed in the ASG is se-creted and performs an essential developmental func-fate mapping experiments (Littlefield and Bryant
1979; Dobendorfer andNothiger 1982) are consis- tion (CavenerandMacIntyre1983;Cox-Fosteret al. 1990). Although GLD is not normally expressed in the tent with this hypothesis although such studies did not
consider the homology of the ejaculatory bulb cortex adult male ejaculatory bulb of melanogaster, we found that aD. pseudoobscura Gldtransgene is expressed in this and medulla separately.
The BCUE activates expression in the male ejacula- organ at the adult stage when inserted into the genome of D. melanogaster (Schiff et al. 1992). Therefore it is tory bulb cortex and uterus, organs which we propose
are developmentally homologous. Previously we had not surprising that the dGpal element, taken out of context of otherGld regulatory elements, can activate identified an adjacent enhancer, downstream in SREC
(denoted segment 34 in Figure 4) that activates expres- expression in the melanogaster adult ejaculatory bulb. Presumably, silencer elements (present inD. melanogas-sion in male ejaculatory bulb and female spermathecae.
Reanalysis of these data and transgenic lines now show terbut notD. pseudoobscura Gld) normally block the acti-vation ofGldin the adult ejaculatory bulb. This hypothe-this segment activates expression specifically in the male
gene is derepressed in this organ upon partial derepres- gest that species-specific variation in adult expression patterns could be due to the unique threshold response sion of the female sex-determining factors in males
(Fenget al.1991). Despite the importance of both SREC of each speciesGld promoter to GPAL, determined by its unique number and quality of Gpal elements. GPAL and the dGpal element to reproductive tract expression,
it is interesting to note that dGpal is apparently incapa- has also been detected in the reproductive organs of two other Drosophila species,pseudoobscura andvirilis. ble of productively interacting with SREC to expand
expression of the heterologoushsp70promoter beyond Enhancer-promoter synergism:While theGldsomatic reproductive organ enhancer complex can activate a the reproductive organs that are independently
acti-vated by SREC and dGpal. Moreover, selectively mutat- heterologous promoter in the male ejaculatory duct similar to many other intronically localized eukaryotic ing the dGpal element within its normal context flanked
by other Gpal elements has a negligible impact on the enhancers (e.g.,Liuet al.2000), we found that the other reproductive tissues require a combination of proximal interaction of SREC and the Gld proximal promoter
region. In contrast, selective mutation of the dGpal ele- promoter elements and the SREC. This interdepen-dence may be the result of one or more mechanisms. ment obliterates expression in the larval anterior
spirac-ular gland, a developmentally essential site of GLD ex- SREC bound factors may make productive contacts with a Gldpromoter specific transcriptional initiation com-pression.
We speculate that over the course of evolution of plex, but not with a canonical transcriptional initiation complex (such as that of the hsp70promoter). Such a Gldfunction and expression, dGpal initially functioned
solely for expression in tissues essential for GLD func- mechanism has been suggested for the muscle-specific enhancer of the mouse myoglobin gene (Wefald et al. tion in cuticle metabolism and cellular immunity. By
the process of slipped-strand mutagenesis, the number 1990) and the Drosophila gooseberry and gooseberry neuro enhancers (Li and Noll 1994). This is an attractive of Gpal elements was expanded and SREC was
trans-posed to the first intron, thus adding expression ofGld model because, as previously noted, Gld has a highly conserved atypical TATA box sequence, TTTAAAA, to the reproductive organs.
Synergism among Gpal elements:Mutation of all six which might direct the assembly of aGldpromoter spe-cific transcriptional initiation complex. We tested this Gpal elements within the⫺425-ntGldpromoter results
in a dramatic attenuation of expression in all reproduc- hypothesis by converting the atypicalGld promoter to canonical TATA box and found that the reproductive tive tract tissues except the ejaculatory duct. Adult
ex-pression patterns are perturbed by progressive mutation tract expression was indeed abrogated in two female tissues. However, in all of the other tissues that depend of subsets of Gpal elements. Adult female seminal
recep-tacle expression is abrogated first, followed by sperma- upon specific interaction of the Gld promoter and SREC, expression was unaltered. Herein we have identi-thecae and vaginal plate expression and finally oviduct
expression. fied a cluster of repetitive sequence elements upstream of the TATA box denoted Gpal elements as indispens-The observed functional interactions among the Gpal
elements may arise from cooperative DNA binding or able for expression in this larger subset of reproductive tissues. Moreover, there is a strong functional synergism from simultaneous contact of different rate-limiting
components of the transcription initiation complex. We between these elements and the SREC. This synergism may be direct, wherein the respective factors interact suggest that synergism among Gpal elements is likely at
the level of cooperative DNA binding to GPAL since to bind DNA in a cooperative manner (Giniger and
Ptashne 1988;Janson andPettersson1990;Xiaoet Gpal elements are closely linked as required for
cooper-ative DNA binding (JiangandLevine1993;Szymanski al. 1991), or indirect, wherein the respective factors simultaneously contact different rate-limiting factors in and Levine1995). Several studies have identified the
number and quality of binding sites as key determinants the transcription initiation complex (Careyet al.1990;
Linet al.1990;JiangandLevine1993;Mannerviket reaching the critical threshold for transcriptional
activa-tion (Struhlet al.1989, 1992;Jianget al.1991). Alter- al.1999).
Why is SREC activation in most reproductive tract ation of binding sites reprograms the critical threshold
required for expression in a specific tissue. VariousGld tissues strictly dependent on Gpal elements whereas adult ejaculatory duct activation is not? One possibility promoter mutations that we have generated and tested
alter the number and/or quality of binding sites. Some is that the male ejaculatory duct is activated by a com-pletely separate set of elements present in SREC. Indeed tissues are considerably more sensitive to the loss of one
or more Gpal elements than others. The differential we have shown that SREC is composed of several dis-persed enhancer elements that activate expression in tissue sensitivity to alterations in Gpal elements may
reflect the level of GPAL in each tissue. Some of the different subsets of the somatic reproductive organs (Keplingeret al.1996). Further mapping experiments reproductive tract expression patterns displayed in Gpal
mutants ofD. melanogasterare reminiscent of the expres- reported herein show that a 140-bp region contains an ejaculatory duct and oviduct enhancer. All of the various sion patterns found in other Drosophila species, which
Figure 6.—A summary of the regulatory elements controlling the transcriptional regulation of Gldand a model of their interac-tions. (Top) Gld regulatory ele-ments controlling Gld tissue-spe-cific regulation are shown in the first row. A plus (⫹) indicates that the element is required to activate expression in the organ shown in the first column. A minus (⫺) in-dicates that the element blocks expression. With the exception of activation of expression in the semi-nal receptacle and repression of oviduct expression, the activity of the SREC complex is composed of the three reproductive organ enhancers, EOE, BCUE, and BMSE, and their interaction among them-selves and with theGldpromoter elements. (Bottom) Model of the interaction of theGldintronic en-hancers, silencers, the proximal promoter elements, and the pro-moter. The locations of these ele-ments have been mapped as de-scribed herein with the exception of the SEXS element which is only known to map within the 17.5-kb fragment defining the total Gld gene.
gene in the ejaculatory duct also activate expression in mRNA in the oviduct at the adult stage. We have been able to relieve the repression ofGld expression in the the oviduct. Only when these fragments are tested with
the heterologous hsp70 promoter do we observe expres- oviduct by transiently expressing male sex-determining factors immediately before differentiation of the oviduct sion restricted to the ejaculatory duct. However, it
should be noted that theGld promoter/SRECtransgene occurs, showing that the sex-determination pathway is responsible for the repression ofGldin the oviduct. activates a substantially higher level of expression than
does the hsp70/SREC transgene. Together these find- Somatic reproductive organ enhancers activate ex-pression in homologous organ pairs in the males and ings suggest that EOE trans-activating factors may be
sufficiently high in the adult ejaculatory duct to obviate females and exhibit epistatic interactions:We have iden-tified three distinct enhancers that activate reporter the need for Gpal elements in order to activate a
moder-ate level of expression but requires those elements for gene expression in specific pairs of somatic reproductive organs. Remarkably, each of these enhancers activates the normally very high level seen in this organ. An
unsolved paradox is why theGldgene is not normally expression in one female organ and one male organ. These male and female organ pairs were previously expressed in the adult female oviduct. We speculate that
although the oviduct requires bothGld promoter and shown or shown herein to be developmentally homolo-gous. To our knowledge such enhancers that activate enhancer elements, the presence of other elements in
reproduc-tive organs between the two sexes have not been re- medulla is mediated by the EOEs that both activate expression in the oviduct and ejaculatory duct and si-ported. The EOE has an additional function to silence
the activity of the nearby BCUE in the ejaculatory bulb lence the activity in the adult ejaculatory bulb medulla and female uterus. Of the five tissues that are activated cortex and uterus and the BCUE in the ejaculatory bulb
medulla at the adult stage. The close distance of the by the EOE, BMSE, and BCUE during preadult develop-ment, only the ejaculatory duct and spermathecae ex-EOE to the BCUE may be important for repression as
was shown to be important for the activity of the giant press GLD at the adult stage; therefore an additional silencing activity is required to specifically repress the repressor in Drosophila (Hewittet al.1999). It is
impor-tant to note that the EOE does not negatively effect EOE in the adult female oviduct. The elements responsi-ble for repression ofGldmRNA in this organ (denoted these other enhancers during preadult development.
The silencing activity of the EOE at the adult stage is sex silencers, SEXS) remain to be mapped within a 17.5-kb fragment that we showed contained all of the Gld in part controlled by the sex-determination pathway,
inasmuch as expression ofGld in the ejaculatory bulb regulatory elements including SEXS. Preliminary data indicate that SEXS is located in the distal half of the medulla is derepressed in males that have received a
brief pulse of feminizing factors by manipulation of first intron downstream of SREC.
With the exception of the somatic reproductive or-the sex-determination pathway. We speculate that this
occurs by specifically disrupting the silencing activity of gans, tissue-specific expression ofGldis highly conserved among the genus Drosophila (Schiffet al.1992). Func-the EOE on Func-the BMSE.
A summary of the tissue-specific regulatory elements tional conservation has also been conserved as demon-strated by the ability of theD. pseudoobscura Gldgene to controlling theGldgene and a model ofGld
transcrip-tional regulation is shown in Figure 6. Control of Gld rescueD. melanogaster Gldnull mutants (Krasneyet al. 1990;Schiffet al.1992). This interspecific gene trans-transcription in most of the nonreproductive tissues
(antenno-maxillary complex (AMC), ASG, hypoderm, formation experiment was particularly revealing with respect to the pattern differences in the adult somatic and wings) is exerted through the Gpal elements
inde-pendent of theGldenhancers. The temporal expression reproductive organs observed between these two spe-cies. The D. pseudoobscura Gld gene when introduced of Gld in these tissues is controlled by the ecdysone
as mediated through the proximal promoter elements into the genome of D. melanogaster was expressed in two organs, the ejaculatory bulb medulla and seminal
(Murtha and Cavener 1989 and our unpublished
data). However for the embryonic labium, tissue speci- receptacle, in which the host (D. melanogaster)Gldgene is not expressed at the adult stage. However, the host ficity is controlled exclusively through an enhancer
(LBE) and does not require Gld proximal promoter D. melanogaster Gldgene can be expressed ectopically in these two organs if these females are subjected to a short elements orGld’s unique TATA box. Regulation in the
somatic reproductive organs is much more complex. burst of male sex-determining factors immediately prior to their differentiation. In addition, the heterospecific All of these organs require to varying degrees the Gpal
elements with possible exception of the ejaculatory duct D. pseudoobscura Gld gene was expressed in the male ejaculatory duct of the hostD. melanogastermales, a tissue at the adult stage. The spermathecae and ejaculatory
bulb medulla are at one extreme since they are the in which theGldgene ofD. pseudoobscurais not normally expressed. Introducing the entire male ejaculatory duct only reproductive organs for which Gpal elements are
sufficient in the absence ofGldenhancers. At the other ofD. pseudoobscurainto aD. melanogasterhost via imaginal disc transplantation methods, however, showed that the extreme, the ejaculatory duct requires only an EOE.
However, despite their differences in regulation, all donor ejaculatory duct did not express the Gld gene. Collectively these experiments indicate that theD. pseu-three of these organs require both the Gpal proximal
promoter elements and intronic enhancers for high doobscuragene contains elements that can activate ex-pression in the adult ejaculatory duct but that the D. levels of expression. The seminal receptacle and uterus
are highly sensitive to mutations in Gpal elements or the pseudoobscura ejaculatory duct lacks the necessary cell autonomous regulatory factors to activate these ele-TATA sequence and the composition of the enhancer
elements. Although certain enhancer regions are re- ments. On the other hand, theD. pseudoobscura Gldgene appears to lack the silencer elements observed in the quired for seminal receptacle expression, we have not
been able to pinpoint a specific element that it requires. D. melanogaster Gld gene that block expression in the female seminal receptacle and male ejaculatory bulb Regulation ofGldin the male ejaculatory bulb and
fe-male oviduct are the most complex and intriguing. medulla.D. teisseri, a species much more closely related toD. melanogaster, lacks GLD expression in the ejacula-These organs normally expressGldmRNA during their
development but not at their sexually mature state. Re- tory duct and oviduct at both the preadult and adult stages but otherwise shows the same pattern of expres-pression of Gld expression at the adult stage in these
organs is predetermined during late larval-early pupal sion as D. melanogaster in other somatic reproductive organs (Cavener1985;Rosset al.1994). Future experi-development by the sex-determination pathway. The