The
what,
when,
where,
and
how
of
visual
word
recognition
Manuel
Carreiras
1,2,
Blair
C.
Armstrong
1,
Manuel
Perea
1,3,
and
Ram
Frost
1,4,51BasqueCenterforCognition,Brain,andLanguage,SanSebastian,Spain 2IKERBASQUEBasqueFoundationforScience,SanSebastian,Spain 3UniversitatdeVale`ncia,Vale`ncia,Spain
4TheHebrewUniversityofJerusalem,Jerusalem,Israel 5HaskinsLaboratories,NewHaven,CT,USA
Along-standingdebateinreadingresearchiswhether printedwordsareperceivedinafeedforwardmanneron thebasisoforthographicinformation,withother repre-sentationssuchassemanticsandphonologyactivated subsequently,orwhetherthesystemisfullyinteractive andfeedback from these representations shapesearly visual word recognition. We review recent evidence from behavioral, functional magnetic resonance ima-ging, electroencephalography, magnetoencephalogra-phy, andbiologically plausibleconnectionist modeling approaches, focusing on how eachapproach provides insight into the temporal flow of information in the lexicalsystem.Weconcludethat,consistentwith inter-activeaccounts,higher-orderlinguisticrepresentations modulate early orthographic processing. We also dis-cusshowbiologically plausibleinteractiveframeworks andcoordinatedempiricalandcomputationalworkcan advance theoriesofvisual wordrecognition andother domains(e.g.,objectrecognition).
Thewhat,when,where,andhowofvisualword recognition
Aviabletheoryofvisualwordrecognitionneedstospecify ‘what’ the building blocks of a printed word are and describe‘how’theyareprocessedandassembled togive risetowordidentification.Thesecentral‘what’and‘how’ questions have been the focus of research (and contro-versy)incognitive sciencesinceits verybeginning,and have traditionally been addressed by combining inven-tive experimental designs and reaction time (RT) mea-sures (Box 1). More recently, the availability of techniques such as functional magnetic resonance ima-ging(fMRI)haveprovidednewopportunitiestoask pre-cise ‘where’ questions, focusing on locating the neurocircuitry involved in recognizing printed words. Giventhearchitecturalconstraintsof thebrain,‘where’
information often tells us something important about ‘what’ types of representations are activated during visual word recognition and ‘how’ readers eventually recognizewords[1–3].
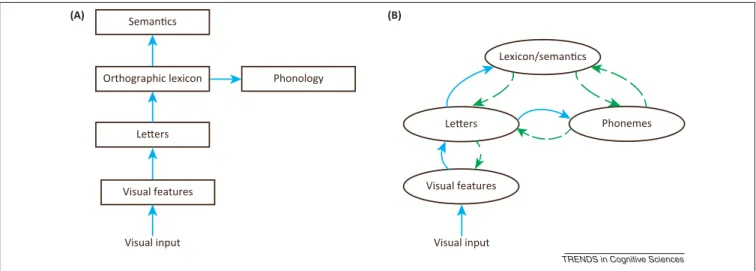
However, a comprehensive account of how complex stimuli suchas words areprocessed requires a detailed descriptionofthetemporalflowofinformationand eluci-dationof‘when’theinternalrepresentationsofwords(e.g., letters, syllables, morphemes, lexical entries) are acti-vated.Figure1presentscontrastingframeworks.Inthis respect,‘when’questionsconstrainanytheoryof‘how’by detailing thesequence ofeventsfromstimulus presenta-tiontowordrecognition.Infact,oneoftheoldestdebatesin visualwordrecognitionconcernsthedemarcationbetween bottom-upandtop-downprocessing,askingwhetherornot the visual stimulus feeds into thelexical level ina pre-dominantly hierarchical manner, wherein orthographic representationsfeedintohigher-levellinguistic represen-tations, or whether higher-level linguistic information such as phonologicaland morphological structureexerts a top-down influence on visual orthographic processing relatively early (Box 2). Cognitive neuroscience has rekindledthisdebate throughtheintroductionof techni-ques such aselectroencephalography (EEG) and magne-toencephalography (MEG), which have the appropriate temporalresolutiontotrackthetimecourseofprocessing. Note, however,thatthe‘where’,‘what’,‘how’,and‘when’ questionsaretoalargeextentinterdependent.Thehuman brain is generally constructed so that the trajectory of increased complexity, intermsof moving fromrelatively simple microfeature representations (e.g., the line seg-mentsinaletter)tocomplex,higher-orderrepresentations (e.g.,arepresentationofthewholewordform)is occipital-to-frontal,whereasthetrajectoryofhigh-levelmodulation isfrontal-to-occipital.Because‘where’informationis cor-related withthe flowof processing (early/simple or late/ higher-order),locationsofbrainactivationsareoftentaken tosupportclaimsregardingthetemporalorderof proces-sing. Here wediscuss thepotentialdanger of using evi-dence of ‘where’ to make inferences about ‘when’ (and ‘how’),reviewthefindingsobtainedusingtechniqueswith theappropriatetemporalresolutionfortrackingthetime courseofprintedwordprocessing,andpointtodesirable cross-fertilizationbetweenbehavioraldata,neuroimaging techniques,andneurobiologicallyplausiblecomputational 1364-6613/$–seefrontmatter
ß2013ElsevierLtd.Allrightsreserved.http://dx.doi.org/10.1016/j.tics.2013.11.005 Correspondingauthor:Carreiras,M. ([email protected]).
Keywords:visualwordrecognition;visualwordformarea;orthographicprocessing; neuralconnectivity;computationalmodeling;feedbackversusfeedforward informa-tion.
modelsforthedevelopment of amechanisticallyexplicit theoryofvisualwordrecognition.
fMRIevidencesuitablefor‘where’butnotfor‘when’
Many fMRI studies have investigated thebrain circuits thatunderliereading.Twopointsonwhichthisresearch convergeisthatthelefthemisphereplaysamajorrolein readingandthereadingcircuitconsistsofanetworkwith twomajor pathways: (i) adorsal pathwayincluding the occipital, supramarginal,and angulargyri, andthe pre-motorandparsopercularisintheinferior frontalcortex; and(ii)aventralpathwaythatintegratestheleftfusiform,
middleandanteriortemporal,andtheparstriangularisin theinferiorfrontalcortex[4].Thisnotwithstanding,there isstillaheated debateregardingthecharacterizationof directionalityofflowofinformationinthesepathways(i.e., ‘when’and‘how’).Specifically, theliteratureisunsettled regarding theextent towhich higher-level lexical repre-sentationsthatarenotnecessarilyorthographicmodulate therelativelyearlyprocessingoforthographicinformation (Box3).
Oneofthemostrelevantexamplesofsuchdebatesisthe role of theleft fusiform gyrus, the putative visual word formarea (VWFA) [5,6].From ananatomical processing
Box1.Measuringtimecoursesinbehavioralresearch
Although behavioral investigations are useful for understanding visual word recognition, these studies suffer from an inherent limitation:they onlyprovide anend-statereflection ofthe stateof processingviaanindirectbehavioralresponse(e.g.,lexicaldecision time as signaledvia a key press). Thus,thesetechniques do not providedirectinsightintotheinternaltemporaldynamicsunderlying ‘how’ different representations are activated. Moreover, these approachesruntheriskofproducingdatathatarecontaminatedby pre-andpost-lexicalprocesses(e.g.,decision-making).
Theselimitationsnotwithstanding,techniquesthatproviderelevant indirectinsightintothetimecourseofdifferentprocesseshavebeen developedthatrelatetothe‘when’questionregardingfeedforward and feedback processes. In this context, the masked priming technique[75]deservesspecialconsideration.Inmaskedpriming,a target word is preceded by a briefly presented masked priming stimulus(e.g.,mln-melon).Bymanipulatingthestructural relation-shipsbetweenprimeandtarget(e.g.,attheorthographic, phonolo-gical, morphological, and other levels) for different exposure durations(e.g.,typically between10 and60ms), researchershave observeddifferenttimecoursesofprocessingfordifferentproperties of printed words (e.g.,orthographic and phonological representa-tions)[76].
Therationalebehindthisexperimentalapproachisthattheminimal primedurationrequiredtoobtainaspecificprimingeffectreflectsthe
timenecessaryforactivationofthatinformation(e.g.,orthographic, phonological,morphological,orsemanticinformation).Nonetheless, this procedure has limitations [77], such as a lack of ecological validity.Arelatedandmoreecologicallyvalidtechniqueistopresent the words in the context of normal silent reading while the participants’eyemovementsareregistered[78].Ofparticularinterest istheveryearlyparafovealpreviewbenefiteffectusingtheboundary technique,inwhichtherelationshipbetween aparafovealpreview andatargetwordismanipulated.Specifically,theparafovealpreview isreplacedbythetargetwordoncethefixationcrossesaninvisible ‘boundary’locatednext to the targetword. Differencesinfixation durationonthetargetwordcausedbydifferentstructural manipula-tionsoftheparafovealpreviewreflect‘what’informationwasalready processedintheparafovea(e.g.,orthographyand/orphonologyand/ ormorphology)[79].
There is ample evidence that high-level information, such as phonological[80,81],morphological[82,83],andlexicalinformation
[84], influences very early aspects of the overall visual word recognitionprocess.Thisevidencechallenges thetraditionalclaim of temporaland structuralmodularity,according to whichprinted words are principally identified on the basis of orthographic information alone in skilled readers (the underlying logic behind some researchers’ concept of the VWFA), with phonological and semanticinformationretrievedsubsequently[64,85].
Visual features
Visual input Orthographic lexicon
Leers Leers
Semancs
Phonology
Visual input Visual features
Phonemes Lexicon/semancs
(A) (B)
TRENDS in Cognitive Sciences
Figure1.Corearchitecturalandfunctionalassumptionsoftemporallymodularfeedforwardversusinteractivemodelsofvisualwordrecognition.(A)Accordingto temporallymodularfeedforwardmodels,visualorthographicinformationisprocessedinasetofdistinct,hierarchicallyorganizedprocessingstages,suchthateachstage (e.g.,activationofletterandorthographiclexicalrepresentations)occursinastrictlyfeedforward–andinthestrongestform,sequential–fashion.Critically,additional non-visualorthographicrepresentations(e.g.,phonology,semantics)arenotaccesseduntilorthographicaccessiscomplete,and/orifaccessedbeforethatpoint,higher-level representationsneverfeedbacktoinfluencetheorthographiccomputation.(B)Accordingtointeractiveactivationmodels[59],visualinformationcontinuouslycascades throughouttheentireorthographic–phonological–lexical–semanticnetwork.Thisenablespartiallyresolvedphonologicalandlexical–semanticrepresentations(among others)tofeedbackandprovideconstraintsonother(lower)levelsofrepresentationinthenetworksuchasorthography.Notethatadditionalintermediatelevelsof representation(e.g.,letterclusters)havebeensuppressedforsimplicityinbothschematics,andthatthesearejusttwoexamplesofeachtypeofnetwork(e.g.,other feedforwardtheoriessuggestadirectsublexicalinputtophonologybutareneverthelessfeedforward).Unbrokenbluelinesdenotefeedforwardconnections;brokengreen linesdenotefeedbackconnections.
perspective(i.e.,a‘where’distinction),thisbrainregionis consideredto bearelatively‘early’ processingarea. The leftfusiformgyrusisactivatedmoreforwordsor pseudo-wordsthanforfalsefontsorconsonantstrings[7–10].Itis thuscommonlyacceptedthattheleftfusiformisinvolved inorthographic processing. There isacontroversy, how-ever,regardingwhatspecificinformationisrepresentedin this brain region and how sensitive it is to top-down information. One theoretical position is that the VWFA isaprelexical hub,specific for writtenwords,that com-putes and stores strictly visual and abstract prelexical orthographic representations in aprimarily feedforward manner [8,11,12]. Anothertheoretical position,however, postulatesthatactivationofthevisualformareais modu-latedbyhigher-orderlinguisticpropertiesofstimulisuch as phonology,morphology, and semantics [13,14]. These
two approaches provide very differentviews of reading: The formeriscompatible withthenotionof feedforward temporal (and structural) modularity (Box 2), whereby readingisconsideredtorelyonasequenceofconsecutive brainareassensitivetoahierarchyoforthographic repre-sentations(e.g.,letters,letterclustersofincreasingsize) thatculminatesinrecognitionofaword. Thelatter con-siders reading as a fully interactive processing system whereby higher-level linguistic information that is not necessarily orthographic modulates early perceptual orthographicprocessing.
Whereasproponentsofthefeedforwardapproachhave relied on theargument thatVWFA activation reflectsa stage oforthographic processing thatisimmune to pho-nologicalandsemanticinfluencesthatcomeintoplayonly lateron[12,15,16],thereismountingevidencesuggesting
Box3.FutureexplorationsofinteractivityusingfMRIandMEG
Severalstudieshaveshownintrinsicfunctionalconnectionsbetween Broca’sareaandventraloccipitotemporalregions[58,93–96] Anatomi-cal connectionsbetween frontaland occipital regions through the superior longitudinal fasciculus and/or the inferior fronto-occipital fasciculushavealsobeendocumented[97–105].Takentogether,these findings provide a neurobiologicalplatform forpossible top-down effectsfromfrontalareas,andthusofferinterestingavenuesforfuture investigations.Moreover,itisimportanttokeepinmindthatreadersdo notonly activatethe leftfusiform andother regionsof theventral pathway when reading. They also activate the dorsal pathway, includingregionssuchastheleftsupramarginalgyrus,theleftsuperior temporalcortex,andtheleftinferiorparietalcortex,andinparticular theangulargyrus,whichhasamodulatoryeffectonthevisualcortex
[4,106].Furthermore,thereareotherfunctionalpathwaysinthereading circuitstartingintheoccipitalcortexthatdonotnecessarilyinvolvethe leftfusiform[107].Thereadingcircuitincludesnotjustonebutmultiple
pathwaysfromvisiontohigher-ordertemporallobelanguageareas. Thus,thefullscopeofinteractivity(orlackthereof)betweenregions spanningthedifferentpathwaysofthereadingcircuitshouldbemore fullyestablished.Finally,itisimportanttonotethatthedorsaland ventralpathwaysarenotmodularsystemsthatoperateindependently ofeachother,butexchangeinformationduringvisualwordrecognition
[108,109].Infact,structuralconnectivitybetweenregionsbelongingto eachofthetwopathways(e.g.,theposteriorinferiortemporalregions, includingtheleftfusiform,andtheposteriorsuperiortemporaland inferiorparietalregions,includingthesupramarginalgyrus)hasbeen documented [110].Moreover,functionalconnectivitybetweenthese regions hasbeenshowninskilledreaders[111]butnotindyslexic individuals[112].Furtherstudiesarerequiredtodeterminehowbrain regions falling alongthe ventral and dorsalpathwaysinteract and cooperateduringvisualwordrecognition,andhowtheseinteractions relatetoothersimilarprocessessuchasobjectrecognition.
Box2.Structuralandtemporalmodularity,interactivity,andorthographicprocessing
Themaintheoreticalstandunderlyingthefeedforwardapproachis thatpureorthographicmodelshavesubstantialdescriptiveadequacy toaccountforalargesetof(mostlybehavioral)datainvisualword recognition[86].Onebasictenetinthisfeedforwardviewisthatin principle,‘feedbackcannotimproveperformanceateitherthelexical or prelexical level’ [86,p. 306] and a number of well-known phenomena(e.g.,wordsuperiorityeffect)thathavebeentraditionally attributedto top-downfeedbackcanindeedbeexplained parsimo-niouslyinafeedforwardmanner.Accordingtothisview,thegameof processing printed words islargely played inthe courtof ortho-graphic processing, suchthat a significantpart ofthe recognition processisdeterminedbyconsideringthesurfacestructuralproperties oftheprintedstimulusalone(i.e.,letters,lettersequences). Interest-ingly,thispositionisconsistentwiththeideathattheidentificationof visualformsingeneralandletterstringsinparticularcanbeachieved through a low-level visual pattern recognition system shared by humansandbaboons[87–89].
Thestrongestversionofthefeedforwardviewpostulatesstructural modularity, according to which orthographic processing is in principlenon-penetrablebyother linguisticdimensions.Moreover, processingwithintheorthographicsystemproceedsbottom-upfrom low-level featuresto full orthographicwords. The weakerversion assumestemporalmodularity[90]andpositsthattheword recogni-tion systemissimplysetsothat theprocessing of printedwords proceedsuntilanorthographicwordunitisrecognized; onlywhen this is accomplished does the orthographic representation make contact with various other linguistic properties (e.g., phonology, morphology,semantics[64]).Notethatthisapproachmayallowfor
top-downinteractivityaslongasitisconstrainedtooccurafterinitial orthographiccodingiscomplete.
The contrasting approach argues for full interactivity between lower-andhigher-orderrepresentationsatallprocessinglevels.Here, thedemarcationlinebeyond‘when’and‘where’‘perceptual ortho-graphic’processingendsand‘linguistic’processingbeginsisblurred
[72].Accordingtothisview,high-levellinguisticconsiderationsthat are not purelyorthographic(e.g.,how some letterscorrelate with phonologyandmeaning,andhowletterclustersareconstrainedby lexical, morphological,and phonologicalstructure) shapethe dis-tributional propertiesofletters inagivenlanguage,andthe word recognitionsystemlearnsthesefeaturestoenableefficient(i.e.,fast and accurate)reading in thatlanguage. Language-specific retinal– perceptuallearningeffects(i.e.,cross-linguisticdifferencein proces-singlettersatdifferentretinaleccentricities)[91]suggestthatreading habits stemming from the overall structure of alanguage indeed affectthefunctionalstructureofearlystagesinthevisualpathway, and are thus compatible with this view. For example, frequently encounteredvisualconfigurationsresultinperceptuallearningthat allowsforrapidandefficientrecognitionofawordform,andthese configurationsareinfluencedbythecorrelationoforthographywith phonology andmeaningthatarecharacteristic toalanguage[92]. Becausedifferentlanguagesarecharacterizedbydifferentrelations between orthography, phonology, and semantics (among other representations), itisarguedthatinteractivemodelsthatallowfor phonological,morphological,andsemanticinformationtocomeinto playearlyonarebetteraccountsforthesubstantialcross-linguistic differencesobservedinearlyorthographicprocessing[85].
thatearlyprintprocessingintheVWFAismodulatedby high-levellexicalinformation.Forexample,sensitivityto high-levelvariables suchas lexical frequency has been observed in the left fusiform [17]. Furthermore, the VWFAwassimilarly activatedwhentargetwords were precededby maskedprinted-word primesor bymasked pictures[18](Box1).Notethatbidirectionalinformation flowwherebyhigher-orderlevelsofprocessingconstrain feedforwardassemblyhasalsobeen proposedforobject recognition [19–24]. Adopting the recycling hypothesis [12](i.e.,theneurocircuitryforvisualobject recognition isrecycledtocompute therepresentationsnecessaryfor humanreading),simpleparsimonyconsiderationswould leadtotheassumptionofsimilarprinciplesregardingthe flow of information for visual object and visual word recognition.
Despite the above evidence, the debate regarding whetherprocessing of visualword recognitionis feedfor-wardornotisstillasactiveasever.Thisisbecausethe criticaldistinctionbetweenthetwoconceptualapproaches regarding‘how’informationflowsinthecircuitsismostly temporalinnature(‘early’vs‘late’).However,fMRI inte-gratesprocessesoverarelativelylongperiodoftime.Thus, althoughthetimingofstimuluspresentationcanbewell controlled (e.g., masked priming, fMRI adaptation), the temporal resolution of the blood-oxygen-level dependent (BOLD)responseistooslowtounequivocallydistinguish betweenactivationsthatarefeedforwardversusfeedback (i.e.,the‘when’question)[25].Thisleadsustotheinherent advantageofMEGandEEG.
MEGandEEG:thedynamicsofthetimecourse
EEGandMEGaretime-sensitivemethodswithatemporal resolution in the range of milliseconds. Combined with appropriate designs, theycan reveal thetemporal order oftheneuralprocessesinvolvedinvisualwordrecognition, tracingthetimecoursefromlow-levelvisualperceptionto letterperceptionandwordmeaning.Itisthistimecourse thatprovides important evidence to adjudicate between differenttheoreticalstancesregardingtheflowof informa-tion(feedforward vsfeedback).Bothtechniques tap syn-chronized neuronalactivityover timetriggered by some cognitiveeventinthebrain[26].UnlikeRTs,whichgiveus theendresultofprocessinginthesystemasawhole(and notspecificallyoflexicalprocessing;Box1),bothMEGand EEG provide a continuous measure of the intermediate events that have led to the final response. MEG also provides some spatiotemporal constraints, allowing for somesynthesisof‘when’and‘where’information.
Oneoftheearliermarkersofvisualwordrecognitionisa left lateralized N150/N170 response that differentiates orthographicstimulisuchaswordsandpseudowordsfrom otherstimulisuchassymbols[27–29].Selectiveresponses tolettersinthistimewindowhavealsobeenfoundinthe inferioroccipitotemporalcortexusingintracranial record-ings [30,31] and MEG [32,33], particularly for normal readers but not for dyslexic children [34–37]. Thus, it has been proposed that the left-lateralized N170 could bean automaticresponse related to typicalvisual word recognition,andthatitcouldbeassociatedwiththe activa-tionfoundusingfMRIintheVWFAleftfusiformgyrus[38]
(butseeRauscheckeretal.[39]forbilateraleffects).Infact, allthisevidencesupportstheclaimthatatapproximately 150ms fromstimulus onset,thevisual system responds only to the frequency of letter combinations, and that lexicalandphonologicaleffectscomeintoplaymuchlater [15,16,40].Asexpoundedbelow,however,higher-level lin-guisticinformationalreadyexertsitsinfluenceat100ms (i.e.,before170ms)fromstimulusonset.Forinstance,it hasbeenreportedthatearlyevent-relatedpotential(ERP) components in the range 100–200ms are sensitive to lexicalfrequency[41–45].Thus,fromasimpletime-scale perspective,anearlymarkerofvisualwordrecognitionas revealedbyERPmeasures(butnotfMRImeasures)seems tobesusceptibletomodulation fromhigher-order lexical information.
AnotherearlymarkerofreadingistheN250,whichwas originallyfoundtobesensitivetoorthographicsimilarity in combined masked priming and EEG studies [46,47]. However, subsequent studies have shown that N250 is alsomodulatedbylexicalfactors[48,49].Inparticular,it wasfoundthatthisERPcomponentissensitivenotjustto letter identitybut also to the phonological status of the letters, thatis, whether letters areconsonant or vowels [46,50–52].Forexample,Carreirasandcolleaguesshowed that masked subset priming of consonants (e.g., mln– melon) and masked full identity priming (e.g., melon– melon) donot significantlydiffer fromeachother inthe N250component,whereasmaskedvowelsubsetpriming (e.g.,aio–amigo)andmaskedidentitypriming(e.g.,amigo– amigo) do [46]. Because consonants are more lexically constraining than vowels in predicting word identity [53],thiseffectdemonstratesthattop-downlexical infor-mation modulates the N250 component. Note that the same pattern of response is revealed in the later N400 component and in RTs in behavioral experiments. This suggeststhataccumulatedlexicalinformation(and/or lex-ical competition) that generates the masked prime has exerteditsfullimpactby250msfromstimulusonset.In fact, the dissociation found between transposed-letter priming effects for word–wordpairs(e.g., casual–causal) andfornonword–wordpairs(e.g.,barin–brain)intheN250 component [48] reinforces the hypothesis of high-order lexical–semantic information constraining orthographic form-levelprocessingintheN250.
Consistentwithsustainedandearlyinteractive coacti-vationofanetworkofsitescontributingtoreading,Thesen and colleagues found strong phase-locking from 170 to 400msbetweentheleftfusiform andmoreanterior lan-guage areas when comparing words versus false fonts usingMEGandintracranialrecording[54].Otherrecent reports ofveryearly neurobiologicalresponsesto phono-logical information inanterior areasare also consistent with atop-down flowof informationduring visual word recognition. UsingMEG inamaskedpriming paradigm, Wheatandcolleaguesobservedstrongerresponsesto pseu-dohomophonesthantoorthographiccontrolprimeswithin 100msoftargetwordonsetinaclusterthatincludedthe leftinferior frontalgyrus (pars opercularis)andthe pre-central gyrus [55–57]. Note that a parallel pattern of activation found in the middle occipital gyrus suggests thatthese regions could oscillate together during visual
wordrecognitionataveryearlystage.Thus,theinferior frontal gyrus may exert feedback control on regions involvedinlower-levelanalysisofwrittenwords.Infact, arecentstudyprovidesevidenceoftop-downfeedbackfrom theinferior frontalgyrus to theleftventral occipitotem-poral cortex via dynamic causal modeling of MEG data [58]. Specifically, the researchers found that words (as comparedtofalsefonts)activatedtheleftinferiorfrontal gyrus.Moreimportantly,theyshowedthatfeedback con-nectionsfromtheinferiorfrontalgyrustotheleftventral occipitotemporal cortexwithin the first200msprovided the best fit for the data relative to a model with only feedforwardconnectivitybetweentheseregions.One pos-sible explanation for this feedback is that the inferior frontalregionsendshigher-levelinformation(e.g., phonol-ogy)toconstraintherepresentationscomputedintheleft fusiform. Alternatively (or complementarily), these two brain regions may be interacting bidirectionallyas part ofaconstraintnetworkwithas-yetunderspecifiedgraded specialization across the different contributing brain regions. Although the specific representations and dynamics of the frontal–occipitotemporal areas remain to beelucidated, thesedata clearlychallenge thenotion of temporal and structural modularity in orthographic processing.
Biologicallyplausibleconnectionistmodeling:a platformforadvancingtheoriesofvisualword recognition
From the empiricaldata outlined above, itis clear that substantive theoretical advances will require an inte-grated understanding of thecontributions of a large set of distributed representations stored in different brain regions that are accessed (at least initially) at different pointsintimeasactivitycascadesthroughoutthebrain. Connectionistmodelingoffersamechanisticplatformthat isideallysuitedfortheseinvestigationsbecauseitallows researcherstoprobethe‘where’and‘when’ofvisualword recognition and directly relate them to thequestions of ‘what’(representations)and‘how’(explicitcomputational processingmechanisms)workinginconcerttoenablethe perception of written words. Moreover, models allow researchers to explore the emergent properties of these systemsanddeveloptargetedempiricalresearchagendas forthefuture.
The basic capacitiesof connectionist networksas out-linedabovewerekeenlydemonstratedviatheinteractive activationmodel[59–61].Inthismodel,informationfrom low-level visual featuredetectors flowed bottom-up to a lexical representation of whole words, while simulta-neouslybeingabletoflowtop-downfromhigherlevelsof representation.Thismodelcanthusexplainandgenerate predictionsregardingtop-downinfluencesrelatedtoword reading,suchasthewordsuperiorityeffect(i.e.,the pro-cessingadvantageforlettersembeddedinwordsrelativeto isolatedletters)[62].
Fromthisworkandthegeneralmechanicsofconstraint satisfactionsystems[63],aswellasthepresenceof bidir-ectionalconnectivity betweenbrain regionsthat process differentaspectsofwordrepresentation(e.g.,letters, pho-nology, semantics),connectionist theories of visual word
recognition will, by default, expect and generate some degreeoftop-downinfluenceformaximallyaccurateword identification.Amorecriticalconcernisifthesetop-down influencesaresubstantiveenoughtobetheoretically sig-nificantandmaynotbedismissedforreasonsofparsimony [64–67]. Proficient reading involves optimization of effi-ciency inadditionto accuracy,thatis,correct readingof wordsasquicklyaspossible.Thus,ifastrictlyfeedforward system could, in principle, enable highly efficient word recognition, under what circumstances would the brain choosetopaythepriceofwaitingforadditionaltop-down constraints becauseof inadequacies inastrictly feedfor-wardsignal[68]?
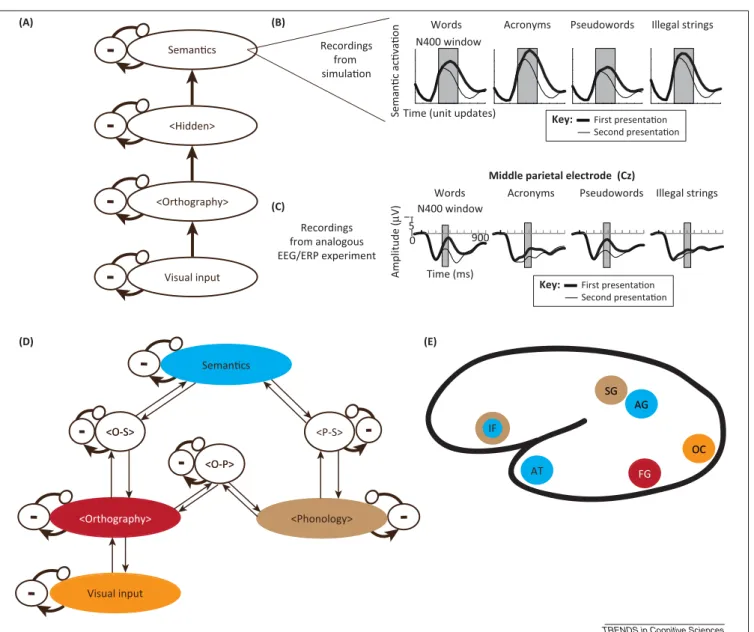
Connectionistmodelsofferseveral avenuesfor explor-ingthesepossibilities.Oneparticularlyimportantrecent advanceistheabilitytoincorporate additional neurobio-logical constraints into standard connectionist models (e.g., byspecifying differentsubpopulations of inhibitory and excitatory neurons) to simulate electrophysiological and behavioralresponses (B.C.Armstrong,Ph.D.thesis, Carnegie Mellon University,2012) [69,70].For example, Laszlo and Plautshowed howa modelthat instantiates theseprinciplescangenerateandexplain electrophysiolo-gicaldynamicscorrespondingtotheN400ERPcomponent [69].In addition, theywere able to advance thefield by offering an account of an important discrepant finding between behavioral andelectrophysiological approaches: why the N400 ERP component is not sensitive to the lexicality of the stimulus (e.g., words and pseudowords vs acronyms and illegal strings), whereas behavioral responses are. Specifically, they showed that the initial settlingdynamics,duringwhichtheprominentdeflection typically associated withthe N400 ERPcomponent was displayed,wereprimarilydrivenbytheorthographic word-likenessofthestimulus(e.g.,intermsofitsorthographic neighborhood). However,nonlinear settling dynamics in thenetworkcausedachangeintheseactivationpatterns lateroninprocessing,suchthatvalidlexicaltypes(words andacronyms)weremoreactivethannonwords (pseudo-words and illegal strings), consistent with typical beha-viorallexicaldecisiondata.
Laszlo and Armstrong further extended this work to accountfor howsimple contexteffects (e.g.,word repeti-tion) modulatetheN400componentassociated with lex-ical–semantic access [70]. This was accomplished via incorporation of a neuron-specific fatigue mechanism so thatrecentlyfiredneuronswouldnotbeabletofireattheir maximumrateforabriefperiodoftime.Thisresultedina reductioninN400-componentamplitudesforstimuliinthe semanticrepresentation,regardlessofthelexicalstatusof thecharacterstringinputtothenetwork.Moreover,they wererecentlyabletogeneratespecificpredictions regard-ingthepower-frequencyspectrathatshouldbeevokedby words and nonwords (Laszlo and Armstrong, unpub-lished),datathatareincreasinglyinfluentialin establish-ingthecausallinksbetweenwhichbrainregionsinfluence oneanotherandthetemporalorder(i.e.,‘when’)inwhich this occurs(Figure2) [58,70]. This workled to targeted insightsinto ‘what’aspects of aword representationare modulatedbyrelatedcontext.Furthermore,thisneurally inspired account therefore presents an alternative
explanationfor thelong-acceptedverbalaccountofN400 repetition effects [71], accordingto which reduced N400 amplitudes results from an (underspecified) facilitation mechanismratherthanafatiguemechanism.
Withsuchexplicitmodelsinhand,itispossibletoaddor subtract different feedback connections and evaluate whichofthesemodelsbestcapturesempirical electrophy-siological data such as ERP waveform amplitudes and powerovertimeinasparsimoniousamanneraspossible. Thesemodelsthuspresentanopportunityforan ‘experi-mental’approach totheoreticaldevelopmentbyallowing
direct assessment of the impact of different theoretical assumptions. Moreover,by virtue of thedomain-general natureoftheframework,itispossibletonaturallyextend theseprinciplestothestudyofotherlevelsof representa-tion.Forinstance,theseprinciplescanbereadilyapplied tostudyERPcomponentsassociatedwithearlierprocesses relatedtovisualwordrecognition(e.g.,N170,N250)and determine ‘when’ and ‘how’ these representations shape otherpurportedlyearlierprocesses.
Combinedwithadomain-generallearning theoryand recent advances in ‘deep’ neural networks [72], it has
(A) (B)
(C)
(D) (E)
Visual input
Semancs <Orthography>
<Hidden>
Time (unit updates)
First presentaon
Key:
Second presentaon
Semancs N400 window
Illegal strings
Words Acronyms
Semanc acvaon
Pseudowords Recordings
from simulaon
Recordings from analogous EEG/ERP experiment
Amplitude (
μ
V)
Words Acronyms Pseudowords
Middle parietal electrode (Cz)
Illegal strings N400 window
0 5
900
Time (ms)
OC
OC
FG
FG
AT
AG
AG
SG
SG
Visual input <O-S> <O-S>
<Phonology> <O-P>
<O-P>
<P-S>
<Orthography>
IF
First presentaon
Key:
Second presentaon
TRENDS in Cognitive Sciences
Figure2.Integrationofinsightsfrommorebiologicallyplausibleconnectionistmodelsandneuroimagingdata.Recentconnectionistmodelsthatuselargepoolsof excitatoryneuronsandsmallpoolsofinhibitoryneurons(here,inhibitorysubpopulationsaredenotedby–;allotherneuronsareexcitatory),aswellassparse/weakdistal connections(thinarrows)anddense/stronglocalconnections(thickarrows),andemergenthiddenrepresentationsthatarenotexplicitlyspecifiedbythemodeler(denoted by<>aroundthenameoftheapproximaterepresentationthatshouldemergeinthatpool)haveledtomajoradvancestowardsexplainingandpredictingbehavioraland neuroimagingdatawithacomputationallyexplicittheory.Forexample,LaszloandPlaut[69]andLaszloandArmstrong[70]usedthearchitectureillustratedin(A)to generate(B)simulatedERPcomponentsthatexplainempiricalERPdatafrom(C)ananalogousexperimentinvolvinglexicaltypesincludingwords(e.g.,DOG),acronyms (e.g.,DVD),pseudowords(e.g.,DOD),andillegalstrings(e.g.,XFQ)bothontheirinitialpresentation(thicklines)andwhentheitemwasrepeated(thinlines).Notethatthe unitsforthemodeldataarearbitraryandareomitted;fortheempiricaldata,thex-axisrangesfrom0to900msandthenegativedirectionisplottedupwards.Allempirical andsimulationdataweredrawnfromLaszloandArmstrong[70],usedwithpermission.Theseevent-relatedpotentialdatawerecollectedfromanelectrodeplacedona middleparietallocation.Thisworksuggeststhat(D)inamorecomprehensivemodelthatalsocontainsorthographicandsemanticrepresentations(herethe<o-p>,<o-s>, and<p-s>labelsdenoteintermediatepoolsofneuronsthatmapbetweenorthography,phonology,andsemantics,respectively),itisinprinciplepossibletostudythe activationdynamicsandrepresentationsthatemergein(E)brainregionsassociatedwithdifferentrepresentations,ofwhichasubsetofthemostcriticalregionsareshown inalateralcross-sectionofthelefthemisphere.Thecolorofthecirclesdenotesthetheoreticalrepresentationsinthemodelthattheseregionsmightsubserve. Abbreviations:IF,inferiorfrontalcortex;SG,supramarginalgyrus;AG,angulargyrus;AT,anteriortemporalcortex;FG,fusiformgyrus(includesvisualwordformarea, VWFA);OC,occipitalcortex.
recentlybecomepossibletotrainnetworkswithmultiple intermediate‘hidden’layerswhoseinputvaluesortarget outputsarenotexplicitlysetbytheexperimenter. Conse-quently,researcherscanstepbackfromexplicitly specify-ing the exact nature of the representations that are expectedtomediatebetweenalow-levelvisualformand higher-orderlexicalinformation.Instead,theycansimply specifyamoreneutralinput(e.g.,alow-levelvisualinput associatedwithawordinaretinotopicallycenteredcoding scheme) and observe how unsupervised learning mechanicscangraduallyextracthigher-orderinformation such as position-insensitive visual representations and intermediate representations of different grain sizes [73]. This approach allows empirical investigation of whetherparticularintermediaterepresentationsassumed tomediatebetweenvisualinputsandhigher-orderlexical outputs (e.g., bigrams, syllables) are in fact representa-tions that will emerge during learning, and how these representationsareconstrainedviafeedforwardand feed-backconstraints.
Taken together, biologically plausible connectionist modelsare showing strong initialpromisein beingable to provide detailed and explicit mechanistic accounts of ‘what’ and ‘where’ different typesof representations are stored[72,73],‘when’theyinteract,and‘how’the proces-singmechanicsofneurobiologicallyconstrained computa-tionsoperatethewaytheydo.Moreover,thesemodelscan beusednotonlyas‘process’modelsofthetypicalmature systembutcanalsotracethedevelopmentoflexicalskills andtop-downversusbottom-upinteractivity[74],andcan seamlesslyfacilitatestudies ofdifferenttypesof reading impairments[61].
Concludingremarks
Olddebatesregardingthemodularity–orlackthereof–of visualwordrecognitionarebackagainwiththeadventof techniques such as fMRI, EEG, and MEG. However, rekindlingofthesedebatesviaresearchusingsuch tech-niquesoffersanopportunityforuniquetheoreticaladvance thatwasnotpossibleviabehavioralinvestigationsalone, andalsoenablestheinvestigationofimportantnew ques-tions (Box 4). No longer are researchers constrained to
‘blackbox’theorizingregardingtheinternalmechanicsof the brain that mediate between stimulus and response. Rather, these representations can now be monitored directly andused to motivate specific theoretical claims abouttheintermediateinternalrepresentationsand pro-cesses thatsubservevisual word recognition. Likenever before, itisthereforepossibleto achieve integrated the-oriesof‘what’,‘when’,‘where’,and‘how’visualwordsare represented andprocessedinthebrain.
Acknowledgments
We acknowledge support of M.C. by the European Research Council (ERC-2011-ADG-295362)andbytheSpanishMinistryofEconomyand Competitiveness CONSOLIDER-INGENIO2010 CSD2008-00048 and PSI2012-31448; ofB.C.A. bytheMarie Curieprogram (PCIG13-2013-618429); of R.F. by the NICHD (RO1 HD067364); of R.F. and M.C. (PO1HD01994);andofM.P.bytheSpanishMinistryofEconomyand Competitiveness(PSI2011-26924).WethankS.Laszlofordiscussionof themodelingsection.
References
1 Rogers,T.T. etal.(2004)Structure anddeteriorationof semantic memory: a neuropsychological and computational investigation.
Psychol.Rev.111,205–235
2 Plaut, D.C. (2002) Graded modality-specific specialisation in semantics: a computational account of optic aphasia. Cogn.
Neuropsychol.19,603–639
3 Plaut, D.C. and Behrmann, M. (2011) Complementary neural representationsforfacesandwords:a computationalexploration.
Cogn.Neuropsychol.28,251–275
4 Price,C.J.(2012)Areviewandsynthesisofthefirst20yearsofPET andfMRIstudiesof heardspeech,spokenlanguageandreading.
Neuroimage62,816–847
5 Cohen,L.etal.(2000)Thevisualwordformarea:spatialandtemporal characterizationofaninitialstageofreadinginnormalsubjectsand posteriorsplit-brainpatients.Brain123,291–307
6 Cohen,L. etal.(2002) Language-specific tuningof visual cortex? Functional properties of the visual word form area. Brain 125, 1054–1069
7 Baker, C.I. etal.(2007) Visual word processing and experiential originsoffunctionalselectivityinhumanextrastriatecortex.Proc.
Natl.Acad.Sci.U.S.A.104,9087–9092
8 Dehaene,S.etal.(2002) Thevisual wordformarea: aprelexical representationofvisualwordsinthefusiformgyrus.Neuroreport13, 321–325
9 Glezer,L.S.etal.(2009)Evidenceforhighlyselectiveneuronaltuning towholewordsinthevisualwordformarea.Neuron62,199–204 10Binder,J.R.etal.(2006)Tuningofthehumanleftfusiformgyrusto
sublexicalorthographicstructure.Neuroimage33,739–748 11Dehaene,S.etal.(2005)Theneuralcodeforwrittenwords:aproposal.
TrendsCogn.Sci.9,335–341
12Dehaene,S.andCohen,L.(2011)Theuniqueroleofthevisualword formareainreading.TrendsCogn.Sci.15,254–262
13Price,C.J.andDevlin,J.T.(2003)Themythofthevisualwordform area.Neuroimage19,473–481
14Price,C.J.andDevlin,J.T.(2011)Theinteractiveaccountofventral occipitotemporalcontributionstoreading.TrendsCogn.Sci.15,246– 253
15Simos,P.G.etal.(2002)Brainmechanismsforreadingwordsand pseudowords:anintegratedapproach.Cereb.Cortex12,297–305 16Solomyak, O. and Marantz, A. (2010) Evidence for early
morphologicaldecomposition in visual wordrecognition. J.Cogn.
Neurosci.22,2042–2057
17Kronbichler,M. etal.(2004) The visualword form areaand the frequency with which words are encountered: evidence from a parametricfMRIstudy.Neuroimage21,946–953
18Kherif, F. et al. (2011) Automatic top-down processing explains common left occipito-temporal responses to visual words and objects.Cereb.Cortex21,103–114
19Bar,M.etal.(2006)Top-downfacilitationofvisualrecognition.Proc.
Natl.Acad.Sci.U.S.A.103,449–454
Box4.Outstandingquestionsandfuturedirections
How does anatomical and functional connectivity, and conse-quentlythetemporalflowofinformation,evolvefrompreliterate toskilledreading?
What are thefunctionaland anatomical differencesunderlying readingdisabilities,andhowcantheirunderstandingcanhelpthe implementationofremediationprograms?
How are connectivity and interactivity modulated by different languages having different scripts, different orthography-to-phonologyrelations,anddifferentmorphologicalsystems? What do the constraints of possessing different orthographic,
phonological,andsemanticrepresentationshaveonvisualword recognitioninthecaseofbilingualandmultilingualreaders? How can more biologically plausible computational models
interact with empirical investigations to produce theories that aremechanisticallyexplicit,comprehensive,andparsimonious? To what extent will a neurobiological theory of visual word
recognitionthatconsidersthefullpatternsofbrainconnectivity and interactivity provideinsightsinto domain-general mechan-ismssharedbyotherrelatedabilitiessuchasobjectrecognition?
20 Bar,M.(2007)Theproactivebrain:usinganalogiesandassociations togeneratepredictions.TrendsCogn.Sci.11,280–289
21 Kveraga,K.etal.(2007)Magnocellularprojectionsasthetriggerof top-downfacilitationinrecognition.J.Neurosci.27,13232–13240 22 Gazzaley,A.etal.(2007)Functionalinteractionsbetweenprefrontal
andvisualassociationcortexcontributetotop-downmodulationof visualprocessing.Cereb.Cortex17(Suppl.1),i125–i135
23 Zanto,T.P.etal.(2011)Causalroleoftheprefrontalcortexin top-downmodulationofvisual processing andworkingmemory.Nat.
Neurosci.14,656–661
24 Bar, M. (2003) A cortical mechanism for triggering top-down facilitationinvisualobjectrecognition.J.Cogn.Neurosci.15,600–609 25 Logothetis,N.K.(2008)Whatwecandoandwhatwecannotdowith
fMRI.Nature453,869–878
26 Ha¨ma¨la¨inen, M. et al.(1993) Magnetoencephalography – theory, instrumentation, and applications to noninvasive studies of the workinghumanbrain.Rev.Mod.Phys.65,413
27 Dunabeitia, J.A. et al. (2012) Differential sensitivity of letters, numbers, and symbols to character transpositions. J. Cogn.
Neurosci.24,1610–1624
28 Bentin,S.etal.(1999) ERPmanifestations ofprocessing printed words at different psycholinguistic levels: time course and scalp distribution.J.Cogn.Neurosci.11,235–260
29 Maurer,U.etal.(2005) Fast,visual specializationforreading in EnglishrevealedbythetopographyoftheN170ERPresponse.Behav.
BrainFunct.1,13
30 Allison,T.etal.(1994)Humanextrastriatevisualcortexandthe perceptionoffaces,words,numbers,andcolors.Cereb.Cortex4,544– 554
31 Nobre,A.C.etal.(1994) Wordrecognition inthe human inferior temporallobe.Nature372,260–263
32 Tarkiainen,A.etal.(1999)Dynamicsofletterstringperceptioninthe humanoccipitotemporalcortex.Brain122,2119–2132
33 Tarkiainen,A.etal.(2002)Dynamicsofvisualfeatureanalysisand object-levelprocessinginfaceversusletter-stringperception.Brain 125,1125–1136
34 Helenius,P.etal.(1999)Dissociationofnormalfeatureanalysisand deficientprocessingofletter-stringsindyslexicadults.Cereb.Cortex 9,476–483
35 Simos,P.G. etal.(2002) Dyslexia-specificbrain activationprofile becomesnormalfollowingsuccessful remedialtraining.Neurology 58,1203–1213
36 Simos,P.G.etal.(2005) Earlydevelopmentofneurophysiological processes involved in normal reading and reading disability: a magneticsourceimagingstudy.Neuropsychology19,787–798 37 Simos,P.G.etal.(2007)Alteringthebraincircuitsforreadingthrough
intervention:amagneticsourceimagingstudy.Neuropsychology21, 485–496
38 Brem,S.etal.(2010)Brainsensitivitytoprintemergeswhenchildren learn letter-speech sound correspondences.Proc. Natl. Acad. Sci.
U.S.A.107,7939–7944
39 Rauschecker,A.M.etal.(2012)Positionsensitivityinthevisualword formarea.Proc.Natl.Acad.Sci.U.S.A.109,E1568–E1577 40 Szwed, M. et al. (2012) Towards a universal neurobiological
architectureforlearningtoread.Behav.BrainSci.35,308 41 Assadollahi,R.andPulvermuller,F.(2003)Earlyinfluencesofword
lengthandfrequency:agroupstudyusingMEG. Neuroreport14, 1183–1187
42 Carreiras,M.etal.(2005)Earlyevent-relatedpotentialeffectsof syllabicprocessingduringvisualwordrecognition.J.Cogn.Neurosci. 17,1803–1817
43 Dambacher,M.etal.(2006)Frequencyandpredictabilityeffectson event-relatedpotentialsduringreading.BrainRes.1084,89–103 44 Hauk,O.etal.(2006)Thetimecourseofvisualwordrecognitionas
revealedbylinearregressionanalysisofERPdata.Neuroimage30, 1383–1400
45 Sereno, S. (2003) Measuring word recognition in reading: eye movementsandevent-relatedpotentials.TrendsCogn.Sci.7,489–493 46 Carreiras, M. et al. (2009) Consonants and vowels contribute differently to visual word recognition: ERPs of relative position priming.Cereb.Cortex19,2659–2670
47 Holcomb, P.J. and Grainger, J. (2007) Exploring the temporal dynamics of visual word recognition in the masked repetition
primingparadigmusingevent-relatedpotentials.BrainRes.1180, 39–58
48Dun˜abeitia,J.A.etal.(2009)N250effectsforlettertranspositions dependonlexicality:‘casual’or‘causal’?Neuroreport20,381–387 49Massol,S.etal.(2011)Whenlessismore:feedback,priming,andthe
pseudowordsuperiorityeffect.BrainRes.1386,153–164
50Carreiras, M. etal. (2009) Are vowels andconsonants processed differently?Event-relatedpotentialevidencewithadelayedletter paradigm.J.Cogn.Neurosci.21,275–288
51Carreiras, M. et al. (2007) ERP correlates of transposed-letter similarity effects: are consonants processed differently from vowels?Neurosci.Lett.419,219–224
52Carreiras,M.etal.(2009)ERPcorrelatesoftransposed-letterpriming effects:theroleofvowelsversusconsonants.Psychophysiology46,34– 42
53Dunabeitia, J.A. and Carreiras, M. (2011) The relative position priming effect depends on whether letters are vowels or consonants.J.Exp.Psychol.Learn.Mem.Cogn.37,1143–1163 54Thesen, T. etal. (2012)Sequential theninteractiveprocessing of
lettersandwordsintheleftfusiformgyrus.Nat.Commun.3,1284 55Cornelissen,P.L.etal.(2009)Activationoftheleftinferiorfrontal gyrus in the first 200ms of reading: evidence from magnetoencephalography(MEG).PLoSONE4,e5359
56Pammer,K.etal.(2004)Visualwordrecognition:thefirsthalfsecond.
Neuroimage22,1819–1825
57Wheat,K.L.etal.(2010)Duringvisualwordrecognition,phonologyis accessedwithin100msandmaybemediatedbyaspeechproduction code:evidencefrommagnetoencephalography.J.Neurosci.30,5229– 5233
58Woodhead,Z.V.et al. (2012)Readingfronttoback:MEGevidencefor earlyfeedbackeffectsduringwordrecognition.Cereb. Cortex http:// dx.doi.org/10.1093/cercor/bhs365
59McClelland, J.L. and Rumelhart, D.E. (1981) An interactive activation model of context effects in letter perception: I. An accountofbasicfindings.Psychol.Rev.88,375
60Seidenberg, M.S. and McClelland, J.L. (1989) A distributed, developmental model of word recognition and naming. Psychol. Rev.96,523–568
61Plaut,D.C.etal.(1996)Understandingnormalandimpairedword reading:computationalprinciplesinquasi-regulardomains.Psychol. Rev.103,56–115
62Reicher, G.M. (1969) Perceptual recognition as a function of meaningfulnessofstimulusmaterial.J.Exp.Psychol.81,275–280 63Ackley, D.H. et al. (1985) A learning algorithm for Boltzmann
machines.Cogn.Sci.9,147–169
64Davis, C.J. (2012) Developing a universal model of reading necessitatescracking the orthographic code.Behav. BrainSci. 1, 21–22
65Cohen,L.andDehaene,S.(2004)Specializationwithintheventral stream:thecaseforthevisualwordformarea.Neuroimage22,466– 476
66Borowsky, R. and Besner, D. (1993) Visual word recognition: a multistageactivation model. J.Exp. Psychol.Learn. Mem.Cogn. 19,813–840
67Borowsky,R.andBesner,D.(2006)Paralleldistributedprocessing andlexical-semantic effectsinvisual wordrecognition:are afew stagesnecessary?Psychol.Rev.113,181–195
68Plaut,D.C.andShallice,T.(1993)Deepdyslexia:acasestudyof connectionistneuropsychology.Cogn.Neuropsychol.10,377–500 69Laszlo, S. and Plaut, D.C. (2012) A neurally plausible parallel
distributed processing model of event-related potential word readingdata.BrainLang.120,271–281
70Laszlo,S.andArmstrong,B.C.(2013)Applyingthedynamicsof post-synapticpotentialstoindividualunitsinsimulationoftemporally extended ERP reading data. In Proceedings of the 35th Annual
ConferenceoftheCognitiveScienceSociety(Knauff,M.etal.,eds),
pp.2826–2831,CognitiveScienceSociety
71Rugg,M.D.(1985)Theeffectsofsemanticprimingandworkrepetition onevent-relatedpotentials.Psychophysiology22,642–647 72Hinton, G.E. and Salakhutdinov, R.R. (2006) Reducing the
dimensionalityofdatawithneuralnetworks.Science313,504–507 73Di Bono, M.G. and Zorzi, M. (2013) Deepgenerative learningof
location-invariantvisualwordrecognition.Front.Psychol.4,635
74 Harm, M.W. and Seidenberg, M.S. (1999) Phonology, reading acquisition, and dyslexia: insights from connectionist models.
Psychol.Rev.106,491–528
75 Forster,K.I.andDavis,C.(1984)Repetitionprimingandfrequency attenuationinlexicalaccess.J.Exp.Psychol.Learn.Mem.Cogn.10,680 76 Ferrand, L. and Grainger, J. (1994) Effects of orthography are independent of phonology in masked form priming. Q. J. Exp.
Psychol.47,365–382
77 Tzur,B.andFrost,R.(2007)SOAdoesnotrevealtheabsolutetime courseofcognitiveprocessinginfastprimingexperiments.J.Mem.
Lang.56,321–335
78 Rayner, K. (1998) Eye movements in reading and information processing:20yearsofresearch.Psychol.Bull.124,372
79 Be´langer,N.N.etal.(2013)Orthographicandphonologicalpreview benefits: parafoveal processing in skilled and less-skilled deaf readers.Q.J.Exp.Psychol.66,2237–2252
80 Frost,R.(1998)Towardastrongphonologicaltheoryofvisualword recognition:trueissuesandfalsetrails.Psychol.Bull.123,71 81 Rastle,K.andBrysbaert,M.(2006)Maskedphonologicalpriming
effectsinEnglish:aretheyreal?Dotheymatter?Cogn.Psychol.53, 97–145
82 Deutsch, A. etal. (2003) Early morphological effects in reading: evidencefromparafovealpreviewbenefitinHebrew.Psychon.Bull. Rev.10,415–422
83 Deutsch,A.etal.(2005)Morphologicalparafovealpreviewbenefiteffects inreading:evidencefromHebrew.Lang.Cogn.Process.20,341–371 84 Dunabeitia,J.A.etal.(2009)Thereisnoclamwithcoatsinthecalm
coast: delimiting the transposed-letter priming effect. Q. J. Exp.
Psychol.62,1930–1947
85 Frost, R. (2012) A universal approach to modeling visual word recognition and reading: not only possible, but also inevitable.
Behav.BrainSci.35,310–329
86 Norris,D.etal.(2000)Merginginformationinspeechrecognition: feedbackisnevernecessary.Behav.BrainSci.23,299–325 87 Grainger,J.etal.(2012)Orthographicprocessinginbaboons(Papio
papio).Science336,245–248
88 Ziegler,J.C.etal.(2013)What can welearn fromhumansabout orthographicprocessinginmonkeys?AreplytoFrostandKeuleers (2013).Psychol.Sci.24,1870–1871
89 Frost,R.andKeuleers,E.(2013)Whatcanwelearnfrommonkeys aboutorthographicprocessinginhumans?AReplytoZiegleretal..
Psychol.Sci.22,1868–1869
90 Andrews, S. (2006) All about words: a lexicalist perspective on reading. In From Ink Marks to Ideas: Current Issues in Lexical
Processing(Andrews,S.,ed.),pp.314–348,PsychologyPress
91 Nazir,T.A.etal.(2004)Reading habits,perceptuallearning,and recognitionofprintedwords.BrainLang.88,294–311
92 Frost,R.etal.(2012)Towardsauniversalmodelofreading.Behav.
BrainSci.35,263
93 Bitan, T. et al. (2007) Developmental changes in activation and effective connectivity in phonological processing. Neuroimage 38, 564–575
94Bitan, T. et al. (2005) Shifts of effective connectivity within a languagenetwork duringrhymingandspelling. J.Neurosci.25, 5397–5403
95Koyama,M.S.etal.(2010)Readingnetworksatrest.Cereb.Cortex20, 2549–2559
96Mechelli,A.etal.(2005)Dissociatingreadingprocessesonthebasisof neuronalinteractions.J.Cogn.Neurosci.17,1753–1765
97Wakana, S.etal. (2004)Fiber tract-based atlasof human white matteranatomy.Radiology230,77–87
98Barbas,H.(2000)Connectionsunderlyingthesynthesisofcognition, memory,andemotioninprimateprefrontalcortices.BrainRes.Bull. 52,319–330
99Bernal,B.andAltman,N.(2010)Theconnectivity ofthesuperior longitudinal fasciculus: a tractographyDTI study. Magn. Reson.
Imaging28,217–225
100 Kringelbach, M.L. (2004) and E. T. Rolls, The functional neuroanatomy of the human orbitofrontal cortex: evidence from neuroimagingandneuropsychology.Prog.Neurobiol.72,341–372 101 Makris,N.etal.(2005)Segmentationofsubcomponentswithinthe
superiorlongitudinalfascicleinhumans:aquantitative,invivo, DT-MRIstudy.Cereb.Cortex15,854–869
102 Bu¨rgel,U.etal.(2006)Whitematterfibertractsofthehumanbrain: three-dimensional mapping at microscopic resolution, topography andintersubjectvariability.Neuroimage29,1092–1105
103 Yeterian,E. and Pandya,D. (2010) Fiber pathways and cortical connections of preoccipital areas in rhesus monkeys. J. Comp.
Neurol.518,3725–3751
104 Yeterian,E.H.etal.(2012)Thecorticalconnectivityoftheprefrontal cortexinthemonkeybrain.Cortex48,58–81
105 Yeatman,J.D.etal.(2013)Anatomyofthevisualwordformarea: adjacentcorticalcircuitsandlong-rangewhitematterconnections.
BrainLang.125,146–155
106 Carreiras, M. et al.(2009) An anatomicalsignature for literacy.
Nature461,983–986
107 Richardson, F.M. et al. (2011) Multiple routes from occipital to temporalcorticesduringreading.J.Neurosci.31,8239–8247 108 Rosazza,C.etal.(2009)Earlyinvolvementofdorsal andventral
pathwaysinvisualwordrecognition:anERPstudy.BrainRes.1272, 32–44
109 Zachariou, V. et al. (2014) Ventral and dorsal visual stream contributionstotheperceptionofobjectshapeandobjectlocation.
J.Cogn.Neurosci.26,189–209
110 ThiebautdeSchotten,M.etal.(2012)Learningtoreadimprovesthe structureofthearcuatefasciculus.Cereb.Cortex http://dx.doi.org/ 10.1093/cercor/bhs383
111 Vogel, A.C. etal. (2012) The putative visual word form area is functionally connected to the dorsal attention network. Cereb.
Cortex22,537–549
112 vanderMark,S.etal.(2011)Theleftoccipitotemporalsystemin reading:disruptionoffocalfMRIconnectivitytoleftinferiorfrontal and inferior parietal language areas in children with dyslexia.
Neuroimage54,2426–2436