INTRODUCTION
Vertebrate fate mapping studies have established that the telencephalon, the retinal anlage and the ventral hypothalamus derive from adjacent domains in the anterior neural plate (ANP) (Sanchez-Arrones et al., 2009; Wilson and Houart, 2004). These domains are specified during gastrulation under the influence of morphogens produced by signalling centres discretely localised to the adjacent mesendoderm and in the ANP itself (Hoch et al., 2009; Wilson and Houart, 2004). In response to the reception of morphogenetic signals, cells express specific transcription factors (TFs), which activate intrinsic cellular programmes. These, in turn, force progenitor cells to commit toward specific forebrain field identities. Many of the TFs expressed in the ANP, including Sox2, Pax6, Otx2, Rx3, Six3, Six6, Foxg1 and Emx1, have been proven essential for the proper development of each of the forebrain regions in which they are expressed (England et al., 2006; Hoch et al., 2009; Shimamura and Rubenstein, 1997; Stern, 2002; Wilson and Houart, 2004). For some, for example Emx1 (Simeone et al., 1992), the contribution to prosencephalic patterning is easily explained by restricted expression to a specific subdomain. Others, including Otx2,Pax6and Six3, are more widely expressed in the forebrain (Bovolenta et al., 1998; Manuel and Price, 2005; Oliver et al., 1995; Simeone et al., 1992) or, like Sox2(Zappone et al., 2000), even throughout the neural tube, raising the as yet unanswered question of how their activity contributes to the identity of each particular forebrain domain in which they are expressed.
Hints to help unravel this problem come from a variety of sources. Characterisation of the regulatory regions governing Sox2, Otx2, Pax6and Six3expression indicates that their activity might be subject to regional and temporal-specific controls (Conte and Bovolenta, 2007; Kurokawa et al., 2004; Morgan, 2004; Uchikawa et al., 2003). Furthermore, Otx2 and Six3, for example, act both as transcriptional activators and repressors, interacting with other co-factors (Heimbucher et al., 2007; Kelley et al., 2000; Kobayashi et al., 2001; Lopez-Rios et al., 2003; Zhu et al., 2002), which might help in specialising their regional functions. Another relevant element of diversification is the level of gene expression. Heterozygous loss-of-function mutations in the human SOX2 or OTX2 genes can cause independent eye (Ragge et al., 2005a; Ragge et al., 2005b) or hypothalamic (Dateki et al., 2010; Tziaferi et al., 2008) defects without affecting other forebrain regions, which suggests a differential sensitivity of forebrain territories to gene dosage. Whether and how these different factors cooperate during normal development to adapt individual gene activity to the specific needs of the different forebrain regions is still unexplored.
The homeobox TF Six3 is a suitable candidate to address the above questions. Its expression is evolutionary conserved in the anterior forebrain (Bovolenta et al., 1998; Loosli et al., 1998; Lopez-Rios et al., 1999; Oliver et al., 1995; Seo et al., 1999), where it is required for the development of the telencephalon (Kobayashi et al., 1998; Lagutin et al., 2003), the eye (Carl et al., 2002; Loosli et al., 1999; Zhu et al., 2002; Zuber et al., 2003) and the hypothalamus (Gaston-Massuet et al., 2008; Jeong et al., 2008; Lavado et al., 2008; Ohyama et al., 2005). Still, it is unclear how it mediates regional differences. Among the vertebrate orthologues, Six3.2, one of the two medaka fish (Oryzias latipes) paralogues, offers particular advantages because its precise cis-regulatory code has been well characterised (Conte and Bovolenta, 2007), facilitating the identification of region-specific trans-acting factors. Furthermore, in teleosts, specific mRNA levels can be easily experimentally modulated (Carl et al., 2002).
Development 139, 151-164 (2012) doi:10.1242/dev.067660 © 2012. Published by The Company of Biologists Ltd
1Centro de Biología Molecular ‘Severo Ochoa’, CSIC-UAM, c/Nicolas Cabrera 1,
Madrid 28049, Spain. 2CIBER de Enfermedades Raras (CIBERER), c/ Nicolas Cabrera
1, Madrid 28049, Spain. 3Instituto Cajal, CSIC, Avda. Doctor Arce 37, 28002
Madrid, Spain.
*These authors contributed equally to this work
‡Present address: Telethon Institute of Genetics and Medicine (TIGEM) and Institute
of Genetics and Biophysics ‘A. Buzzati-Traverso’, CNR, via Pietro Castellino 111, 80131 Naples, Italy
§Author for correspondence ([email protected])
Accepted 28 October 2011 SUMMARY
The vertebrate forebrain is patterned during gastrulation into telencephalic, retinal, hypothalamic and diencephalic primordia. Specification of each of these domains requires the concerted activity of combinations of transcription factors (TFs). Paradoxically, some of these factors are widely expressed in the forebrain, which raises the question of how they can mediate regional
differences. To address this issue, we focused on the homeobox TF Six3.2. With genomic and functional approaches we
demonstrate that, in medaka fish, Six3.2 regulates, in a concentration-dependent manner, telencephalic and retinal specification under the direct control of Sox2.Six3.2and Sox2have antagonistic functions in hypothalamic development. These activities are, in part, executed by Foxg1and Rx3, which seem to be differentially and directly regulated by Six3.2 andSox2. Together, these data delineate the mechanisms by which Six3.2diversifies its activity in the forebrain and highlight a novel function for Sox2 as one of the main regulators of anterior forebrain development. They also demonstrate that graded levels of the same TF, probably operating in partially independent transcriptional networks, pattern the vertebrate forebrain along the anterior-posterior axis.
KEY WORDS: Gene networks, Hypothalamus, Forebrain patterning, Telencephalon, Transcriptional regulation, Medaka
Sox2-mediated differential activation of Six3.2
contributes to
forebrain patterning
Leonardo Beccari1,2,3, Ivan Conte1,*,‡, Elsa Cisneros1,2,3,* and Paola Bovolenta1,2,3,§
D
E
V
E
LO
P
M
E
N
Exploiting these advantages, here we delineate a gene regulatory network in which Six3.2diversifies its activity in the forebrain. We demonstrate that differential expression levels of Six3.2, operating under the differential control of Sox2, are likely to pattern the forebrain by directly activating the telencephalic determinant Foxg1 (Tao and Lai, 1992), restraining, at the same time, the expression of the hypothalamic and retinal determinant Rx3 (Deschet et al., 1999; Loosli et al., 2001; Stigloher et al., 2006). We also show that Sox2, besides controlling Six3.2 expression, activates that of Rx3, thus establishing a four gene ‘kernel’ that contributes to forebrain specification.
MATERIALS AND METHODS Medaka stocks
Wild-type (wt) Oryzias latipes of thecabstrain were maintained in an in-house facility (28°C on a 14/10-hour light/dark cycle). Embryos were staged according to Iwamatsu (Iwamatsu, 2004).
In silico analysis
Jaspar (http://jaspar.genereg.net) and rVista (http://rvista.dcode.org/) were used to identify putative TF binding sites (BSs) within the Six3.2regulatory region, setting the core identity to ≥90%. Positive sites were further screened for evolutionary conservation by phylogenetic footprinting among different teleost species (Fugu rubripes,Gasteropus aculeatus,Tetraodon nigroviridis andDanio rerio) using mVista (http://genome.lbl.gov/vista/ mvista/about.shtml) and multalin(http://multalin.toulouse.inra.fr/multalin). Identified candidates were further selected using expression pattern information available in the literature and in the ZFIN database (http://zfin.org).
Plasmid construction
The thymidine kinase (TK) minimal promoter was cloned into the pGL3-basic vector (Promega) to generate pGL3bTK, into which was then inserted PCR-amplified combinations (supplementary material Table S1) of the characterised Six3.2 regulatory elements (Conte and Bovolenta, 2007). Final constructs were: cI, comprising the entire Six3.2 regulatory region; cII, lacking the ABC elements; cIII, cIV and cV, respectively containing the D and IL elements or the TK minimal promoter alone (Fig. 1). Deletion constructs of the D element (cVI-cXI) were generated by PCR amplification from cIII using specific primers (supplementary material Table S1). Mutagenesis of the putative Sox BS was performed using the QuikChange Multi Site-Directed Mutagenesis Kit (Stratagene) using specific primers (cXII; supplementary material Table S1) and following the manufacturer’s instructions. The pd51LucII vector carrying the characterised 258 bp Nestinenhancer (Tanaka et al., 2004) was a kind gift of S. Nicolis (University of Bicocca, Milan). The 3.5 kb Rx3 regulatory region and full-length medaka Sox2,Six3.1and Six3.2were amplified by PCR and cloned into the pGL3b and pCMV-myc vectors (Clontech). For in vitro RNA transcription, myc-Sox2,myc-Six3.1and myc-Six3.2were subcloned into the pCS2+ vector.
RNA extraction and quantitative (Q) PCR
Total RNA from stage (st) 19 injected embryos was extracted using Trizol (Invitrogen) and treated with DNase I (Roche). cDNAs were obtained using the First-Strand cDNA Synthesis Kit (GE Healthcare). RT-PCR reactions were run in triplicate in 96-well plates with the SYBR Green PCR Master Mix using an ABI PRISM 7500 Sequence Detection System (Applied Biosystems). Q-RT-PCR reactions were performed with 2 ml cDNA, which was diluted tenfold for 18S rRNA amplifications used for normalisations. Primers specific for each of the analysed targets (supplementary material Table S1) were designed using Primer3Plus (http://www.primer3plus.com/). Analysis of the Q-RT-PCR assays was performed using the 7500 system SDS software (v2.0.1, Applied Biosystems). The SYBR Green-double-stranded DNA complex signal was normalised to the passive reference dye (ROX) to correct for non-PCR-related well-to-well fluorescence fluctuations. Experiments were independently replicated at least three times.
Cell transfection and luciferase assay
BHK21 or P19 cells were co-transfected using the Fugene HD transfection reagent (Roche) in MEMa-Glutamax medium (Gibco) and cultured for 48 hours in the presence of 5% foetal calf serum. For luciferase assays, cells were transfected with the reporter (50 ng) and Renilla luciferase (10 ng) plasmids and different amounts (25-200 ng) of the effector plasmid. Experiments were performed in triplicate and replicated at least three times as described (Conte et al., 2010b). Data are presented as fold induction of the luciferase activity observed in the control vector normalised to 1. Basal activity of the cI-cV construct is illustrated in supplementary material Fig. S2C.
Chromatin immunoprecipitation (ChIP) assays
ChIPs were performed as described (Conte et al., 2010b), using P19 cells transfected with a 3:1 ratio of reporter:effector plasmids (10 mg total). Fold enrichment was expressed as the ratio of myc-Sox2 to IgG signal and calculated with the formula 2–(Ct), where CtCt
c-Myc –CtIgG.
RNA and morpholino antisense oligonucleotide (MO) injections
Sox2,Six3.1and Six3.2synthetic mRNAs were obtained and injected as described (Conte et al., 2010b), using concentrations ranging from 5-50 ng, 20-80 ng and 1.0-30 ng, respectively. Selected working concentrations were 20 ng (Sox2), 20-40 ng (Six3.1) and 1.0-10 ng (Six3.2). MOs (supplementary material Table S1) (Gene Tools) were designed against the 5⬘UTR of Sox2,
Six3.1(Carl et al., 2002) and Six3.2. A standard control MO (Gene Tools) or an MO carrying five mismatches was used as control (generically referred to as cMO). The specificity and efficiency of each MO were determined as described (Eisen and Smith, 2008; Ruiz et al., 2009). cMO had no effects at any of the tested concentrations (20-300 mM) and embryos were undistinguishable from wt. Sox2-MO, Six3.1-MO and Six3.2-MO were tested at concentrations ranging from 50-300, 20-80 and 12.5-120 mM, and the selected working concentrations were 200, 20-40 and 20-60 mM, respectively. None of the MOs had off-target effects (Robu et al., 2007) at these concentrations. MOs were injected into two-cell stage embryos. At least three independent experiments were performed for each marker and condition.
In situ hybridisation
Whole-mount in situ hybridisations were performed as described (Lopez-Rios et al., 2008). Antisense and sense riboprobes for the medaka Fgf8,
Rx3, Rx2, Six3.2, Fezl, Foxb1.2, Foxg1, Sox1, Sox2, Sox3, Sox19,Dmbx1,
Emx3,Nkx2.1,Nkx2.2 andArxwere used. Probes were generated from cDNA amplified from st16-24 embryos using specific primers (supplementary material Table S1) and cloned into the pDrive vector (Qiagen). mRNAs were detected with NBT/BCIP (purple), INT/BCIP (orange/dark red), BCIP (light blue) or Fast Red (fluorescent red) substrates (Roche).
Cell proliferation analysis
Embryos were immunostained with anti-phospho-histone H3 antibodies (Roche Diagnostics) as described (Ruiz et al., 2009). Immunopositive cells were counted with the aid of ImageJ software (NIH) and normalised to the total area of each territory.
Image analysis and quantification
Embryos were photographed with a Leica M205FA and analysed with ImageJ. In st 16-18 embryos, the telencephalic field was identified by the rostral expression of Foxg1, Fgf8and Fezl, whereas the hypothalamic and diencephalic fields were identified by expression of Foxb1.2/mariposa and by the caudal domain of Fezl. At st 19-21, the telencephalic, diencephalic, mesencephalic and retinal territories were identified using the expression domains of Fgf8, Emx3, Foxg1, Arx, Nkx2.1, Dmbx1and Rx3or Rx2as depicted in supplementary material Fig. S1. Mean ± s.e.m. or mode and percentiles were calculated and plotted using Prism software (GraphPad). Student’s t-test was applied for median comparison between treatments. The frequency distribution of the Six3.2overexpression phenotypes was calculated with Prism. Telencephalic and retinal domains were considered expanded when their size was above the 75 percentile of the frequency distribution observed in controls and reduced when below the 25 percentile. Binomial distribution or Fisher’s exact test was used to compare
proportions of phenotypes.
D
E
V
E
LO
P
M
E
N
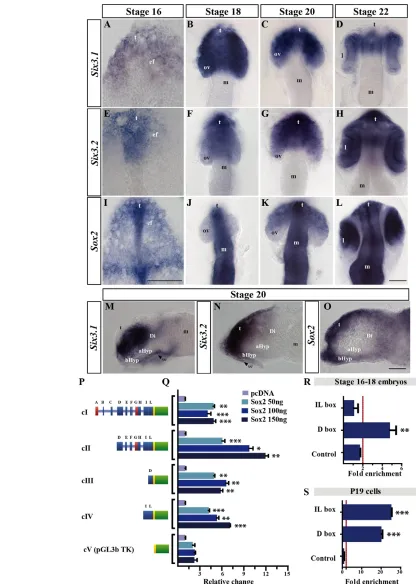
Fig. 1. Sox2is coexpressed with Six3.2in the forebrain and transactivates its regulatory region. (A-O)Dorsal (A-L) and lateral (M-O) views of medaka embryos hybridised with probes forSix3.1(A-D,M), Six3.2(E-H,N), Sox2(I-L,O). Eyes were removed in the lateral views. (P)TheSix3.2 promoter constructs (cI-cV) used in this study. Boxes represent silencers (red), enhancers or silencer blockers (blue), the basal TK promoter (yellow) and luciferase (green). Letters indicate identified HCNES. (Q) Luciferase assays performed with the cI-cV constructs. Bars represent mean + s.e.m. normalised to control. (R,S)ChIP of the predicted D and IL Sox binding sites (BSs) performed with anti-Sox2 on st 16-18 medaka embryos (R) and with anti-myc in P19 cells co-transfected with cII and myc-Sox2 (S). Mean + s.e.m. of a representative experiment performed in triplicate and repeated four times. Fold enrichment for each tested region was normalised to control IgG. Values above the red lines (R,S) were considered positive. Values above 3 were considered positive. *, P<0.01; **, P<0.001; ***, P<0.0001. aHyp, alar hypothalamus; bHyp, basal hypothalamus;
Di, diencephalon; ef, eye field; l, lens; m, mesencephalon; ov, optic vesicle; se, surface ectoderm; th, thalamus; t, telencephalon. Scale bars: 40mm.
D
E
V
E
LO
P
M
E
N
RESULTS
Identification of putative Six3.2transcriptional regulators
The medaka fish genome contains two Six3paralogues: Six3.1and Six3.2. The first is preponderantly expressed in the eye and the alar hypothalamic region (Fig. 1A-D,M; Fig. 2I,L) (Conte and Bovolenta, 2007; Loosli et al., 1998; Lopez-Rios et al., 2003), whereas the second is strongly expressed in the telencephalon and with lower intensity in the retinal and hypothalamic primordia (Conte and Bovolenta, 2007) (Fig. 1E-H,N; Fig. 2C,F). To investigate whether and how Six3.2mediates regional differences in the anterior forebrain, we sought to identify factors that control its graded expression, searching the previously characterised Six3.2 regulatory region (Fig. 1P) (Conte and Bovolenta, 2007) for the presence of evolutionary conserved TF BSs. Comparative searches in different databases (see Materials and methods) pinpointed a number of established TF BSs, which were exclusively clustered in the previously described highly conserved non-coding elements (HNCEs) present in the Six3.2 regulatory region (not shown). By integrating this information with available expression and functional data, members of the Sox, Pax, Otx, Tcf, Msx, Fox and TALE families were identified as potential transcriptional regulators (not shown). Among these, we focused on the SoxB subfamily. This was particularly attractive because verified Sox consensus matrices (Engelen et al., 2011; Kamachi et al., 2000)
were identified in both the D and IL HNCEs (supplementary material Fig. S2A,B). These two elements have been previously described to drive Six3.2expression in the ANP and prosencephalic structures at early (D) and late (IL) stages of development (Conte and Bovolenta, 2007), suggesting a possible relevant interaction between Sox and Six3.2activities.
Sox2 specifically binds the Six3.2regulatory region to control expression
SoxB genes are subdivided into SoxB1 (Sox1,2,3) and SoxB2 (Sox14,21) subgroups according to their preponderant activity as transcriptional activators or repressors (Chew and Gallo, 2009). Two additional SoxB1 paralogues, sox19a andsox19b, have been found in zebrafish (Okuda et al., 2010) (supplementary material Fig. S3A). As is the case with other TFs belonging to the same family, SoxB factors share a high degree of conservation in their DNA BS, suggesting a possible functional redundancy. Indeed, zebrafish sox2, sox3, sox19aand sox19bhave been shown to act redundantly at an early stage of neural specification (Okuda et al., 2010). We thus considered that the medaka orthologues of the SoxB1 subfamily could be potential regulators of Six3.2expression.
[image:4.612.50.350.339.739.2]Comparative in silico genome searches identified the medaka orthologues of all SoxB genes, although only one Sox19 paralogue was found (supplementary material Fig. S3A).
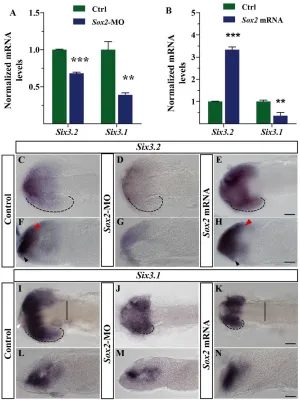
Fig. 2. Sox2 regulates Six3.2expression in vivo. (A,B)Six3.2and Six3.1mRNA levels (mean + s.e.m.) normalised to 18S rRNA in control medaka embryos and those injected with Sox2-MO orSox2 mRNA. **, P<0.001; ***, P<0.0001. (C-N)Dorsal (C-E,I-K) and lateral (F-H,L-N) views of st19 embryos treated as indicated and hybridised in toto with probes specific for Six3.1and Six3.2. Black (C-E,I-K) dashed lines indicate the extent of the retinal Six3.2or Six3.1expression domain. Red and black arrowheads in F,H indicate the telencephalic and hypothalamic Six3.2 expression domains, respectively. White arrowhead in I indicates the lack of Six3.1expression. Solid lines in I,K indicate the mesencephalic border. Scale bars: 40mm.
D
E
V
E
LO
P
M
E
N
Expression analysis revealed that Sox2 mRNA distribution coincided with that of Six3.2in the ANP from gastrulation stages (Fig. 1E,I), as described in other species (Papanayotou et al., 2008; Wood and Episkopou, 1999). Although broadly expressed throughout the entire neural tube, Sox2 was, as with Six3.2, differentially distributed in the forebrain, with high levels in the telencephalic domain (Fig. 1F,G,J,K,N,O; supplementary material Fig. S3C,G) and lower and graded (anteriorhigh to
posteriorlow) levels in the optic vesicles (Fig. 1F,G,J,K). Both
genes also overlapped in the basal hypothalamus (Fig. 1N,O), albeit with different intensities. Sox3 and Sox19 were also expressed in the forebrain but were virtually absent from the optic vesicles (supplementary material Fig. S3D,E,H,I), whereas Sox1 colocalised with Six3.2 only in the presumptive hypothalamic region at st20 (supplementary material Fig. S3B,F). According to this spatiotemporal expression, Sox2 appeared to be the most suitable candidate to activate Six3.2.
To corroborate this hypothesis, we compared the ability of Sox2 to transactivate luciferase reporter plasmids carrying different combinations of the Six3.2 regulatory elements (Fig. 1P). Co-transfection of increasing concentrations of Sox2in P19 (Fig. 1Q) or BHK21 (supplementary material Fig. S2D) cells transactivated those constructs carrying the D or IL HNCE. Furthermore, the cII construct, lacking the ABC HNCEs, was more efficiently activated by Sox2 than cI (Fig. 1Q), confirming the predicted existence of a silencer in the A element (Conte and Bovolenta, 2007). Notably, Sox3 and Sox1 showed only a lower or no transactivation activity on the cIII or cII constructs, when transfected into BHK21 or P19 cells (supplementary material Fig. S2D,E), although they activated, as efficiently as Sox2, a construct containing a characterised Nestin enhancer that responds to all SoxB1 genes (Tanaka et al., 2004). Similarly, Sox21, a SoxB2 representative, did not appear to modify
basal (supplementary material Fig. S2E,F) or Pax6-activated (not shown; see below) reporter expression from the cII or cIII constructs.
The D regulatory element contains clusters of six potential Sox BSs (S1-S6; supplementary material Fig. S2A,B), the sequence of which has recently been proven to functionally bind Sox2 in other contexts (Engelen et al., 2011). When deletion constructs (cVI-cXI) lacking different Sox BSs were probed with Sox2, reporter activation was virtually abolished in all constructs lacking the S2-S5 BS region (supplementary material Fig. S2G). Consistently, specific mutagenesis of S3-S5 had the same effect (supplementary material Fig. S2G), although the mutated enhancer responded efficiently to other characterised Six3.2activators such as Pax6 (data not shown), further supporting the conclusion that these BSs are specifically required for Sox2-mediated activation.
To verify whether Sox2 binds to the Six3.2 regulatory region in vivo, we performed ChIP assays using st16-18 medaka embryos. As expected for these stages, Sox2-specific antibodies revealed a fourfold enrichment of Sox2 on the D but not the IL element (Fig. 1R), which is active only at later stages. However, when P19 cells were co-transfected with the cII construct and a Sox2-myc-tagged expression plasmid, anti-myc antibody, but not control IgG, revealed a significant enrichment of Sox2 on both the D and IL enhancers (Fig. 1S), consistent with a report showing that Sox2 occupies the Six3promoter region in mouse embryonic stem cells (Boyer et al., 2005).
[image:5.612.55.521.429.680.2]According to the idea that Sox2 is a direct and physiological regulator ofSix3.2 expression, MO-mediated (200 mM) knockdown of Sox2caused a significant reduction of the total Six3.2 mRNA levels compared with control (cMO)-treated embryos (Fig. 2A). By in situ hybridisation, Six3.2 expression appeared strongly diminished throughout the prosencephalon (Fig. 2C,D,F,G).
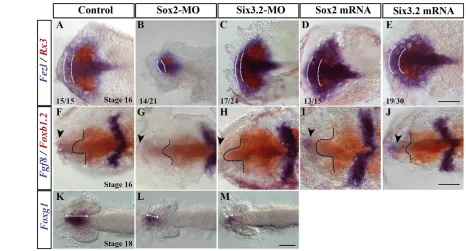
Fig. 3. Interference with Sox2and Six3.2expression alters anterior neural plate patterning. (A-M)Dorsal views of st16-18 medaka embryos treated as indicated and hybridised with probes forFezl and Rx3 (A-E), Fgf8and Foxb1.2(F-J) or Foxg1(K-M). The telencephalic domain of Fezland hypothalamic expression of Foxb1.2are outlined with white and black dashed lines. Black arrowheads in F-J mark the telencephalic expression of Fgf8. The extent the Foxg1-positive domain is indicated in K-M by double-headed arrows. The frequency of embryos with altered
forebrain patterning is indicated in A-E. Scale bars: 40mm.
D
E
V
E
LO
P
M
E
N
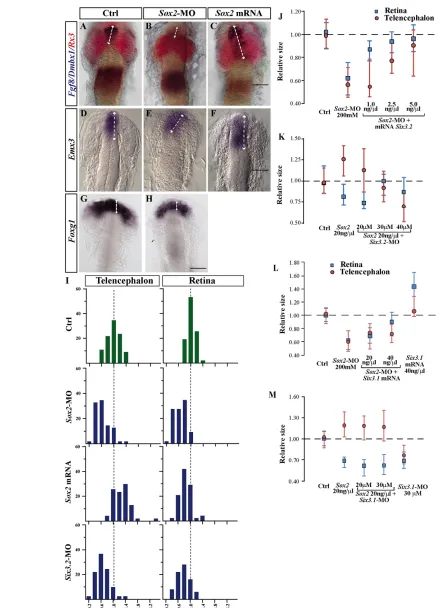
Fig. 4. Sox2and Six3.2favour telencephalic over retinal specification. (A-H)Dorsal views of medaka embryos injected as indicated and hybridised with probes for telencephalic (Emx3, Fgf8,Foxg1), retinal (Rx3) and mesencephalic (Dmbx1) markers. Controls were injected with GFPor cMO. Telencephalic marker expression is expanded (double-headed arrows) in C,F. (I) Frequency distribution of the telencephalic and retinal size of embryos injected as indicated. The size of the telencephalon and retina was quantified according to Fgf8and Rx3expression domains (see Materials and methods); values were normalised to those of controls. (J-M)Median representations of telencephalic (red circles) and retinal (blue squares) size in embryos treated as indicated. The whiskers represent the 75 and 25 percentile of the distribution. Scale bars: 40mm.
D
E
V
E
LO
P
M
E
N
Conversely, injections of Sox2, but not EGFP, mRNAs (20 ng/ml) significantly increased the levels of Six3.2transcripts (Fig. 2B), the localisation of which was caudally expanded (Fig. 2C,E,F,H). Sox2 morphants were also characterised by a significant decrease in the mRNA levels of the Six3.1paralogue (Fig. 2A), a reduction that was mostly evident in the subpallium, diencephalon and hypothalamus (Fig. 2I,J,L,M). However, Sox2 gain-of-function had a negative effect on total Six3.1mRNA levels (Fig. 2B), probably owing to the small size of the optic vesicles and altered expression in the diencephalon and hypothalamus (Fig. 2I,K,L,N). Because Sox2and Six3.1expression do not overlap in the latter regions at this stage (Fig. 1C,K,M,O), this effect was likely to be indirect.
Together, these data indicate that, in vivo, Sox2 predominantly activates the Six3.2medaka paralogue, but indirectly influences the domain of Six3.1expression. Next, we investigated the functional consequences of the direct Sox2-mediated regulation of Six3.2.
Telencephalic and retinal specification require different levels of Sox2/Six3.2activity
Patterning of the ANP begins at early gastrulation (Wilson and Houart, 2004). Consistent with an important role of Sox2 in establishing neural identity (Uchikawa et al., 2011), st16 Sox2 morphants displayed reduced telencephalic, retinal and hypothalamic fields, identified by the expression of Fgf8/Fezl/Foxg1, Rx3and Foxb1.2, respectively (Fig. 3). Notably, this reduction was more evident in the prospective telencephalic domain, where the expression of Fezl and Fgf8 was strongly reduced (Fig. 3A,B,F,G) and, in the case of Fgf8, also slightly delayed when compared with controls (not shown). Supporting that Six3.2is among Sox2 targets, Six3.2morphants showed a similar, although less evident, phenotype, with a pronounced reduction of the telencephalic versus retinal field but, surprisingly, an enlarged hypothalamic domain (Fig. 3C,H). Conversely, Sox2 or Six3.2 overexpression expanded the telencephalic field but reduced the Rx3-positive domain (Fig. 3A,D-F,I,J). Furthermore, as observed in the respective morphants, Sox2and Six3.2 had opposite effects in the hypothalamus when overexpressed (Fig. 3F,I,J).
Similarities between the phenotypes induced by the manipulation of Sox2and Six3.2expression were more apparent at slightly later stages (st18-20), when Sox2and Six3.2morphants were characterised by significantly reduced retinal (Rx3or Rx2 positive) and telencephalic (Fgf8-, Emx3- or Foxg1-positive) primordia, when compared with cMO-injected embryos (Fig. 3K-M; Fig. 4A,B,D,E,G-J; supplementary material Table S2). Conversely, Sox2overexpression caused telencephalic enlargement and retinal reduction (Fig. 4C,F,I,K; supplementary material Table S2), whereas other caudal morphogenetic events were not significantly affected (not shown).
A reduction of the retinal primordia upon Sox2 or Six3.2 overexpression was somewhat unexpected because both genes are required for eye formation in mammals and Six3.1 expands the eye field and induces the formation of ectopic retina-like tissue (Lagutin et al., 2003; Loosli et al., 1999; Lopez-Rios et al., 2003; Taranova et al., 2006). We therefore hypothesized that Sox2-mediated regulation of Six3.2 must predominantly favour telencephalic development at the expense of the retina, overriding Six3.1activity in this tissue. If this were the case, Six3.2but not Six3.1should preferentially counteract the impaired telencephalic development observed in Sox2 morphants. Indeed, co-injection of Sox2-MO with increasing concentrations of Six3.2 mRNA progressively rescued telencephalic and retinal size (Fig. 4J; supplementary material Table S2), but the levels necessary to
rescue the retina were lower than those required for the telencephalon (Fig. 4J; supplementary material Table S2). Increasing Six3.1mRNA doses instead did not modify the size of the telencephalon in Sox2morphants but did partially rescue the retina (Fig. 4L; supplementary material Table S2). Similarly, when co-injected, Six3.2-MO but not Six3.1-MO counteracted the Sox2 mRNA-mediated alterations of the retinal and telencephalic domains in a concentration-dependent and region-specific manner (Fig. 4K,M; supplementary material Table S2): low Six3.2-MO doses (20 mM) partially rescued telencephalic enlargement but had no effect on the retina (Fig. 4K; supplementary material Table S2), whereas a large proportion of the embryos showed a wt phenotype when intermediate Six3.2-MO doses (30 mM) were used. Notably, high Six3.2-MO levels (40 mM) significantly reduced the average size of both the telencephalon and the retina below that of controls (Fig. 4K; supplementary material Table S2), indicating a likely decrease of Six3.2expression below wt thresholds. Consistently, Six3.2morphants (30-60 mM) showed a similar phenotype, with their average telencephalic reduction more prominent than that observed for the retina (Fig. 4I; supplementary material Table S2). Together, these data indicate that, in medaka, Six3.2 levels differentially affect the specification of the telencephalon and the retina. The first depends on high Six3.2activity, whereas retinal development is likely to rely on Six3.1function (Carl et al., 2002) but requires only low Six3.2activity.
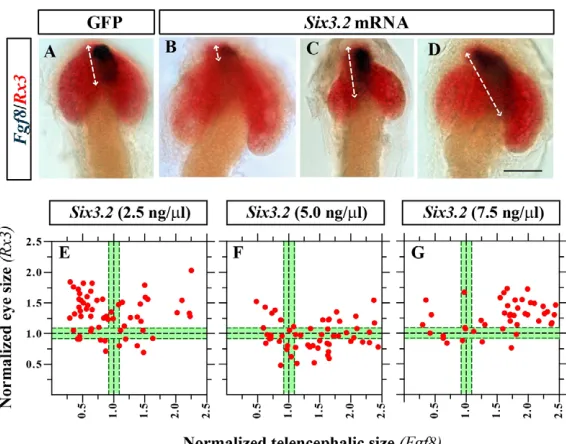
To corroborate this dose-dependent regulation of telencephalic and retinal development, we overexpressed different concentrations of Six3.2mRNA. When compared with GFP-injected controls, Six3.2 gain-of-function resulted in different phenotypes characterised by either reduced or enlarged optic vesicles (Fig. 5). Embryos with reduced Rx-positive territories presented a significant expansion of the Fgf8-or Emx3-positive telencephalon (Fig. 5C; supplementary material Table S4), whereas embryos with enlarged Rx-positive tissue showed either reduced (Fig. 5B) or enlarged (Fig. 5D) telencephalic marker expression. Although a mixture of at least two phenotypes was always observed, the relative proportion of each phenotype differed significantly among embryos injected with distinct Six3.2mRNA doses (supplementary material Table S4). At low doses (2.5 ng/ml) the majority of the embryos showed enlarged retinas with a smaller telencephalon (Fig. 5B,E), whereas at intermediate doses (5 ng/ml) most embryos resembled those observed upon Sox2overexpression, with small optic vesicles and enlarged telencephalon (Fig. 5C,F). Surprisingly, at high Six3.2 mRNA levels (7.5 ng/ml) embryos with both telencephalic and retinal enlargement predominated (Fig. 5D,G). This phenotype resembles that observed upon Six3 or Six6 overexpression (Loosli et al., 1999; Lopez-Rios et al., 2003; Zuber et al., 2003), suggesting that at high doses Six3.2might acquire functions that are normally exerted by the related Six3.1and Six6.
Six3.2represses hypothalamic specification by a
Sox2-independent mechanism
Taken together, these results support a model in which Sox2 differentially activates Six3.2in the forebrain. Higher Six3.2 levels would promote telencephalic specification while limiting retinal marker expression, which, instead, would only require low Six3.2 doses.
Sox2and Six3.2were also co-expressed in the hypothalamus (Fig. 1N,O), where alteration of Sox2expression also appeared to modify Six3.2mRNA levels (Fig. 2F-H). Mutations in SOX2 cause hypothalamic defects in humans (Hagstrom et al., 2005; Kelberman et al., 2008) and in Six3–/– mouse embryos the
D
E
V
E
LO
P
M
E
N
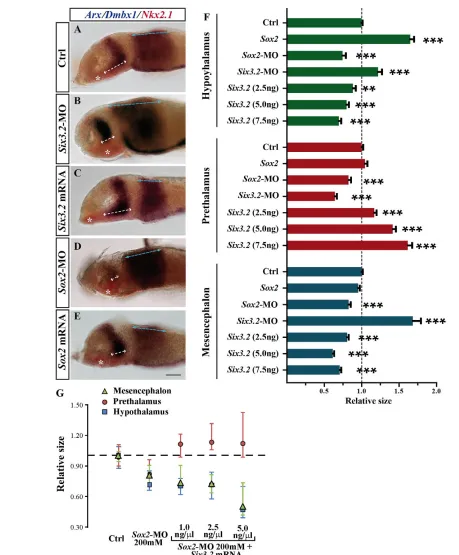
anterior forebrain, including the hypothalamus, is absent (Lavado et al., 2008). Furthermore, in medaka, moderate reduction of Six3.1activity interferes with ventral forebrain formation (Carl et al., 2002). By contrast, in st16 Sox2and Six3.2morphants, the Foxb1.2-positive hypothalamic domain suffered opposite alterations (Fig. 3F-H), suggesting that the effects of Six3.2in the hypothalamus are Sox2independent. In agreement with this possibility, st21 Six3.2 morphants showed a statistically significant increase of the Rx3-positive (not shown) and Nkx2.1-positive hypothalamic domain associated with a decrease in the Arx-positive prethalamic region (Fig. 6A,B,F,G). Conversely, injections of increasing concentrations of Six3.2 mRNA progressively reduced the hypothalamic and Dbmx1-positive mesencephalic domains but expanded the prethalamic region (Fig. 6A,B,C,F). Sox2overexpression instead caused a notable increase of the Nkx2.1-positive hypothalamus and had no effect on the diencephalic or mesencephalic domains (Fig. 6A,E,F), whereas in Sox2 morphants the hypothalamus, and to a much lower extent the prethalamus and the mesencephalon, were decreased (Fig. 6A,D,F,G; supplementary material Table S3). Furthermore, and in contrast to what was observed in the telencephalon, Six3.2mRNA (1.0 and 2.5 ng/ml) did not rescue the reduced hypothalamic and mesencephalic size of Sox2 morphants; rather, when compared with Sox2morphants, high Six3.2doses further reduced the Nkx2.1- and Dmbx1-positive territories and expanded the prethalamic domain (Fig. 6G).
These results suggest that Six3.2 normally antagonises hypothalamic development via a mechanism that seems independent from Sox2 function. Sox2, by contrast, promotes hypothalamic development by activating other target genes.
Increased telencephalic precursor proliferation and direct activation of Foxg1and Rx3may account forSox2and Six3.2activities in the anterior forebrain
Taken together, our results suggested that graded Six3.2 levels pattern the forebrain along the anterior-posterior axis, rostrally upon Sox2 control and caudally with a Sox2-independent mechanism. To explain these observations we took into consideration two non-mutually exclusive mechanisms.
First, differential mitotic rates could account for the observed forebrain alterations because both Sox2and Six3.2control neural precursor proliferation (Del Bene et al., 2004; Ferri et al., 2004; Gestri et al., 2005; Taranova et al., 2006). Evaluation of regional restricted proliferation using phospho-histone H3 immunostaining revealed that Sox2or Six3.2overexpression specifically promoted the proliferation of telencephalic precursors but, respectively, barely affected or reduced those of the retina. Their knockdown, instead, reduced proliferation in both domains when compared with controls (supplementary material Fig. S4). This suggests that Sox2-mediated activation of Six3.2is likely to recruit more proliferating progenitors to the telencephalic field.
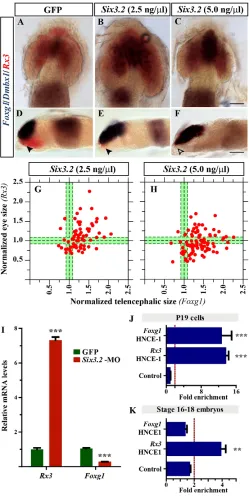
Second, and to corroborate the above possibility, we considered that Six3.2 could impose tissue identity by controlling the expression of other anterior forebrain determinants. Recent studies in zebrafish showed that rx3promotes retinal specification at the expense of the telencephalon, repressing, directly or indirectly, foxg1activity (Stigloher et al., 2006). Notably, increasing Six3.2 mRNA concentrations significantly reduced and totally abolished Rx3expression in the retina and hypothalamus, respectively (Fig. 7A-H; supplementary material Table S4), even at concentrations at which other hypothalamic markers, including Nkx2.1, were still observed (Fig. 6A,C). Conversely, Six3.2 overexpression significantly expanded Foxg1expression even at low doses (Fig. 7A-H; supplementary material Table S4), but had milder effects on the expansion of other telencephalic markers such as Fgf8and Emx3 (Fig. 5E,F; supplementary material Table S4).Consistently, the levels of Rx3 mRNA were upregulated fourfold in Six3.2 morphants, whereas those of Foxg1were threefold lower than those detected in GFP-injected embryos (Fig. 7I).
[image:8.612.53.336.58.280.2]Thus, Six3.2appeared to counteract the activity reported for Rx3 (Stigloher et al., 2006), and its marked effects on Rx3and Foxg1 expression pointed to a possible direct regulation. Phylogenetic footprinting of the vertebrate Foxg1and Rx3loci identified HNCEs containing putative conserved Six3 BSs (supplementary material Fig. S5). ChIP assays performed in P19 cells confirmed that transfected Six3.2-mycbound with high enrichment to HNCE-1 of the co-transfected Rx3medaka promoter and to HNCE-1 of the endogenous Foxg1regulatory region (Fig. 7J). Previous studies have shown that Sox2 binds to, and transactivates, an enhancer
Fig. 5. Six3.2gain-of-function differentially affects telencephalic and retinal development. (A-D)Dorsal views of medaka embryos hybridised with probes specific for Fgf8(blue) and Rx3(red) and injected with GFP(A) or increasing doses of Six3.2mRNA (B-D). Six3.2
overexpression causes retinal reduction (C) or expansion (B,D). The extension of the telencephalic domain is marked by double-headed arrows. (E-G)Plot representation of eye (y) versus telencephalic (x) size in embryos injected with Six3.2mRNA (2.5-7.5 ng/ml). Sizes were determined using the expression domains of Rx3and Fgf8, respectively. The green area represents the size of GFP-injected embryos and the black dashed lines indicate the mean and the 25 and 75 percentiles of the distribution. Scale bar: 40mm
D
E
V
E
LO
P
M
E
N
present in the Xenopus Rx1locus (Danno et al., 2008). The medaka Rx3 HNCE-1 also contains a Sox BS (supplementary material Fig. S5C) to which Sox2 bound with fourfold enrichment in st16-18
[image:9.612.55.514.56.611.2]medaka embryos (Fig. 7K), offering a possible explanation for the Sox2-mediated expansion of the hypothalamic domain (Fig. 6A,E-G).
Fig. 6. Sox2and Six3.2have opposite roles in hypothalamic specification. (A-E)Lateral views of medaka embryos treated as indicated and hybridised withNkx2.1(hypothalamus), Arx(diencephalon) and Dmbx1(mesencephalon) probes. Eyes were removed for photography. Blue and white dashed lines indicate the extension of the mesencephalic and Arx-positive prethalamic domains, respectively. Asterisks mark the
hypothalamus. (F)Quantification of the hypothalamic, diencephalic and mesencephalic size of treated embryos. Bars represent mean + s.e.m. ***, P<0.0001; **, P<0.001. (G)Median representations of hypothalamic (blue squares), prethalamic (red circles) and mesencephalic (green triangles) size of treated embryos. The whiskers represent the 75 and 25 percentiles of the distribution. Scale bar: 40mm.
D
E
V
E
LO
P
M
E
N
Together, these studies suggest a gene regulatory network in which Six3.2, together with Sox2, operates to pattern the vertebrate forebrain (Fig. 8). Disturbance of the gene network balance causes patterning alterations.
DISCUSSION
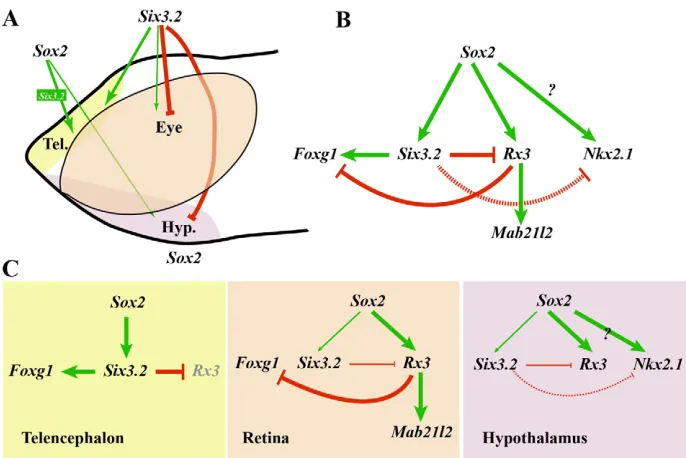
The identification of gene regulatory networks responsible for the specification of cell identity is important with a view to understanding how embryonic development is driven and provides fundamental information that can be exploited to reprogram stem cells for tissue repair. Yet disentangling cell-specific genetic programmes is a major challenge, especially for vertebrate tissues because master regulatory genes have often been co-opted to specify cells of related embryological origin. Combining genomic and functional approaches we show that different levels of Six3.2, acting both as a transcriptional activator and repressor, contribute to the specification of the distinct forebrain domains. In doing so, we have highlighted a novel role for Sox2in forebrain development and delineated a possible hierarchy of four TFs – Sox2, Six3.2, Foxg1and Rx3– that act to define the telencephalic, retinal and hypothalamic domains within the ANP (Fig. 8).
Specification and patterning of the anterior forebrain require sustained inhibition of Wnt signalling (Wilson and Houart, 2004). Soluble Wnt antagonists, the repressor activity of Tcf3 (a member of the Tcf/Lef family of TFs) and Six3 constrain Wnt canonical signalling, promote telencephalic and retinal fates and delimit the diencephalic territory. Abrogation of the function of any of these molecules increases canonical Wnt signalling, enlarges the diencephalon and impairs anterior forebrain formation (Houart et al., 2002; Kim et al., 2000; Lagutin et al., 2003). We provide compelling genomic and functional evidence that Sox2is an additional major regulator of anterior forebrain development by directly activating Six3.2. Although, Sox1 and Sox3 showed a lower transactivation activity than Sox2 or no transactivation activity – especially in P19 cells, in which appropriate Sox co-factors might exist and contribute to refine transactivation – on the Six3.2regulatory region, our study does not exclude the possibility that SoxB1 factors other than Sox2 might contribute to Six3.2regulation. Indeed, SoxB1 genes have been shown to act redundantly in other contexts (Okuda et al., 2010; Tanaka et al., 2004) and, in medaka, their partially overlapping distribution (supplementary material Fig. S3) implies a similar role in forebrain patterning. Nevertheless, specific distribution, differential affinity for the Sox BS and unique protein-protein interactions (Kamachi et al., 2000) may underlie the preponderant role of Sox2 in medaka anterior forebrain specification uncovered here. Consistent with this idea, SoxB1 genes have recently been shown to activate distinct repertoires of target genes during Xenopusneurogenesis (Archer et al., 2011).
[image:10.612.50.300.60.561.2]Sox2function has been extensively investigated because of its capacity to confer pluripotency to embryonic and reprogrammed stem cells (Avilion et al., 2003; Masui et al., 2007; Takahashi and Yamanaka, 2006), but the consequent early embryonic lethality of homozygous Sox2mutants (Avilion et al., 2003) has hampered the establishment of its function in the CNS. Conditional inactivation of Sox2in the embryonic mouse brain, after forebrain patterning has occurred, causes only minor defects at birth, although postnatal neurogenesis in the hippocampus is severely affected (Favaro et al., 2009). Our study reveals a novel and early aspect of Sox2function in the forebrain. Knockdown of Sox2 impairs telencephalic development and decreases Six3.2 expression, whereas Sox2 Fig. 7. Six3.2 and Sox2 differentially bind to Foxg1and Rx3
enhancers and regulate their expression. (A-F)Dorsal (A-C) and lateral (D-F) views of Foxg1,Dmbx1 and Rx3 hybridised embryos injected with GFPor Six3.2mRNA. Even low doses of Six3.2 significantly expand Foxg1expression and reduce (arrowheads in D,E) or abrogate (open arrowhead in F) hypothalamic Rx3expression. (G,H)Telencephalic versus eye size in embryos injected with Six3.2 mRNA. Tissue size was determined according to the Foxg1and Rx3 expression domains. (I)Foxg1and Rx3mRNA levels (mean + s.e.m.) normalised to 18S rRNA in control and Six3.2-MO-injected embryos. *** P<0.0001. (J,K)ChIP of the predicted Six and Sox BSs performed with anti-myc antibodies in P19 cells transfected with myc-tagged Six3.2(J) or anti-Sox2 in st16-18 medaka embryos (K). Histograms show the mean value of a representative experiment performed in triplicate and repeated three times. Fold enrichment was normalised to control IgG. Values above the red dotted line were considered positive.
***, P<0.0001; **, P<0.001. Scale bars: 40mm.
D
E
V
E
LO
P
M
E
N
overexpression has the opposite effects. These phenotypes are progressively rescued by the co-injection of increasing concentrations of Six3.2mRNA and MO, respectively, whereas similar experiments using the Six3.1paralog were less efficient, especially when the Six3.1-MO was used. These findings, together with luciferase and ChIP data, indicate that Six3.2is a major target of Sox2in the telencephalon and that telencephalic specification requires high Six3.2 activity.
Notably, modifications to Sox2 and Six3.2 expression also interfere with retinal development, but with notable differences to their effects on the telencephalon. First, a reduction in retinal size was evident with gain- and loss-of-function of both genes, similar to what has been reported for mice with high and low Pax6levels (Schedl et al., 1996), suggesting that, within the forebrain, the retina might be particularly sensitive to extreme gene dosage. Second, low Six3.2 mRNA concentrations were sufficient to normalise the retina of Sox2morphants. Third, whereas Six3.2 knockdown could easily counterbalance the effects of Sox2 overexpression, co-injections of Six3.1-MO had almost the opposite effect. Therefore, proper retinal formation requires lower Sox2 and Six3.2 levels than those necessary to pattern the telencephalon. This interpretation is supported by the mRNA distribution of Six3.2 and Sox2 in the forebrain, which is characterised by higher levels in the telencephalon than in the retina. Distinct dose-dependent Sox2functions are not unique to the forebrain. Trachea and oesophagus patterning is also Sox2 dependent, but Sox2hypomorphic embryos develop a trachea with oesophagus-like morphology, indicating that the trachea requires lower Sox2 levels (Que et al., 2007).
What determines these graded levels is unclear but the control might be post-transcriptional. MicroRNAs with retinal-restricted expression have been reported (Alfano et al., 2005; Conte et al., 2010a; Huang et al., 2008), pointing to this mechanism as an attractive possibility. Graded expression might be also a direct consequence of Sox2regulation. Sox2is expressed with the same graded intensity in the anterior forebrain and the D HNCE contains several Sox BSs, for which Sox2 might have different affinities, differentially contributing to full Six3.2activation. The activation of cis-regulatory modules by concentration gradients of TFs
depends upon the use of high- and low-affinity DNA BSs, where only high TF concentrations can activate the low-affinity sites (Bonn and Furlong, 2008; Papatsenko and Levine, 2005). It is thus conceivable that only high Sox2 levels in the anterior-most forebrain can fully activate the D enhancer, providing the required Six3.2activity for telencephalic development.
The telencephalic and retinal domains are initially closely related because manipulations of canonical Wnt signalling simultaneously affect both fields (Wilson and Houart, 2004). Activation of non-canonical Frizzled receptors and expression of Sfrp1 and Rx3 contribute to define the retinal territory, segregating it from the telencephalon in teleosts (Cavodeassi et al., 2005; Esteve et al., 2004; Lopez-Rios et al., 2008; Stigloher et al., 2006). Our study, showing that Sox2and Six3.2alterations always affect the telencephalon more than the retina, has two important implications.
[image:11.612.50.393.58.287.2]First, it suggests that the preponderant role of Sox2/Six3.2in medaka telencephalic development counteracts the activity of retinal and hypothalamic determinants such as Rx3 (Stigloher et al., 2006). Supporting this interpretation, we show that Six3.2 binds to the regulatory region and directly controls the expression of the telencephalic determinant Foxg1(Tao and Lai, 1992), providing experimental evidence to support previous suggestions in chick (Kobayashi et al., 2002). Six3.2 also binds to an Rx3enhancer but is likely to repress its expression, at least in the hypothalamus. Rx3, in turn, seems to antagonise the telencephalic determinants Foxg1 and Fez1 and to activate expression of the retinal determinant Mab21l2 (Kennedy et al., 2004; Stigloher et al., 2006). Therefore, we propose a regulatory network (Fig. 8) in which high levels of Six3.2 expression, under the direct control of Sox2, repress Rx3 expression and thereby favour that of Foxg1in the telencephalon. In the retina, however, a lower Six3.2 activity would not be sufficient to repress Rx3, the activation of which would consequently repress Foxg1-mediated telencephalic fate, as previously suggested (Kennedy et al., 2004; Stigloher et al., 2006). In support of this hypothesis, a recent study has shown that differences in the extent of the Six3expression domain positively correlate with the size of the telencephalon in cichlid fishes (Sylvester et al., 2010). Furthermore, a dual function of Six3.2 is
Fig. 8. Proposed gene regulatory network involved in anterior forebrain patterning. (A)The dose-dependent requirement of Sox2and Six3.2for telencephalic, retinal and hypothalamic patterning. High (thick arrows) and low (thin arrows) Sox2and Six3.2activity are required for telencephalic and retinal specification, respectively. Sox2and Six3.2 have antagonist functions in hypothalamic specification. (B)The gene regulatory network implicated in prosencephalic specification. (C)The operating mode of the proposed transcriptional network in each of the forebrain territories. Green and red arrows represent positive and negative regulations, respectively. Continuous lines indicate direct regulation, dashed lines indirect regulation. Grey symbols represent inactive or poorly expressed genes or inactive regulations. Question mark indicates possible direct regulations. Tel., telencephalon; Hyp., hypothalamus.
D
E
V
E
LO
P
M
E
N
compatible with studies of other Six3orthologues showing that activator and repressor roles depend on context-specific interactions. Indeed, Six3 binds a variety of proteins, including other TFs (Tessmar et al., 2002), transcriptional co-repressors of the Groucho family (Kobayashi et al., 2001; Lopez-Rios et al., 2003), coactivators such as EYA4 (Abe et al., 2009) and components of the chromatin remodelling complexes, including MTA1 and HDAC2 (Manavathi et al., 2007).
Second, our results show that the medaka Six3paralogues have undergone at least partial subfunctionalisation, with a major function for Six3.1in the eye and significant involvement of Six3.2 in telencephalon formation. Six3.2gain-of-function did not induce the formation of ectopic retinal tissue, in contrast to the effect of Six3.1 overexpression (Loosli et al., 1999; Lopez-Rios et al., 2003), but rather appeared to recruit proliferating progenitors into the telencephalic domain. Conversely, alterations of Six3.1expression had a clear influence on retinal development but only a marginal effect on that of the telencephalon (Carl et al., 2002; Loosli et al., 1999; Lopez-Rios et al., 2003). The two medaka Six3paralogues have a virtually identical homeobox sequence, which probably explains why high Six3.2concentrations cause a phenotype similar to that observed upon Six3.1overexpression (Fig. 5G,J). However, there are also appreciable divergences in the Six and C-terminal domains, which are known to mediate binding to various interactors (Kumar, 2009; Weasner and Kumar, 2009). This suggests that the two paralogues normally participate in different transcriptional complexes. Functional differences might also be linked to Sox2-mediated differential regulation, as Sox2 strongly activatesSix3.2but seems to reduce, directly or indirectly, Six3.1 expression (Fig. 2). Furthermore, Sox2 overexpression never expanded retinal size, which might reflect its opposing effects on Six3.2and Six3.1expression, as well as the influence of Sox2 on other, as yet undefined, genes implicated in telencephalic versus eye and hypothalamic fate acquisition.
Whether Six3.1and Six3.2exhibit reciprocal regulation, refining telencephalic and retinal differences, is unclear. Injections of Six3.2-MO appeared to lower Six3.1mRNA levels and vice versa, and Six3.2 activates the regulatory region of Six3.1in luciferase assays (L.B. and P.B., unpublished). The existence of this loop could explain why low doses of Six3.2reduce the telencephalon and expand eye size, possibly favouring the proposed Six3.1-mediated maintenance of Rx3 expression (Carl et al., 2002). Changes in relative retinal and telencephalic size are also related to cell proliferation. Sox2 has been shown to maintain a neural progenitor state by repressing proneural genes (Agathocleous et al., 2009; Wen et al., 2008), and Six3orthologues, including Six3.1, promote cell proliferation with transcription-dependent and -independent mechanisms (Appolloni et al., 2008; Del Bene et al., 2004; Gestri et al., 2005). The more pronounced contribution of Sox2 and Six3.2 to telencephalic progenitor proliferation, as suggested by the overexpression of both genes, is however intriguing and again suggests that participation in different transcriptional complexes might best explain the paralogue and regional differences.
This does seem to be the case for the hypothalamus. In humans, SOX2heterozygous or de novo mutations are associated with eye defects and hypopituitarism, and mouse embryos with reduced Sox2expression have small pituitary glands (Kelberman et al., 2006). Providing a molecular mechanism for these defects, we show that Sox2 binds to an Rx3 HNCE, as previously shown for the related Xenopus Raxorthologue (Danno et al., 2008; Kennedy et al., 2004), and, upon overexpression, expands the hypothalamic
domain. Although not tested, Sox2 effects in the hypothalamus might also be mediated by Nkx2.1 and Shh, which are, respectively, postulated and proven targets of Sox2 activity (Favaro et al., 2009; Que et al., 2007). Despite expression overlap and expansion of the Six3.2domain upon Sox2 injection, Six3.2 gain-of-function resulted in an opposite phenotype, with a marked reduction of the Rx3- and Nkx2.1-positive hypothalamic region. This suggests that, in the hypothalamus, Sox2and Six3.2act in at least partially independent pathways and are likely to compete for Rx3and possibly Nkx2.1regulation (Fig. 8). This differs from what has been observed in mammals, where Six3positively contributes to hypothalamic development (Gaston-Massuet et al., 2008; Jeong et al., 2008; Lavado et al., 2008). This discrepancy could be easily explained by the proposed subfunctionalisation of the medaka fish paralogues, whereby Six3.2and Six3.1would respectively favour dorsal and ventral forebrain development. According to this hypothesis, hypothalamic development (not shown) and ventral prosencephalic specification (Carl et al., 2002) would be impaired in Six3.1morphants.
In conclusion, our study highlights the role of a TF core composed of Sox2, Six3.2, Foxg1and Rx3in medaka forebrain development, uncovering new aspects of Six3 and Sox2 function. Similar regulatory mechanisms might be at least partially conserved in mammals, as suggested by the phenotypes caused by loss-of-function or haploinsufficiency of Six3 or Sox2 (Geng et al., 2008; Kelberman et al., 2008; Morrison et al., 2002; Suh et al., 2010).
Acknowledgements
We thank I. Dompablo and N. Tabanera for excellent technical assistance and other members of the laboratory for advice and support; Drs J. R. Martinez-Morales, M. Nieto and E. Martí for critical reading of the manuscript; and J. L. Gomez-Skarmeta, E. Martí, S. Nicolis and J. Wittbrodt for sharing plasmids.
Funding
This work was supported by grants from the Spanish Ministry of Science and Innovation (MICINN) [BFU2007-61774, BFU2010-16031]; Comunidad Autonoma de Madrid (CAM) [P-SAL-0190-2006]; and The Centre for Biomedical Network Research on Rare Diseases (CIBERER) to P.B. L.B. holds a predoctoral contract from the CAM, E.C. is supported by the CIBERER, and I.C. held a Telethon [GFP03007] postdoctoral fellowship.
Competing interests statement
The authors declare no competing financial interests.
Supplementary material
Supplementary material available online at
http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.067660/-/DC1
References
Abe, Y., Oka, A., Mizuguchi, M., Igarashi, T., Ishikawa, S., Aburatani, H., Yokoyama, S., Asahara, H., Nagao, K., Yamada, M. et al.(2009). EYA4, deleted in a case with middle interhemispheric variant of holoprosencephaly, interacts with SIX3 both physically and functionally. Hum. Mutat. 30, E946-E955.
Agathocleous, M., Iordanova, I., Willardsen, M. I., Xue, X. Y., Vetter, M. L., Harris, W. A. and Moore, K. B.(2009). A directional Wnt/beta-catenin-Sox2-proneural pathway regulates the transition from proliferation to differentiation in the Xenopusretina. Development136, 3289-3299.
Alfano, G., Vitiello, C., Caccioppoli, C., Caramico, T., Carola, A., Szego, M. J., McInnes, R. R., Auricchio, A. and Banfi, S.(2005). Natural antisense transcripts associated with genes involved in eye development. Hum. Mol. Genet. 14, 913-923.
Appolloni, I., Calzolari, F., Corte, G., Perris, R. and Malatesta, P.(2008). Six3 controls the neural progenitor status in the murine CNS. Cereb. Cortex18, 553-562.
Archer, T. C., Jin, J. and Casey, E. S.(2011). Interaction of Sox1, Sox2, Sox3 and Oct4 during primary neurogenesis. Dev. Biol. 350, 429-440.
Avilion, A. A., Nicolis, S. K., Pevny, L. H., Perez, L., Vivian, N. and Lovell-Badge, R.(2003). Multipotent cell lineages in early mouse development depend
on SOX2 function. Genes Dev. 17, 126-140.
D
E
V
E
LO
P
M
E
N
Bonn, S. and Furlong, E. E.(2008). cis-Regulatory networks during development: a view of Drosophila. Curr. Opin. Genet. Dev. 18, 513-520.
Bovolenta, P., Mallamaci, A., Puelles, L. and Boncinelli, E.(1998). Expression pattern of cSix3, a member of the Six/sine oculis family of transcription factors.
Mech. Dev. 70, 201-203.
Boyer, L. A., Lee, T. I., Cole, M. F., Johnstone, S. E., Levine, S. S., Zucker, J. P., Guenther, M. G., Kumar, R. M., Murray, H. L., Jenner, R. G. et al.(2005). Core transcriptional regulatory circuitry in human embryonic stem cells. Cell122, 947-956.
Carl, M., Loosli, F. and Wittbrodt, J.(2002). Six3 inactivation reveals its essential role for the formation and patterning of the vertebrate eye. Development129, 4057-4063.
Cavodeassi, F., Carreira-Barbosa, F., Young, R. M., Concha, M. L., Allende, M. L., Houart, C., Tada, M. and Wilson, S. W.(2005). Early stages of zebrafish eye formation require the coordinated activity of Wnt11, Fz5, and the Wnt/beta-catenin pathway. Neuron47, 43-56.
Chew, L. J. and Gallo, V.(2009). The Yin and Yang of Sox proteins: activation and repression in development and disease. J. Neurosci. Res. 87, 3277-3287. Conte, I. and Bovolenta, P.(2007). Comprehensive characterization of the
cis-regulatory code responsible for the spatio-temporal expression of olSix3.2 in the developing medaka forebrain. Genome Biol. 8, R137.
Conte, I., Carrella, S., Avellino, R., Karali, M., Marco-Ferreres, R., Bovolenta, P. and Banfi, S.(2010a). miR-204 is required for lens and retinal development via Meis2 targeting. Proc. Natl. Acad. Sci. USA107, 15491-15496.
Conte, I., Marco-Ferreres, R., Beccari, L., Cisneros, E., Ruiz, J. M., Tabanera, N. and Bovolenta, P.(2010b). Proper differentiation of photoreceptors and amacrine cells depends on a regulatory loop between NeuroD and Six6.
Development 137, 2307-2317.
Danno, H., Michiue, T., Hitachi, K., Yukita, A., Ishiura, S. and Asashima, M. (2008). Molecular links among the causative genes for ocular malformation: Otx2 and Sox2 coregulate Rax expression. Proc. Natl. Acad. Sci. USA 105, 5408-5413.
Dateki, S., Kosaka, K., Hasegawa, K., Tanaka, H., Azuma, N., Yokoya, S., Muroya, K., Adachi, M., Tajima, T., Motomura, K. et al.(2010).
Heterozygous orthodenticle homeobox 2 mutations are associated with variable pituitary phenotype. J. Clin. Endocrinol. Metab. 95, 756-764.
Del Bene, F., Tessmar-Raible, K. and Wittbrodt, J.(2004). Direct interaction of geminin and Six3 in eye development. Nature427, 745-749.
Deschet, K., Bourrat, F., Ristoratore, F., Chourrout, D. and Joly, J. S.(1999). Expression of the medaka (Oryzias latipes) Ol-Rx3 paired-like gene in two diencephalic derivatives, the eye and the hypothalamus. Mech. Dev. 83, 179-182.
Eisen, J. S. and Smith, J. C.(2008). Controlling morpholino experiments: don’t stop making antisense. Development135, 1735-1743.
Engelen, E., Akinci, U., Bryne, J. C., Hou, J., Gontan, C., Moen, M., Szumska, D., Kockx, C., van Ijcken, W., Dekkers, D. H. et al.(2011). Sox2 cooperates with Chd7 to regulate genes that are mutated in human syndromes. Nat. Genet.
43, 607-611.
England, S. J., Blanchard, G. B., Mahadevan, L. and Adams, R. J.(2006). A dynamic fate map of the forebrain shows how vertebrate eyes form and explains two causes of cyclopia. Development133, 4613-4617.
Esteve, P., Lopez-Rios, J. and Bovolenta, P.(2004). SFRP1 is required for the proper establishment of the eye field in the medaka fish. Mech. Dev. 121, 687-701.
Favaro, R., Valotta, M., Ferri, A. L., Latorre, E., Mariani, J., Giachino, C., Lancini, C., Tosetti, V., Ottolenghi, S., Taylor, V. et al.(2009). Hippocampal development and neural stem cell maintenance require Sox2-dependent regulation of Shh. Nat. Neurosci. 12, 1248-1256.
Ferri, A. L., Cavallaro, M., Braida, D., Di Cristofano, A., Canta, A., Vezzani, A., Ottolenghi, S., Pandolfi, P. P., Sala, M., DeBiasi, S. et al.(2004). Sox2 deficiency causes neurodegeneration and impaired neurogenesis in the adult mouse brain. Development131, 3805-3819.
Gaston-Massuet, C., Andoniadou, C. L., Signore, M., Sajedi, E., Bird, S., Turner, J. M. and Martinez-Barbera, J. P.(2008). Genetic interaction between the homeobox transcription factors HESX1 and SIX3 is required for normal pituitary development. Dev. Biol. 324, 322-333.
Geng, X., Speirs, C., Lagutin, O., Inbal, A., Liu, W., Solnica-Krezel, L., Jeong, Y., Epstein, D. J. and Oliver, G.(2008). Haploinsufficiency of Six3 fails to activate Sonic hedgehog expression in the ventral forebrain and causes holoprosencephaly. Dev. Cell 15, 236-247.
Gestri, G., Carl, M., Appolloni, I., Wilson, S. W., Barsacchi, G. and
Andreazzoli, M.(2005). Six3 functions in anterior neural plate specification by promoting cell proliferation and inhibiting Bmp4 expression. Development132, 2401-2413.
Hagstrom, S. A., Pauer, G. J., Reid, J., Simpson, E., Crowe, S., Maumenee, I. H. and Traboulsi, E. I.(2005). SOX2 mutation causes anophthalmia, hearing loss, and brain anomalies. Am. J. Med. Genet. A138, 95-98.
Heimbucher, T., Murko, C., Bajoghli, B., Aghaallaei, N., Huber, A., Stebegg, R., Eberhard, D., Fink, M., Simeone, A. and Czerny, T.(2007). Gbx2 and
Otx2 interact with the WD40 domain of Groucho/Tle corepressors. Mol. Cell. Biol. 27, 340-351.
Hoch, R. V., Rubenstein, J. L. and Pleasure, S.(2009). Genes and signaling events that establish regional patterning of the mammalian forebrain. Semin. Cell Dev. Biol. 20, 378-386.
Houart, C., Caneparo, L., Heisenberg, C., Barth, K., Take-Uchi, M. and Wilson, S.(2002). Establishment of the telencephalon during gastrulation by local antagonism of Wnt signaling. Neuron35, 255-265.
Huang, K. M., Dentchev, T. and Stambolian, D.(2008). MiRNA expression in the eye. Mamm. Genome19, 510-516.
Iwamatsu, T.(2004). Stages of normal development in the medaka Oryzias latipes. Mech. Dev. 121, 605-618.
Jeong, Y., Leskow, F. C., El-Jaick, K., Roessler, E., Muenke, M., Yocum, A., Dubourg, C., Li, X., Geng, X., Oliver, G. et al.(2008). Regulation of a remote Shh forebrain enhancer by the Six3 homeoprotein. Nat. Genet. 40, 1348-1353. Kamachi, Y., Uchikawa, M. and Kondoh, H.(2000). Pairing SOX off: with
partners in the regulation of embryonic development. Trends Genet. 16, 182-187.
Kelberman, D., Rizzoti, K., Avilion, A., Bitner-Glindzicz, M., Cianfarani, S., Collins, J., Chong, W. K., Kirk, J. M., Achermann, J. C., Ross, R. et al. (2006). Mutations within Sox2/SOX2 are associated with abnormalities in the hypothalamo-pituitary-gonadal axis in mice and humans. J. Clin. Invest. 116, 2442-2455.
Kelberman, D., de Castro, S. C., Huang, S., Crolla, J. A., Palmer, R., Gregory, J. W., Taylor, D., Cavallo, L., Faienza, M. F., Fischetto, R. et al.(2008). SOX2 Plays a critical role in the pituitary, forebrain, and eye during human embryonic development. J. Clin. Endocrinol. Metab. 93, 1865-1873.
Kelley, C. G., Lavorgna, G., Clark, M. E., Boncinelli, E. and Mellon, P. L. (2000). The Otx2 homeoprotein regulates expression from the gonadotropin-releasing hormone proximal promoter. Mol. Endocrinol. 14, 1246-1256. Kennedy, B. N., Stearns, G. W., Smyth, V. A., Ramamurthy, V., van Eeden, F.,
Ankoudinova, I., Raible, D., Hurley, J. B. and Brockerhoff, S. E.(2004). Zebrafish rx3 and mab21l2 are required during eye morphogenesis. Dev. Biol.
270, 336-349.
Kim, C. H., Oda, T., Itoh, M., Jiang, D., Artinger, K. B., Chandrasekharappa, S. C., Driever, W. and Chitnis, A. B.(2000). Repressor activity of Headless/Tcf3 is essential for vertebrate head formation. Nature407, 913-916.
Kobayashi, M., Toyama, R., Takeda, H., Dawid, I. B. and Kawakami, K. (1998). Overexpression of the forebrain-specific homeobox gene six3 induces rostral forebrain enlargement in zebrafish. Development125, 2973-2982. Kobayashi, M., Nishikawa, K., Suzuki, T. and Yamamoto, M.(2001). The
homeobox protein Six3 interacts with the Groucho corepressor and acts as a transcriptional repressor in eye and forebrain formation. Dev. Biol. 232, 315-326. Kobayashi, D., Kobayashi, M., Matsumoto, K., Ogura, T., Nakafuku, M. and
Shimamura, K.(2002). Early subdivisions in the neural plate define distinct competence for inductive signals. Development129, 83-93.
Kumar, J. P.(2009). The sine oculis homeobox (SIX) family of transcription factors as regulators of development and disease. Cell. Mol. Life Sci. 66, 565-583. Kurokawa, D., Kiyonari, H., Nakayama, R., Kimura-Yoshida, C., Matsuo, I.
and Aizawa, S.(2004). Regulation of Otx2 expression and its functions in mouse forebrain and midbrain. Development131, 3319-3331.
Lagutin, O. V., Zhu, C. C., Kobayashi, D., Topczewski, J., Shimamura, K., Puelles, L., Russell, H. R., McKinnon, P. J., Solnica-Krezel, L. and Oliver, G. (2003). Six3 repression of Wnt signaling in the anterior neuroectoderm is essential for vertebrate forebrain development. Genes Dev. 17, 368-379. Lavado, A., Lagutin, O. V. and Oliver, G.(2008). Six3 inactivation causes
progressive caudalization and aberrant patterning of the mammalian diencephalon. Development135, 441-450.
Loosli, F., Koster, R. W., Carl, M., Krone, A. and Wittbrodt, J.(1998). Six3, a medaka homologue of the Drosophilahomeobox gene sine oculis is expressed in the anterior embryonic shield and the developing eye. Mech. Dev. 74, 159-164.
Loosli, F., Winkler, S. and Wittbrodt, J.(1999). Six3 overexpression initiates the formation of ectopic retina. Genes Dev. 13, 649-654.
Loosli, F., Winkler, S., Burgtorf, C., Wurmbach, E., Ansorge, W., Henrich, T., Grabher, C., Arendt, D., Carl, M., Krone, A. et al.(2001). Medaka eyeless is the key factor linking retinal determination and eye growth. Development128, 4035-4044.
Lopez-Rios, J., Gallardo, M. E., Rodriguez de Cordoba, S. and Bovolenta, P. (1999). Six9 (Optx2), a new member of the six gene family of transcription factors, is expressed at early stages of vertebrate ocular and pituitary development. Mech. Dev. 83, 155-159.
Lopez-Rios, J., Tessmar, K., Loosli, F., Wittbrodt, J. and Bovolenta, P.(2003). Six3 and Six6 activity is modulated by members of the groucho family.
Development130, 185-195.
Lopez-Rios, J., Esteve, P., Ruiz, J. M. and Bovolenta, P.(2008). The Netrin-related domain of Sfrp1 interacts with Wnt ligands and antagonizes their activity in the anterior neural plate. Neural Dev. 3, 19.
Manavathi, B., Peng, S., Rayala, S. K., Talukder, A. H., Wang, M. H., Wang, R. A., Balasenthil, S., Agarwal, N., Frishman, L. J. and Kumar, R.(2007).