COMMENTARY
Loss-of-function approaches in comparative physiology: is there a
future for knockdown experiments in the era of genome editing?
Alex M. Zimmer1,*,‡, Yihang K. Pan1,*, Theanuga Chandrapalan2, Raymond W. M. Kwong2and Steve F. Perry1ABSTRACT
Loss-of-function technologies, such as morpholino- and RNAi-mediated gene knockdown, and TALEN- and CRISPR/Cas9-mediated gene knockout, are widely used to investigate gene function and its physiological significance. Here, we provide a general overview of the various knockdown and knockout technologies commonly used in comparative physiology and discuss the merits and drawbacks of these technologies with a particular focus on research conducted in zebrafish. Despite their widespread use, there is an ongoing debate surrounding the use of knockdown versus knockout approaches and their potential off-target effects. This debate is primarily fueled by the observations that, in some studies, knockout mutants exhibit phenotypes different from those observed in response to knockdown using morpholinos or RNAi. We discuss the current debate and focus on the discrepancies between knockdown and knockout phenotypes, providing literature and primary data to show that the different phenotypes are not necessarily a direct result of the off-target effects of the knockdown agents used. Nevertheless, given the recent evidence of some knockdown phenotypes being recapitulated in knockout mutants lacking the morpholino or RNAi target, we stress that results of knockdown experiments need to be interpreted with caution. We ultimately argue that knockdown experiments should not be discontinued if proper control experiments are performed, and that with careful interpretation, knockdown approaches remain useful to complement the limitations of knockout studies (e.g. lethality of knockout and compensatory responses).
KEY WORDS: CRISPR/Cas9, RNAi, Morpholino, Reverse genetics, Knockdown, Knockout
Introduction
Techniques in loss-of-function (LOF) experiments have continually expanded in their applicability, ease of use and affordability. As such, LOF experiments, involving either knockdown or knockout approaches (see Glossary), are now used widely in many research fields, including comparative physiology. Knockdown refers to the transcriptional or translational repression of gene expression, ultimately leading to a reduction in protein expression, without any genome modifications. The most widely used knockdown techniques are RNA interference (RNAi) and morpholino antisense oligonucleotide knockdown. Importantly, knockdown approaches often result in only a‘partial’LOF, whereby some protein function may be retained. By contrast, knockout refers to a mutation in the
genome aimed at ablating gene function. LOF mutations may be null (amorphic; see Glossary), resulting in the complete loss of gene function, or may only partially reduce gene function (hypomorphic; see Glossary), depending on the nature of the mutation (Housden et al., 2017). While mutant lines (see Glossary) in many species were initially established through forward genetic screens (Brenner, 1974; Nüsslein-volhard and Wieschaus, 1980; Haffter et al., 1996; Driever et al., 1996; Nolan et al., 2000; de Angelis et al., 2000), the recent development of zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs) and clustered regularly interspaced palindromic repeats (CRISPR)/CRISPR-associated nuclease (Cas) has allowed for precise targeted gene knockout.
Currently, the merits of knockdown versus knockout approaches in biological research are being debated. Since the advent of relatively simple genetic knockout approaches ( primarily CRISPR/ Cas), there is a growing sentiment that these techniques should be considered as‘gold standards’for determining gene function and that knockdown approaches, which often result in only a partial LOF, should be used only when null mutants are available for comparison (Schulte-Merker and Stainier, 2014; Lawson, 2016; Stainier et al., 2017). The view that knockdown approaches should be used only when comparison can be made to a null mutant stems mainly from reports of off-target effects (see Glossary): either binding of knockdown reagents to non-target gene products or toxicity leading to non-specific phenotypes, with the use of RNAi and morpholino knockdown (Jackson et al., 2003, 2006a; Jackson and Linsley, 2004; Aleman et al., 2007; Robu et al., 2007; Kok et al., 2015; Joris et al., 2017). Knockout approaches are thought to have lower incidences of off-target effects compared with RNAi or morpholino knockdown, and thus are argued to be the preferred LOF approach moving forward. Perhaps most worrying is that in a screen of more than 24 genes, 80% of morphant (morpholino-treated; see Glossary) zebrafish did not phenocopy individuals carrying null mutations of the targeted gene, suggesting widespread off-target effects of morpholinos (Kok et al., 2015) and spurring debate in some fields (e.g. Faucherre et al., 2014, 2016; Shmukler et al., 2015, 2016). Here, we provide experimental evidence and present support from the literature to argue that differences between knockdown and knockout-induced phenotypes may not always be a result of non-specific, off-target effects in carefully controlled knockdown experiments. We advocate that knockdown approaches should continue to be used in comparative physiology research.
Knockdown techniques RNA interference
The development of RNAi for targeted gene knockdown was pioneered by Andrew Fire and Craig Mello, who–along with co-workers – discovered that the administration of double-stranded RNA (dsRNA) targeting unc-22 in Caenorhabditis elegans
produced phenotypes that mimicked those ofunc-22null mutants (Fire et al., 1998). RNAi can be achieved through dsRNA 1Department of Biology, University of Ottawa, Ottawa, ON K1N 6N5, Canada.
2Department of Biology, York University, Toronto, ON M3J 1P3, Canada.
*These authors contributed equally to this work
‡Author for correspondence ([email protected])
A.M.Z., 0000-0002-4574-1334
Journal
of
Experimental
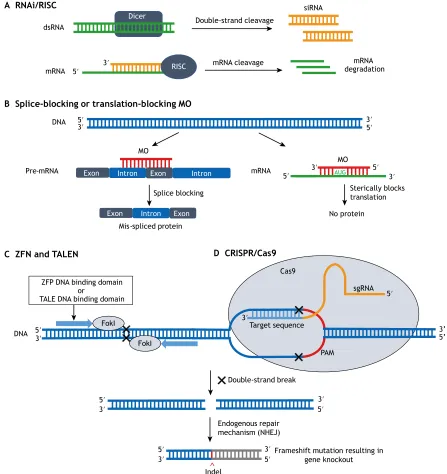
administration or introduction of short hairpin RNA (shRNA) expression vectors (Hannon, 2002; Meister and Tuschl, 2004; Rao et al., 2009). While the cellular processing of dsRNA and shRNA differ, both ultimately produce small/short interfering RNA (siRNA) that is incorporated into the RNA-induced silencing complex (RISC). The antisense siRNA strand guides RISC to the target mRNA for degradation or translational repression (Fig. 1A; Hannon, 2002; Novina and Sharp, 2004; Carthew and Sontheimer, 2009; Rao et al., 2009; Wilson and Doudna, 2013; Housden et al., 2017). Importantly, mechanisms of RNAi may differ between species (Meister and Tuschl, 2004).
RNAi-mediated gene knockdown has been used in an attempt to silence gene expression in zebrafish (Wargelius et al., 1999),
Drosophila(Kennerdell and Carthew, 1998) and mice (Wianny and
Zernicka-Goetz, 2000), and is used extensively in comparative physiology research (Table 1), although concerns have been raised about off-target effects of RNAi. In zebrafish, injection of dsRNA designed to targettbx16/sptproduced novel phenotypes not seen in
tbx16/spt null mutants (Oates et al., 2000). Moreover, while
injection of the dsRNA resulted in a dose-dependent decrease in
tbx16/sptmRNA, it also resulted in a similar decrease in the mRNA
of non-target genes. Most strikingly, injection of dsRNA targeting thelacZgene ofEscherichia coli, for which there is no homolog in zebrafish, caused the same phenotype and decrease in tbx16/spt
mRNA as thetbx16/sptdsRNA (Oates et al., 2000). Several studies have demonstrated off-target effects of RNAi at the transcript, protein and phenotype levels (Jackson et al., 2003, 2006a; Scacheri et al., 2004; Lin et al., 2005; Aleman et al., 2007). These effects are suggested to result from binding to non-target transcripts, activation of the interferon immune response or interference with endogenous micro-RNA (miRNA) pathways (Jackson and Linsley, 2004; Cullen, 2006; Svoboda, 2007; Seok et al., 2018). There is evidence, however, that specific chemical modifications of dsRNA can limit off-target effects (Jackson et al., 2006b). Compared with dsRNA, shRNA is thought to produce fewer off-target effects because this approach utilizes endogenous miRNA cellular processing pathways and is thus less likely to trigger immune responses (Rao et al., 2009).
Morpholino antisense oligonucleotides
Morpholino antisense oligonucleotides (morpholinos) are another class of antisense knockdown reagent that consist of a 6-member morpholine ring backbone and phosphorodiamidate inter-subunit linkages (Summerton, 2007). Morpholinos [typically about 25 base pairs (bp) in length] can be designed to bind to the start of the coding region of an mRNA or to the intron/exon boundaries of a pre-mRNA to block translation or splicing, respectively, by complementary base-pairing (Fig. 1B). Morpholinos were first demonstrated to be capable of phenocopying null mutant phenotypes in frogs (Heasman et al., 2000) and zebrafish (Nasevicius and Ekker, 2000), and rapidly became an important tool for assessing gene function in biological research (Ekker, 2000; Corey and Abrams, 2001; Heasman, 2002; Bill et al., 2009). Guidelines for morpholino use have since been established (Ekker and Larson, 2001; Eisen and Smith, 2008; Bill et al., 2009; Stainier et al., 2017; Xin and Duan, 2018).
The use of morpholino oligonucleotides has expanded markedly to include several species and applications in comparative physiology research (Table 1). Initially, morpholinos were used in early embryonic/larval stages, where they are directly administered into embryos at the one-cell stage, limiting effective knockdown to a narrow window of early-life development (Bill et al., 2009). However, the advent of vivo-morpholinos (Morcos et al., 2008; Moulton and Shan, 2009; Ferguson et al., 2014a) possessing a membrane-penetrating, eight-guanidine conjugate has allowed gene knockdown in adults and in cell cultures (Table 1). As controls, a morpholino in which 5 bases of the original sequence are altered to prevent binding to the target mRNA or a standard control morpholino that has no biological activity except in reticulocytes of humans with a specific mutation linked toβ-thalassemia can be used (Gene-Tools, LLC).
Like RNAi, morpholinos are also prone to off-target effects. Ekker and Larson (2001) reported that 15–20% of morpholinos display off-target phenotypic effects ( phenotypes that do not copy known null mutations), even when dose is carefully controlled. These off-target phenotypes are generally characterized by reduced head and eye size, notochord malformations and craniofacial defects. One major cause of these effects is the activation of p53-mediated apoptosis, which can be attenuated by co-injection of a morpholino that targets the p53 gene (Robu et al., 2007). In addition to p53 activation, evidence from Xenopus tropicalis has demonstrated widespread regulation of non-target genes, primarily those with immune function, and mis-splicing of non-target genes in response to morpholino injection (Gentsch et al., 2018). Indeed, there is now mounting evidence, derived primarily from zebrafish, that morphants often do not phenocopy null mutants (Robu et al., 2007; Gerety and Wilkinson, 2011; Kok et al., 2015; Novodvorsky et al., 2015; Rossi et al., 2015; Shmukler et al., 2015; Eve et al., 2017; Joris et al., 2017). In addition, unlike RNAi, where many of the mechanisms of off-target effects have been characterized (Jackson and Linsley, 2004; Seok et al., 2018), the mechanisms of p53 activation, immune responses, splice defects and non-target binding are not as well understood for morpholinos, and minimizing off-target effects may therefore be challenging.
Knockout techniques
Genome editing using targetable nucleases such as ZFNs (Bibikova et al., 2002; Doyon et al., 2008; Geurts et al., 2009; Urnov et al., 2010), TALENs (Huang et al., 2011; Tesson et al., 2011; Joung and Sander, 2013) and CRISPR/Cas9 (Cong et al., 2013; Jinek et al., 2013; Mali et al., 2013a; Hwang et al., 2013) allows for the generation of specific gene knockouts exhibiting complete and Glossary
Genetic compensation
Changes in RNA or protein levels that can functionally compensate for the loss of function of another gene.
Hypomorphic mutation
Mutation that leads to only a partial loss or reduction in gene function. Knockdown
Perturbation at the DNA, RNA or protein level that reduces the amount of functional protein.
Knockout
Genetic perturbation that completely ablates gene function. Morphant
Individual treated with morpholino antisense oligonucleotide. Mutant
Individual carrying a knockout mutation in the genome. Null (amorphic) mutation
Mutation that leads to complete loss of gene function. Off-target effects
Non-specific effects of loss-of-function reagents resulting from targeting of unintended genes or from toxic effects such as activation of immune or apoptotic pathways.
Transcriptional adaptation
Changes in RNA levels that result from a genetic mutation and not from the loss of gene function.
Journal
of
Experimental
permanent silencing of target gene function (Box 1). These techniques introduce double-strand breaks (DSBs) at specific sites, and error-prone cellular repair mechanisms introduce insertion or deletion mutations when DSBs are repaired. Repair occurs either through non-homologous end-joining (NHEJ; Fig. 1C,D) or homology-directed repair (HDR) when a donor DNA template is present. Knockout techniques are generally used at the single-cell embryo stage, with the aim of inducing a heritable mutation in the germline. This results in the generation of homozygous mutants that can be used for experiments at most stages of the organism’s life
cycle (see Table 1). Exceptions exist where homozygous mutants are non-viable or more subtly where the homozygous mutant is viable, yet the gene of interest has early developmental effects, resulting in subsequent phenotypic analysis being unable to identify the primary function of the gene of interest.
ZFNs and TALENs
Chimeric ZFNs and TALENs are composed of a DNA-binding domain that recognizes specific sequences (Box 1), and a restriction enzyme FokI nuclease domain for DNA cleavage. However, Double-strand cleavage
siRNA
dsRNA
mRNA
Dicer
mRNA cleavage mRNA
degradation A RNAi/RISC
5⬘
3⬘ RISC
DNA
Sterically blocks translation
No protein MO mRNA
Pre-mRNA Exon Intron Exon Intron
MO
Splice blocking
Mis-spliced protein
Exon Intron Exon
B Splice-blocking or translation-blocking MO
5⬘
3⬘
3⬘
5⬘
5⬘
3⬘ 5⬘
3⬘
AUG
Double-strand break
Endogenous repair mechanism (NHEJ)
C ZFN and TALEN D CRISPR/Cas9
Frameshift mutation resulting in gene knockout
3⬘
5⬘
Cas9
DNA 3’5’
sgRNA
PAM Target sequence
3⬘
5⬘
FokI ZFP DNA binding domain
or
TALE DNA binding domain
5⬘
3⬘
5⬘
3⬘
5⬘
3⬘
5⬘
3⬘
[image:3.612.81.528.58.532.2]^
Indel FokIFig. 1. Mechanisms of loss-of-function technologies.(A) Gene knockdown via RNA interference (RNAi). RNA-induced silencing complex (RISC) is mediated by Dicer to produce small interfering RNA (siRNA), which guides RISC to the target mRNA, initiating its degradation. (B) Splice-blocking or translation-blocking morpholinos result in a mis-spliced or absent protein, respectively. (C) ZFN and TALEN, and (D) CRISPR/Cas9 introduce double-strand breaks (DSBs) in target DNA. These DSBs are repaired by non-homologous end-joining (NHEJ), causing frameshift mutations and ultimate knockout of the gene. Abbreviations: Cas9, CRISPR-associated protein 9; FokI, restriction endonuclease FokI; Indel, insertions/deletions; MO, morpholino; PAM, protospacer adjacent motif; sgRNA, single-guide RNA; TALEN, transcription activator-like effector nuclease; ZFN, zinc-finger nuclease. See Box 1 for further details on the mechanisms of programmable nucleases for gene editing.
Journal
of
Experimental
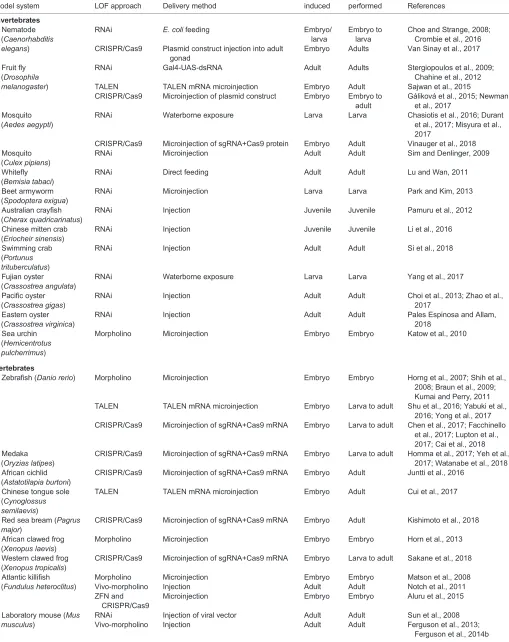
Table 1. Examples of LOF studies in comparative physiology
Model system LOF approach Delivery method
LOF induced
Experiment
performed References
Invertebrates Nematode (Caenorhabditis elegans)
RNAi E. colifeeding Embryo/ larva
Embryo to larva
Choe and Strange, 2008; Crombie et al., 2016 CRISPR/Cas9 Plasmid construct injection into adult
gonad
Embryo Adults Van Sinay et al., 2017
Fruit fly (Drosophila melanogaster)
RNAi Gal4-UAS-dsRNA Adult Adults Stergiopoulos et al., 2009;
Chahine et al., 2012
TALEN TALEN mRNA microinjection Embryo Adult Sajwan et al., 2015
CRISPR/Cas9 Microinjection of plasmid construct Embryo Embryo to adult
Gáliková et al., 2015; Newman et al., 2017
Mosquito (Aedes aegypti)
RNAi Waterborne exposure Larva Larva Chasiotis et al., 2016; Durant
et al., 2017; Misyura et al., 2017
CRISPR/Cas9 Microinjection of sgRNA+Cas9 protein Embryo Adult Vinauger et al., 2018 Mosquito
(Culex pipiens)
RNAi Microinjection Adult Adult Sim and Denlinger, 2009
Whitefly (Bemisia tabaci)
RNAi Direct feeding Adult Adult Lu and Wan, 2011
Beet armyworm (Spodoptera exigua)
RNAi Microinjection Larva Larva Park and Kim, 2013
Australian crayfish (Cherax quadricarinatus)
RNAi Injection Juvenile Juvenile Pamuru et al., 2012
Chinese mitten crab (Eriocheir sinensis)
RNAi Injection Juvenile Juvenile Li et al., 2016
Swimming crab (Portunus trituberculatus)
RNAi Injection Adult Adult Si et al., 2018
Fujian oyster (Crassostrea angulata)
RNAi Waterborne exposure Larva Larva Yang et al., 2017
Pacific oyster (Crassostrea gigas)
RNAi Injection Adult Adult Choi et al., 2013; Zhao et al.,
2017 Eastern oyster
(Crassostrea virginica)
RNAi Injection Adult Adult Pales Espinosa and Allam,
2018 Sea urchin
(Hemicentrotus pulcherrimus)
Morpholino Microinjection Embryo Embryo Katow et al., 2010
Vertebrates
Zebrafish (Danio rerio) Morpholino Microinjection Embryo Embryo Horng et al., 2007; Shih et al., 2008; Braun et al., 2009; Kumai and Perry, 2011 TALEN TALEN mRNA microinjection Embryo Larva to adult Shu et al., 2016; Yabuki et al.,
2016; Yong et al., 2017 CRISPR/Cas9 Microinjection of sgRNA+Cas9 mRNA Embryo Larva to adult Chen et al., 2017; Facchinello
et al., 2017; Lupton et al., 2017; Cai et al., 2018 Medaka
(Oryzias latipes)
CRISPR/Cas9 Microinjection of sgRNA+Cas9 mRNA Embryo Larva to adult Homma et al., 2017; Yeh et al., 2017; Watanabe et al., 2018 African cichlid
(Astatotilapia burtoni)
CRISPR/Cas9 Microinjection of sgRNA+Cas9 mRNA Embryo Adult Juntti et al., 2016
Chinese tongue sole (Cynoglossus semilaevis)
TALEN TALEN mRNA microinjection Embryo Adult Cui et al., 2017
Red sea bream (Pagrus major)
CRISPR/Cas9 Microinjection of sgRNA+Cas9 mRNA Embryo Adult Kishimoto et al., 2018
African clawed frog (Xenopus laevis)
Morpholino Microinjection Embryo Embryo Horn et al., 2013
Western clawed frog (Xenopus tropicalis)
CRISPR/Cas9 Microinjection of sgRNA+Cas9 mRNA Embryo Larva to adult Sakane et al., 2018
Atlantic killifish (Fundulus heteroclitus)
Morpholino Microinjection Embryo Embryo Matson et al., 2008
Vivo-morpholino Injection Adult Adult Notch et al., 2011
ZFN and CRISPR/Cas9
Microinjection Embryo Embryo Aluru et al., 2015
Laboratory mouse (Mus musculus)
RNAi Injection of viral vector Adult Adult Sun et al., 2008
Vivo-morpholino Injection Adult Adult Ferguson et al., 2013;
Ferguson et al., 2014b
Continued
Journal
of
Experimental
construction of sequence-specific ZFNs and TALENs is challenging and time-consuming, which has led to their gradual replacement by CRISPR/Cas. The two main approaches for constructing zinc-finger proteins with unique DNA-binding
specificity are the modular assembly approach and selection-based approach (see Beerli and Barbas, 2002; Ochiai and Yamamoto, 2015 for review). The recognition code of TALEs is simpler than that of zinc fingers (Box 1), but provides similar technical challenges to synthesis due to extensive identical repeat sequences. Several methods have been developed that enable rapid assembly of custom TALE arrays (see Joung and Sander, 2013; Ochiai and Yamamoto, 2015 for review).
CRISPR/Cas
CRISPR has been transformative across nearly all biological and medical science-related fields, and rapidly became the method of choice for genome editing. Unlike ZFNs and TALENs, where each new target site requires the design and construction of a new nuclease, CRISPR/Cas9 uses a CRISPR RNA (crRNA) and a trans-activating crRNA (tracrRNA) that can be engineered into a single guide RNA (sgRNA) to target the Cas9 nuclease to a specific DNA sequence (Box 1; Fig. 1D). For a more comprehensive review of the history and mechanisms of CRISPR/Cas9-mediated genome targeting, see Doudna and Charpentier (2014).
Following the first demonstrations that CRISPR/Cas9 can be repurposed for genome editing in eukaryotic cells (Cong et al., 2013; Mali et al., 2013a; Jinek et al., 2013), this technique was adopted for studies on fruit flies (Bassett et al., 2013; Gratz et al., 2013), zebrafish (Chang et al., 2013; Hwang et al., 2013; Xin and Duan, 2018; Box 2), nematodes (Friedland et al., 2013), mice (Yang et al., 2013) and other organisms (Blitz et al., 2013; Flowers et al., 2014; Niu et al., 2014; Whitworth et al., 2014). CRISPR has also been adopted for LOF experiments to study gene function in physiological processes (Table 1).
Despite the simplicity of CRISPR, some studies have shown that Cas9 nucleases can induce mutations at off-target sites (reviewed by Tsai and Joung, 2016). However, the majority of off-target sites exhibit mutation frequencies at least 2-fold lower than on-target mutation frequency if the off-target site has greater than one bp mismatch compared with the on-target site (Hsu et al., 2013; Pattanayak et al., 2013; Cho et al., 2014). Table 1. Continued
Model system LOF approach Delivery method
LOF induced
Experiment
performed References
TALEN Microinjection of TALEN mRNA into fertilized one-cell embryos
Embryo Adult Wang et al., 2016
CRISPR/Cas9 Electroporation of sgRNA plasmid into embryonic stem cell/microinjection of sgRNA+Cas9 mRNA into fertilized eggs
Embryo Adult Tsuchiya et al., 2015; Davies et al., 2017
Laboratory rat (Rattus norvegicus)
RNAi Tracheal administration/lipofection of plasmid DNA
Adult Adult Li and Folkesson, 2006
Pig (Sus scrofa domesticus)
TALEN Cell transfection followed by somatic cell nuclear transfer
Somatic cell
Juvenile Quadalti et al., 2018
CRISPR/Cas9 Cell transfection followed by somatic cell nuclear transfer
Somatic cell
Juvenile Quadalti et al., 2018
In vitrocell/tissue culture Rainbow trout
(Oncorhynchus mykiss)
RNAi Lipofection of cultured gill cells – – Kolosov and Kelly, 2017;
Kolosov et al., 2017 Laboratory rat (Rattus
norvegicus)
RNAi Reversible chemical permeabilization of cultured brain tissue
– – Wickramasekera et al., 2013
Morpholino Endo-porter delivery into seeded pancreatic INS-1 cells
– – Figeac et al., 2010
CRISPR/Cas9 sgRNA plasmid transfection – – Roy et al., 2014
A non-exhaustive list of studies in comparative physiology that have employed LOF techniques, including the study species, LOF approach, delivery method, life stage at which the LOF was induced and life stage at which the experiments were performed.
Box 1. Mechanism of ZFNs, TALENs and CRISPR/Cas-mediated genome targeting
Zinc-finger nucleases (ZFNs) are artificial restriction enzymes generated by fusing a zinc-finger DNA-binding domain to a DNA-cleavage domain. ZFN specificity is conferred by Cys2–His2 zinc-finger domains, which consist of 30 amino acids arranged in a ββα configuration. These domains bind a specific 3 bp DNA sequence (Beerli and Barbas, 2002). Sequence specificity of zinc-finger proteins can be determined by combining up to three contiguous domains, resulting in a 9 bp recognition sequence. The design of a linker sequence to connect two three-finger proteins further increased ZFN specificity to 18 bp in length, allowing specificity in genomes up to 68 billion bp (Liu et al., 1997). By fusing zinc-finger proteins to the cleavage domain of FokI nuclease, ZFNs capable of cleaving DNA in a sequence-specific manner were generated (Fig. 1C; Kim et al., 1996).
Similarly to ZFNs, the specificity of TALENs is conferred by DNA-binding proteins termed transcription activator-like effectors (TALEs). TALEs are naturally occurring proteins found in phytopathogenic bacteria of the genus
Xanthomonas. The DNA-binding domain of TALE is composed of multiple, nearly identical repeats, each of which comprises∼34 amino acids. Two hypervariable amino acid residues in each repeat recognize one bp in target DNA (Boch et al., 2009; Moscou and Bogdanove, 2009). By joining appropriate repeat units, TALEs can be redesigned to bind to user-defined sequences (Boch et al., 2009), and their fusion with FokI nuclease generated TALENs for genome editing (Fig. 1C; Christian et al., 2010; Li et al., 2010; Mahfouz et al., 2011; Miller et al., 2011).
CRISPR/Cas-mediated genome editing utilizes the microbial adaptive immune system CRISPR. Unlike ZFNs and TALENs, the CRISPR nuclease Cas9 is targeted by a short guide RNA that comprises a constant region (which facilitates the binding of Cas9 nuclease and maintains the complex in its active form) and a target-specific region (typically 20 bp in length) that is complementary to the target DNA sequence (Jinek et al., 2012; Lim et al., 2016). In addition, binding of a protospacer-adjacent motif (PAM) downstream of the target locus is required to direct Cas9-mediated double-strand breaks (Gasiunas et al., 2012).
Journal
of
Experimental
Refinements in the CRISPR/Cas9 system have further improved targeting specificity; these include truncating sgRNA to have fewer than 20 nucleotides of complementarity with its target DNA (Fu et al., 2014), converting Cas9 to a nickase enzyme (Cas9n) requiring two separate Cas9 binding events to occur at
the same locus to result in DNA cleavage (Ran et al., 2013; Mali et al., 2013b), and fusing catalytic residue inactive,‘dead’Cas9 (dCas9) with FokI, requiring dual sgRNAs to coordinate FokI– dCas9 dimerization at a specific locus to enable DNA cleavage (Guilinger et al., 2014; Tsai et al., 2014). Even if off-target effects are unavoidable, unlinked off-target mutations should segregate through breeding when generating a homozygous mutant line (Box 2), and phenotyping of mutants generated from two independent founder animals would be sufficient to establish causality between phenotype and genotype.
The success of CRISPR/Cas9 as a genome editing tool has resulted in a search for additional Cas nucleases for applications in genome editing. Notably, Cas12a ( previously named Cpf1) has emerged as an alternative for Cas9. Cas12a only requires a∼ 44-nucleotide crRNA for activation (Zetsche et al., 2015), which is much shorter than the∼100-nucleotide sgRNA required for Cas9, providing a significant advantage for the production of synthetic guide RNAs. Furthermore, instead of the blunt cuts produced by Cas9, Cas12a generates staggered cuts at target DNA, allowing gene fragments to be inserted in the desired orientation via complementary sticky ends, as in traditional restriction enzyme cloning methods (Zetsche et al., 2015).
The knockdown versus knockout debate
The knockdown versus knockout debate in the zebrafish community has been ongoing for over a decade and has led to the publication of specific guidelines for the use of morpholinos in zebrafish research (Eisen and Smith, 2008; Stainier et al., 2017). Similarly, guidelines for the use and design of RNAi have also been established (Cullen, 2006; Bofill-De Ros and Gu, 2016) and research in RNAi approaches continues to produce methods to minimize off-target effects (Jackson et al., 2006b; Mockenhaupt et al., 2015; Suter et al., 2016). Recently, the knockdown versus knockout debate was spurred by Kok et al. (2015), who demonstrated that in 80% of the 24 genes studied, morphants failed to phenocopy mutants. Some researchers have advocated limiting the use of morpholinos to cases in which comparison can be made to a null mutant phenotype (Schulte-Merker and Stainier, 2014; Lawson, 2016; Stainier et al., 2017). Others propose the continued use of morpholinos even in the absence of a null mutant in order to maintain progress in biological research (Blum et al., 2015).
Even with proper experimental design and careful control of dose, morpholinos and RNAi can exert non-specific off-target effects that are unrelated to target gene function. However, there are several additional reasons for the discrepancy between knockdown and mutant phenotypes that need to be considered to appreciate how the comparison of differences between mutant and morphant phenotypes has likely overestimated the prevalence of off-target effects of this particular knockdown reagent. First, in many of the studies cited by Kok et al. (2015), morpholino dose was not properly controlled, a factor that we demonstrate with experimental evidence to have profound gross morphological off-target effects in zebrafish. Second, transcriptional adaptation (see Glossary) in mutants can lead to genetic compensation (see Glossary) of functionally related gene pathways (El-Brolosy and Stainier, 2017), which would not be observed in knockdown approaches that do not alter the genome. Third, LOF mutant progeny generated from heterozygous parents (Box 2) can carry sufficient wild-type maternal mRNA to maintain normal gene function in the absence of wild-type zygotic mRNA in early life. These maternal mRNA-carrying null mutants will display phenotypes that differ from those generated with knockdown approaches that target both maternal and zygotic mRNA. Finally, Box 2. Workflow for establishing a mutant line of zebrafish
using CRISPR/Cas9 gene editing
CRISPR/Cas9 technology provides an efficient way of generating knockout lines in zebrafish. First, wild-type (WT) zebrafish embryos are microinjected with sgRNA-Cas9 solution at the one-cell stage of development. These embryos are reared to reproductive maturity (∼90 dpf ). Founder fish (F0, mosaic red and blue stripes) are then out-crossed with WT fish (solid blue stripes) to yield heterozygous F1 fish (F1 +/–, solid red and blue stripes). Mature F1 fish that carry the same mutation are in-crossed to obtain homozygous F2 (F2−/−, solid red stripes). Genotypic and phenotypic screening may take place periodically at either the embryonic or adult stage (e.g. using fin biopsy) to ensure the presence of a frameshift mutation.
X
X
Microinjected embryos
Screen for mutagenesis
WT Founder
Genotypic and phenotypic characterization
Genotypic and phenotypic characterization
F1 +/−
F1 +/−
F2 −/−
F0
F1
F2
F0 animals generated by CRISPR/Cas9 technology are often mosaic; to screen for the presence of insertion/deletion mutations from a large number of F0 animals, a variety of methods can be used other than Sanger sequencing. The easiest methods are to take screening into consideration when selecting target sites, such that the target site contains a restriction enzyme site that will be lost along with the lesion; this loss can be identified through PCR/restriction enzyme analysis (Gao et al., 2015; Bhowmik et al., 2018). Alternatively, segmental deletions can be generated by simultaneously targeting multiple sites. This results in fragment size differences between WT and mutant alleles that can be detected through PCR and agarose gel electrophoresis (Tabebordbar et al., 2016). Other screening methods that are applicable to all target sites include T7E1 and surveyor assay (Mashal et al., 1995; Qiu et al., 2004; Guschin et al., 2010), PAGE-based assay (Zhu et al., 2014), high-resolution melting (HRM) curve analysis (Montgomery et al., 2007; Thomas et al., 2014) and annealing at critical temperature (ACT) PCR (Hua et al., 2017). In F1 and F2 animals, the precise alteration of the target loci can be further characterized by using Sanger sequencing.
Journal
of
Experimental
partial knockdown by morpholino or RNAi can result in the complete retention of normal physiological function if the targeted protein has a high affinity for its substrate or has a high catalytic efficiency. In such cases, partial LOF phenotypes that retain normal protein function will differ from amorphic mutant phenotypes.
Dose-specific off-target effects
The prevalence of off-target effects of morpholinos has likely been inflated by inadequate control of dose in previous research. For instance, Topczewska et al. (2001) reported that administration of a morpholino targetingfoxc1aat a dose of 10 ng/embryo resulted in a complete loss of somite boundaries in developing zebrafish larvae. In a later study,foxc1awas knocked out using TALENs, resulting in only mild somite defects (Li et al., 2015). However, whenfoxc1a
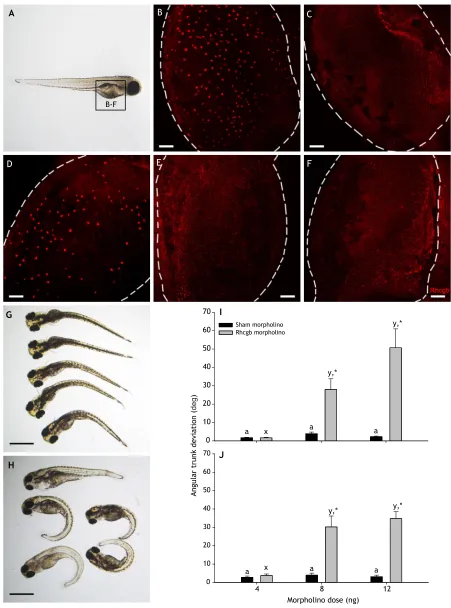
morpholinos were administered at 4 ng per embryo (Topczewska et al., 2001), the resulting phenotype was milder and more similar to the mutant phenotype reported by Li et al. (2015). To further demonstrate how an incorrect dose of morpholino can lead to morphological off-target effects, we performed an experiment with a morpholino designed to block the translation of the ammonia-transporting protein Rhcgb (Shih et al., 2012). At a dose of 4 ng, no gross morphological defects were observed, and Rhcgb expression was effectively abolished, such that the Rhcgb immunostaining pattern was indistinguishable from that of CRISPR/Cas9-generated
rhcgbnull mutants (Fig. 2; see Appendix for details of methods
used). However, at higher doses, a specific phenotype was observed whereby the trunk/tail was curved or kinked, as quantified by the angular deviation from the midline of the body in the dorso–ventral or sagittal planes. Importantly, this effect was not observed in larvae treated with the sham (standard control) morpholino even up to 12 ng per embryo (Fig. 2). This curved trunk phenotype was also observed, and perhaps was even more severe, inrhcgbnull mutants treated with 8–12 ng doses of morpholino (Fig. 2), demonstrating that the observed phenotype can be considered an off-target effect that is present only at morpholino doses that exceed those necessary to effectively knock down Rhcgb protein expression. Importantly, the lack of overt phenotype at the 4 ng dose does not necessarily imply that this morpholino was free of off-target effects. For both morpholinos and RNAi, studies have demonstrated widespread regulation of non-target genes thought to be related to immune responses, splice defects or promiscuous binding in the case of morpholinos (Joris et al., 2017; Gentsch et al., 2018) or to alteration of miRNA pathways by shRNA (Baek et al., 2014). It is possible that these sub-phenotypic effects can have confounding effects on the interpretation of knockdown results.
These experimental results highlight how the proportion of morpholinos that are non-specific can be inflated by poor control of morpholino dose, resulting in off-target effects that have been misinterpreted as the effect of targeted knockdown. Indeed, it is important to point out several features of the morphant phenotypes analyzed by Kok et al. (2015) who concluded that off-target effects occurred in 80% of morphants. First, of the 24 morphant phenotypes analyzed, 7 did not report the information necessary to determine dose. Second, of those doses that were reported, several were >8 ng, a dose that we found to result in a kinked-tail off-target phenotype with the Rhcgb morpholino (Fig. 2). Third, the curved or kinked trunk/tail phenotype that we observed in larvae treated with 8–12 ng Rhcgb morpholino was observed in 11 of 24 morphant phenotypes. In fact, Kok et al. (2015) conceded that a caveat of their study is that the phenotypes they chose to analyze were those most typically associated with off-target effects of morpholinos. It is important to stress here that even with careful control of dose, morpholinos can
still cause off-target effects (Ekker and Larson, 2001; Schulte-Merker and Stainier, 2014; Lawson, 2016), highlighting the rationale for the recent suggestion that this knockdown reagent needs to be validated in a null mutant of the same target (Stainier et al., 2017). Indeed, in our experiment, treating the Rhcgb null mutant with Rhcgb morpholino demonstrated the kinked-tail phenotype is an off-target effect of the high morpholino dose (Fig. 2). However, it is likely that the prevalence of gross morphological off-target effects of morpholinos has been inflated by poor control of reagent dose.
Transcriptional adaptation and genetic compensation Genetic knockout can lead to transcriptional adaptation and resulting compensatory gene regulation through DSB-induced RNAs, mutant mRNA transcripts or RNA-binding proteins, though the exact mechanisms and heritability of these transcriptomic changes are still unclear (El-Brolosy and Stainier, 2017). Since transcriptional adaptation is triggered by changes in the genome, this response would not be observed in knockdown approaches. For instance, Rossi et al. (2015) showed that severe vascular defects observed in Egfl7 knockdown zebrafish were not observed inegfl7mutants, which would suggest off-target effects of Egfl7 knockdown. However, egfl7 knockout resulted in the upregulation of a different class of proteins and genes related to vascular development, which was not observed in response to Egfl7 knockdown, indicating that theegfl7mutants compensated for the loss of Egfl7 function (Rossi et al., 2015). Importantly, knockdown can also result in genetic compensation if the compensatory mechanism is regulated by changes downstream of the gene (i.e. protein function).
Influence of maternal mRNA
Where knockout results in death or loss of reproductive viability, researchers have used F2 homozygous mutant progeny of heterozygous parents to assess LOF outcomes. In these cases, heterozygous mutant parents may pass sufficient wild-type mRNA of the target gene on to mutant offspring, resulting in early developmental expression of the target protein from maternal mRNA (Wu et al., 2003). Alternatively, germ line replacement (Ciruna et al., 2002) can be used to generate wild-type females with homozygous mutant germ lines to obtain mutant offspring without contribution of maternal mRNA; however, this method would not be feasible if the gene of interest is essential for germline development. In contrast, RNAi and translation-blocking morpholinos can knock down the expression of both maternal and zygotic transcripts, uncovering phenotypes that may not be observed in LOF experiments when using mutant offspring generated from heterozygous parents. In instances where maternal proteins are directly deposited into embryos, careful comparisons of mutants generated from heterozygous and homozygous crosses can help elucidate the function of the gene of interest (Cote et al., 2007).
Proteins with high affinity or efficiency
When utilizing knockdown approaches, protein function must be considered. For example, Pena et al. (2017) used morpholinos and CRISPR/Cas9 to knock down or knock out, respectively,aldh7a1. This gene encodes α-aminoadipic-semialdehyde dehydrogenase (antiquitin), an enzyme important in the pathology of pyridoxine-dependent epilepsy (PDE). In the absence of antiquitin, the toxic lysine intermediate piperideine-6-carboxylic acid (P6C) accumulates and causes the pathophysiological effects of PDE. Injection of a splice-blocking morpholino (2.5 ng per embryo) led
Journal
of
Experimental
B-F
A B C
D E F
G
H
I
0 10 20 30 40 50 60 70
Sham morpholino Rhcgb morpholino
a x a a
y,*
y,*
J
Morpholino dose (ng)
4 8 12
Angular trunk deviation (
deg
)
0 10 20 30 40 50 60 70
a x a a
[image:8.612.79.530.54.662.2]y,* y,*
Fig. 2. Off-target effects of Rhcgb morpholino in zebrafish when administered at doses in excess of that causing effective protein knockdown. Anti-Rhcgb whole-mount immunostaining of 4 dpf wild-type zebrafish larvae (A) treated with 4 ng sham (B), 4 ng Rhcgb (C), 12 ng sham (D) and 12 ng Rhcgb (E) morpholino, and of 4 dpfrhcgbmutants (F). Scale bars: 50 µm. Representative images of wild-type (G) andrhcgbmutant (H) larvae injected with 12 ng Rhcgb MO. Scale bars: 0.1 cm. (I,J) Quantitative analysis of the angular deviation from the midline of the body in wild-type (I) andrhcgbmutant (J) larvae injected with 4, 8 or 12 ng sham or Rhcgb morpholino. In I and J, different letters within a morpholino treatment represent statistically significant differences between doses and asterisks represent statistically significant differences of morpholino treatment within a given dose as determined by a two-way ANOVA followed by a Holm–Sıdá ́k
post hoctest (n=16–25; *P<0.05).
Journal
of
Experimental
to a >80% reduction in Aldh7a1 protein expression in 4 dpf zebrafish larvae, yet the levels of P6C were unchanged compared with levels in sham-injected larvae. However, P6C was found to accumulate inaldh7a1null mutants compared with wild-type larvae (Pena et al., 2017). The authors concluded that this difference in P6C accumulation between morphants and mutants is likely due to the high catalytic efficiency of antiquitin for its substrate, such that normal physiological function is maintained even when protein expression is reduced by >80% (Pena et al., 2017). Therefore, protein/enzyme affinity/efficiency is certainly an important feature of physiological systems to bear in mind when interpreting results from partial knockdown of target proteins. This example highlights that, in some instances, complete knockout is the only way to assess gene or protein function in physiological systems, and that maintenance of normal phenotypes in response to partial knockdown should be interpreted cautiously.
Should knockdown approaches continue to be used in comparative physiology?
Knockdown approaches have been used in comparative physiology research for over a decade, and their use continues today (Table 1). These studies have undoubtedly advanced our understanding of various physiological systems, yet the current view in the zebrafish community for morpholinos is that knockdown reagents should be limited to instances where the reagent can be tested in a null mutant background (Schulte-Merker and Stainier, 2014; Lawson, 2016; Stainier et al., 2017). Indeed, this important control has demonstrated that some morphant and RNAi-induced phenotypes can be recapitulated in null mutants lacking the gene target (Baek et al., 2014; Kok et al., 2015; Song et al., 2015; Jahangiri et al., 2016; Liu et al., 2017). While we agree that administration of knockdown reagents in a null background is the most definitive way to prove knockdown specificity, many researchers in comparative physiology work with non-model organisms (Table 1) for which mutant lines may not exist. Furthermore, generation time and husbandry of some species would make it impractical or impossible to establish mutant founder lines. Indeed, the production of mutant lines in zebrafish can take over a year (Box 2), and this species has a relatively short generation time (∼2–3 months). Therefore, we strongly believe that eliminating knockdown experiments as an acceptable approach will limit progress in the field of comparative physiology, in agreement with the sentiment raised previously for developmental research (Blum et al., 2015).
The prevalence of off-target phenotypes arising from knockdown approaches, in particular morpholinos, may have been overestimated in the literature, but these reagents can still produce off-target effects even when reagents and experiments are carefully designed and controlled. However, if researchers in comparative physiology follow some of the earlier guidelines developed for morpholinos and RNAi (Cullen, 2006; Eisen and Smith, 2008) such as: (1) confirmation of knockdown at the transcript or protein level; (2) the use of multiple antisense reagents designed to target the same gene to demonstrate phenotype specificity; (3) concomitant treatment with an antisense reagent control that does not target any gene within the host genome; and (4) careful titration of dose to the lowest causing target knockdown, rather than overt phenotype, we believe that knockdown results should still be admissible in peer-reviewed journals. It is important to bear in mind, however, that without the ability to confirm specificity in a mutant background, results from even the most carefully conducted knockdown
experiments should be interpreted with caution. While the advent of simple and precise gene editing technologies such as CRISPR/ Cas9 has revolutionized LOF approaches, it is clear that knockdown approaches will continue to be used in comparative physiology (Table 1). We advocate that carefully executed and interpreted knockdown experiments in comparative physiology, alongside knockout experiments when possible, will be important for maintaining progress in our understanding of gene function in physiological systems.
Appendix
Generation ofrhcgb−/−null mutant zebrafish
Expression of rhcgb (formerly rhcg1; NCBI gene accession: NM_001320382.1) was knocked out using CRISPR/Cas9. A sgRNA was synthesized using a cloning-free method described previously (Talbot and Amacher, 2014), and the guide sequence of the sgRNA which was specific to a region of exon 1 of rhcgb
(GGGCAACTGCTTCGGCTCCA) was designed using CHOPCHOP (Montague et al., 2014; Labun et al., 2016). sgRNA size and quality were verified by gel electrophoresis, and RNA concentration was determined spectrophotometrically following purification (RNeasy mini kit, QIAGEN). Cas9 mRNA was synthesized from zebrafish codon-optimized Cas9 (pCS2-nls-zCas9-nls; Jao et al., 2013) using mMESSAGE mMACHINE SP6 kit (Invitrogen). Following purification (RNeasy Mini kit, Qiagen), mRNA size and quality was checked by gel electrophoresis and concentration was determined spectrophotometrically.
One-cell stage embryos were injected with 1 nl injection solution containing 150 pg Cas9 mRNA, 50 pg sgRNA and 0.01% Phenol Red suspended in Danieau buffer (Nasevicius and Ekker, 2000). Embryos were reared to sexual maturity (60–90 dpf ) and mutants in this adult F0 population were identified through DNA extraction of fin clips and Sanger sequencing (Genome Quebec, McGill University, Montreal, Canada), which confirmed that a portion (∼80%) of the embryos had a mutatedrhcgbgene. A mutant line was then established following the protocol outlined in Box 1 using founders carrying a 1 bp insertion 3 bp upstream of the PAM. Effective knockout of rhcgb was confirmed by whole-mount immunohistochemistry using an antibody and methods described previously (Nakada et al., 2007).
Morpholino knockdown
One-cell stage embryos were injected with either sham (standard control) morpholino (Gene-Tools, LLC, Philomath, OR, USA; 5′-CCTCTTACCTCAGTTACAATTTATA-3′), which has no biological target in zebrafish, or a translation-blocking morpholino that targeted the translation start site ofrhcgb (5′ -CAGTTGCCCATGTCTACAGCTTGAG-3′; Shih et al., 2012) diluted in Danieau buffer and 0.01% Phenol Red. Effective knockdown of Rhcgb was confirmed by whole-mount immunohistochemistry.
Acknowledgements
The authors wish to thank Christine Archer and ACVS staff at the University of Ottawa and also extend thanks to Dr Izabella Pena for many thoughtful conversations that aided in this work.
Competing interests
The authors declare no competing or financial interests.
Funding
A.M.Z. was supported by a Natural Sciences and Engineering Research Council of Canada (NSERC) postdoctoral fellowship. This research was supported by NSERC Discovery grants (G13017 to S.F.P. and 05984 to R.W.M.K.). R.W.M.K. was also supported by the Canada Research Chairs Program.
Journal
of
Experimental
References
Aleman, L. M., Doench, J. and Sharp, P. A.(2007). Comparison of siRNA-induced off-target RNA and protein effects.RNA13, 385-395.
Aluru, N., Karchner, S. I., Franks, D. G., Nacci, D., Champlin, D. and Hahn, M. E.
(2015). Targeted mutagenesis of aryl hydrocarbon receptor 2a and 2b genes in Atlantic killifish (Fundulus heteroclitus).Aquat. Toxicol.158, 192-201.
Baek, S. T., Kerjan, G., Bielas, S. L., Lee, J. E., Fenstermaker, A. G., Novarino, G. and Gleeson, J. G.(2014). Off-target effect ofdoublecortinfamily shRNA on neuronal migration associated with endogenous microRNA dysregulation.Neuron
82, 1255-1262.
Bassett, A. R., Tibbit, C., Ponting, C. P. and Liu, J.-L.(2013). Highly efficient targeted mutagenesis ofDrosophilawith the CRISPR/Cas9 system.Cell Rep.4, 220-228.
Beerli, R. R. and Barbas, C. F., III(2002). Engineering polydactyl zinc-finger transcription factors.Nat. Biotechnol.20, 135.
Bhowmik, P., Ellison, E., Polley, B., Bollina, V., Kulkarni, M., Ghanbarnia, K., Song, H., Gao, C., Voytas, D. F. and Kagale, S.(2018). Targeted mutagenesis in wheat microspores using CRISPR/Cas9.Sci. Rep.8, 6502.
Bibikova, M., Golic, M., Golic, K. G. and Carroll, D.(2002). Targeted chromosomal cleavage and mutagenesis inDrosophilausing zinc-finger nucleases.Genetics
161, 1169-1175.
Bill, B. R., Petzold, A. M., Clark, K. J., Schimmenti, L. A. and Ekker, S. C.(2009). A primer for morpholino use in zebrafish.Zebrafish6, 69-77.
Blitz, I. L., Biesinger, J., Xie, X. and Cho, K. W. Y.(2013). Biallelic genome modification in F(0)Xenopus tropicalisembryos using the CRISPR/Cas system.
Genesis51, 827-834.
Blum, M., De Robertis, E. M., Wallingford, J. B. and Niehrs, C. (2015). Morpholinos: antisense and sensibility.Dev. Cell35, 145-149.
Boch, J., Scholze, H., Schornack, S., Landgraf, A., Hahn, S., Kay, S., Lahaye, T., Nickstadt, A. and Bonas, U.(2009). Breaking the code of DNA binding specificity of TAL-type III effectors.Science326, 1509-1512.
Bofill-De Ros, X. and Gu, S.(2016). Guidelines for the optimal design of miRNA-based shRNAs.Methods103, 157-166.
Braun, M. H., Steele, S. L., Ekker, M. and Perry, S. F.(2009). Nitrogen excretion in developing zebrafish (Danio rerio): a role for Rh proteins and urea transporters.
Am. J. Physiol. Ren. Physiol.296, F994-F1005.
Brenner, S.(1974). The DNA ofCaenorhabditis elegans.Genetics77, 95-104.
Cai, X., Zhang, D., Wang, J., Liu, X., Ouyang, G. and Xiao, W.(2018). Deletion of thefihgene encoding an inhibitor of hypoxia-inducible factors increases hypoxia tolerance in zebrafish.J. Biol. Chem.293, 15370-15380.
Carthew, R. W. and Sontheimer, E. J.(2009). Origins and mechanisms of miRNAs and siRNAs.Cell136, 642-655.
Chahine, S., Campos, A. and O’Donnell, M. J.(2012). Genetic knockdown of a single organic anion transporter alters the expression of functionally related genes in Malpighian tubules ofDrosophila melanogaster.J. Exp. Biol.215, 2601-2610.
Chang, N., Sun, C., Gao, L., Zhu, D., Xu, X., Zhu, X., Xiong, J.-W. and Xi, J. J.
(2013). Genome editing with RNA-guided Cas9 nuclease in zebrafish embryos.
Cell Res.23, 465-472.
Chasiotis, H., Ionescu, A., Misyura, L., Bui, P., Fazio, K., Wang, J., Patrick, M., Weihrauch, D. and Donini, A.(2016). An animal homolog of plant Mep/Amt transporters promotes ammonia excretion by the anal papillae of the disease vector mosquitoAedes aegypti.J. Exp. Biol.219, 1346-1355.
Chen, S., Reichert, S., Singh, C., Oikonomou, G., Rihel, J. and Prober, D. A.
(2017). Light-dependent regulation of sleep and wake states by prokineticin 2 in zebrafish.Neuron95, 153-168.e6.
Cho, S. W., Kim, S., Kim, Y., Kweon, J., Kim, H. S., Bae, S. and Kim, J.-S.(2014). Analysis of off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases.Genome Res.24, 132-141.
Choe, K. P. and Strange, K.(2008). Genome-wide RNAi screen and in vivo protein aggregation reporters identify degradation of damaged proteins as an essential hypertonic stress response.Am. J. Physiol. Cell Physiol.295, C1488-C1498.
Choi, S. H., Jee, B. Y., Lee, S. J., Cho, M. Y., Lee, S. J., Kim, J. W., Jeong, H. D. and Kim, K. H.(2013). Effects of RNA interference-mediated knock-down of hypoxia-inducible factor-α on respiratory burst activity of the Pacific oyster
Crassostrea gigashemocytes.Fish Shellfish Immunol.35, 476-479.
Christian, M., Cermak, T., Doyle, E. L., Schmidt, C., Zhang, F., Hummel, A., Bogdanove, A. J. and Voytas, D. F.(2010). Targeting DNA double-strand breaks with TAL effector nucleases.Genetics186, 757-761.
Ciruna, B., Weidinger, G., Knaut, H., Thisse, B., Thisse, C., Raz, E. and Schier, A. F. (2002). Production of maternal-zygotic mutant zebrafish by germ-line replacement.Proc. Natl. Acad. Sci. USA99, 14919-14924.
Cong, L., Ran, F. A., Cox, D., Lin, S., Barretto, R., Habib, N., Hsu, P. D., Wu, X., Jiang, W. and Marraffini, L. (2013). Multiplex genome engineering using CRISPR/Cas systems.Science339, 819-823.
Corey, D. R. and Abrams, J. M.(2001). Morpholino antisense oligonucleotides: tools for investigating vertebrate development.Genome Biol.2, reviews1015-r1.
Cote, F., Fligny, C., Bayard, E., Launay, J.-M., Gershon, M. D., Mallet, J. and Vodjdani, G. (2007). Maternal serotonin is crucial for murine embryonic development.Proc. Natl. Acad. Sci. USA104, 329-334.
Crombie, T. A., Tang, L., Choe, K. P. and Julian, D.(2016). Inhibition of the oxidative stress response by heat stress inCaenorhabditis elegans.J. Exp. Biol.
219, 2201-2211.
Cui, Z., Liu, Y., Wang, W., Wang, Q., Zhang, N., Lin, F., Wang, N., Shao, C., Dong, Z. and Li, Y.(2017). Genome editing reveals dmrt1 as an essential male sex-determining gene in Chinese tongue sole (Cynoglossus semilaevis).Sci. Rep.7, 42213.
Cullen, B. R.(2006). Enhancing and confirming the specificity of RNAi experiments.
Nat. Methods3, 677-681.
Davies, B., Brown, L. A., Cais, O., Watson, J., Clayton, A. J., Chang, V. T., Biggs, D., Preece, C., Hernandez-Pliego, P. and Krohn, J.(2017). A point mutation in the ion conduction pore of AMPA receptor GRIA3 causes dramatically perturbed sleep patterns as well as intellectual disability.Hum. Mol. Genet.26, 3869-3882.
de Angelis, M. H., Flaswinkel, H., Fuchs, H., Rathkolb, B., Soewarto, D., Heffner, S., Pargent, W., Wuensch, K., Jung, M., Reis, A. et al.(2000). Genome-wide, large-scale production of mutant mice by ENU mutagenesis.Nat. Genet.25, 1-4.
Doudna, J. A. Charpentier, E.(2014). The new frontier of genome engineering with CRISPR-Cas9.Science346, 1258096.
Doyon, Y., McCammon, J. M., Miller, J. C., Faraji, F., Ngo, C., Katibah, G. E., Amora, R., Hocking, T. D., Zhang, L. and Rebar, E. J.(2008). Heritable targeted gene disruption in zebrafish using designed zinc-finger nucleases. Nat.
Biotechnol.26, 702.
Driever, W., Solnica-Krezel, L., Schier, A. F., Neuhauss, S. C., Malicki, J., Stemple, D. L., Stainier, D. Y., Zwartkruis, F., Abdelilah, S., Rangini, Z. et al.
(1996). A genetic screen for mutations affecting embryogenesis in zebrafish.
Development.123:37-46., 37-46.
Durant, A. C., Chasiotis, H., Misyura, L. and Donini, A.(2017).Aedes aegypti Rhesus glycoproteins contribute to ammonia excretion by larval anal papillae.
J. Exp. Biol.220, 588-596.
Eisen, J. S. and Smith, J. C.(2008). Controlling morpholino experiments: don’t stop making antisense.Development135, 1735-1743.
Ekker, S. C.(2000). Morphants: a new systematic vertebrate functional genomics approach.Yeast17, 302-306.
Ekker, S. C. and Larson, J. D. (2001). Morphant technology in model developmental systems.Genesis30, 89-93.
El-Brolosy, M. A. and Stainier, D. Y. R. (2017). Genetic compensation: a phenomenon in search of mechanisms.PLoS Genet.13, 1-17.
Eve, A. M. J., Place, E. S. and Smith, J. C.(2017). Comparison of zebrafish
tmem88amutant and morpholino knockdown phenotypes.PLoS ONE12, 1-17.
Facchinello, N., Skobo, T., Meneghetti, G., Colletti, E., Dinarello, A., Tiso, N., Costa, R., Gioacchini, G., Carnevali, O. and Argenton, F.(2017).nr3c1null mutant zebrafish are viable and reveal DNA-binding-independent activities of the glucocorticoid receptor.Sci. Rep.7, 4371.
Faucherre, A., Kissa, K., Nargeot, J., Mangoni, M. E. and Jopling, C.(2014).
Piezo1plays a role in erythrocyte volume homeostasis.Haematologica99, 70-75.
Faucherre, A., Kissa, K., Nargeot, J., Mangoni, M. E. and Jopling, C.(2016). Comment on:“Homozygous knockout of thepiezo1gene in the zebrafish is not associated with anemia”.Haematologica101, e38.
Ferguson, D. P., Schmitt, E. E. and Lightfoot, J. T.(2013). Vivo-morpholinos induced transient knockdown of physical activity related proteins.PLoS ONE8, e61472.
Ferguson, D. P., Dangott, L. J. and Lightfoot, J. T.(2014a). Lessons learned from vivo-morpholinos: How to avoid vivo-morpholino toxicity. BioTechniques 56, 251-256.
Ferguson, D. P., Dangott, L. J., Schmitt, E. E., Vellers, H. L. and Lightfoot, J. T.
(2014b). Differential skeletal muscle proteome of high- and low-active mice.
J. Appl. Physiol.116, 1057-1067.
Figeac, F., Uzan, B., Faro, M., Chelali, N., Portha, B. and Movassat, J.(2010). Neonatal growth and regeneration ofβ-cells are regulated by the Wnt/β-catenin signaling in normal and diabetic rats.Am. J. Physiol. Endocrinol. Metab.298, E245-E256.
Fire, A., Xu, S. Q., Montgomery, M. K., Kostas, S. A., Driver, S. E. and Mello, C. C.(1998). Potent and specific genetic interference by double-stranded RNA in
Caenorhabditis elegans.Nature391, 806.
Flowers, G. P., Timberlake, A. T., McLean, K. C., Monaghan, J. R. and Crews, C. M.(2014). Highly efficient targeted mutagenesis in axolotl using Cas9 RNA-guided nuclease.Development141, 2165-2171.
Friedland, A. E., Tzur, Y. B., Esvelt, K. M., Colaiácovo, M. P., Church, G. M. and Calarco, J. A.(2013). Heritable genome editing inC. elegansvia a CRISPR-Cas9 system.Nat. Methods10, 741.
Fu, Y., Sander, J. D., Reyon, D., Cascio, V. M. and Joung, J. K.(2014). Improving CRISPR-Cas nuclease specificity using truncated guide RNAs.Nat. Biotechnol.
32, 279.
Gáliková, M., Diesner, M., Klepsatel, P., Hehlert, P., Xu, Y., Bickmeyer, I., Predel, R. and Kühnlein, R. P. (2015). Energy homeostasis control inDrosophila adipokinetic hormone mutants.Genetics201, 665-683.
Gao, J., Wang, G., Ma, S., Xie, X., Wu, X., Zhang, X., Wu, Y., Zhao, P. and Xia, Q.
(2015). CRISPR/Cas9-mediated targeted mutagenesis inNicotiana tabacum.
Plant Mol. Biol.87, 99-110.
Journal
of
Experimental
Gasiunas, G., Barrangou, R., Horvath, P. and Siksnys, V.(2012). Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria.Proc. Natl. Acad. Sci. USA109, E2579-E2586.
Gentsch, G. E., Spruce, T., Monteiro, R. S., Owens, N. D. L., Martin, S. R., Smith, J. C., Gentsch, G. E., Spruce, T., Monteiro, R. S., Owens, N. D. L. et al.(2018). Innate immune response and off-target mis-splicing are common morpholino-inuced side effects inXenopus.Dev. Cell44, 1-14.
Gerety, S. S. and Wilkinson, D. G.(2011). Morpholino artifacts provide pitfalls and reveal a novel role for pro-apoptotic genes in hindbrain boundary development.
Dev. Biol.350, 279-289.
Geurts, A. M., Cost, G. J., Freyvert, Y., Zeitler, B., Miller, J. C., Choi, V. M., Jenkins, S. S., Wood, A., Cui, X. and Meng, X.(2009). Knockout rats via embryo microinjection of zinc-finger nucleases.Science325, 433.
Gratz, S. J., Cummings, A. M., Nguyen, J. N., Hamm, D. C., Donohue, L. K., Harrison, M. M., Wildonger, J. and O’Connor-Giles, K. M.(2013). Genome engineering of Drosophila with the CRISPR RNA-guided Cas9 nuclease.
Genetics194, 1029-1035.
Guilinger, J. P., Thompson, D. B. and Liu, D. R.(2014). Fusion of catalytically inactive Cas9 to FokI nuclease improves the specificity of genome modification.
Nat. Biotechnol.32, 577.
Guschin, D. Y., Waite, A. J., Katibah, G. E., Miller, J. C., Holmes, M. C. and Rebar, E. J.(2010). A rapid and general assay for monitoring endogenous gene modification.Methods Mol. Biol.649, 247-256.
Haffter, P., Granato, M., Brand, M., Mullins, M. C., Hammerschmidt, M., Kane, D. A., Odenthal, J., van Eeden, F. J. M., Jiang, Y.-J., Heisenberg, C.-P. et al.
(1996). The identification of genes with unique and essential functions in the development of the zebrafish,Danio rerio.Development123, 1-36.
Hannon, G. J.(2002). RNA interference.Nature418, 244-251.
Heasman, J.(2002). Morpholino oligos: making sense of antisense?Dev. Biol.243, 209-214.
Heasman, J., Kofron, M. and Wylie, C. (2000). β-catenin signaling activity dissected in the earlyXenopusembryo: a novel antisense approach.Dev. Biol.
222, 124-134.
Homma, N., Harada, Y., Uchikawa, T., Kamei, Y. and Fukamachi, S.(2017). Protanopia (red blindness) in medaka: a simple system for producing color-blind fish and testing their spectral sensitivity.BMC Genet.18, 10.
Horn, E. R., El-Yamany, N. A. and Gradl, D.(2013). The vestibuloocular reflex of tadpoles (Xenopus laevis) after knock-down of the isthmus related transcription factor XTcf-4.J. Exp. Biol.733-741.
Horng, J.-L., Lin, L.-Y., Huang, C.-J., Katoh, F., Kaneko, T. and Hwang, P.-P.
(2007). Knockdown of V-ATPase subunit A (atp6v1a) impairs acid secretion and ion balance in zebrafish (Danio rerio).Am. J. Physiol. Regul. Integr. Comp.
Physiol.292, R2068-R2076.
Housden, B. E., Muhar, M., Gemberling, M., Gersbach, C. A., Stainier, D. Y. R., Seydoux, G., Mohr, S. E., Zuber, J. and Perrimon, N.(2017). Loss-of-function genetic tools for animal models: cross-species and cross-platform differences.
Nat. Rev. Genet.18, 24.
Hsu, P. D., Scott, D. A., Weinstein, J. A., Ran, F. A., Konermann, S., Agarwala, V., Li, Y., Fine, E. J., Wu, X., Shalem, O. et al.(2013). DNA targeting specificity of RNA-guided Cas9 nucleases.Nat. Biotechnol.31, 827-832.
Hua, Y., Wang, C., Huang, J. and Wang, K.(2017). A simple and efficient method for CRISPR/Cas9-induced mutant screening.J. Genet. Genomics44, 207-213.
Huang, P., Xiao, A., Zhou, M., Zhu, Z., Lin, S. and Zhang, B.(2011). Heritable gene targeting in zebrafish using customized TALENs.Nat. Biotechnol.29, 699.
Hwang, W. Y., Fu, Y., Reyon, D., Maeder, M. L., Tsai, S. Q., Sander, J. D., Peterson, R. T., Yeh, J. R. J. and Joung, J. K.(2013). Efficient genome editing in zebrafish using a CRISPR-Cas system.Nat. Biotechnol.31, 227.
Jackson, A. L. and Linsley, P. S.(2004). Noise amidst the silence: off-target effects of siRNAs?Trends Genet.20, 521-524.
Jackson, A. L., Bartz, S. R., Schelter, J., Kobayashi, S. V., Burchard, J., Mao, M., Li, B., Cavet, G. and Linsley, P. S.(2003). Expression profiling reveals off-target gene regulation by RNAi.Nat. Biotechnol.21, 635.
Jackson, A. L., Burchard, J., Schelter, J., Jackson, A. L., Burchard, J., Schelter, J., Chau, B. N., Cleary, M., Lim, L. E. E. and Linsley, P. S.(2006a). Widespread siRNA “off-target” transcript silencing mediated by seed region sequence complementarity.RNA12, 1179-1187.
Jackson, A. L., Burchard, J., Leake, D., Reynolds, A., Schelter, J., Guo, J., Johnson, J. M., Lim, L., Karpilow, J., Nichols, K. et al.(2006b). Position-specific chemical modification of siRNAs reduces“off-target”transcript silencing. RNA12, 1197-1205.
Jahangiri, L., Sharpe, M., Novikov, N., González-Rosa, J. M., Borikova, A., Nevis, K., Paffett-lugassy, N., Zhao, L., Adams, M., Guner-ataman, B. et al.
(2016). The AP-1 transcription factor component Fosl2 potentiates the rate of myocardial differentiation from the zebrafish second heart field.Development143, 113-122.
Jao, L.-E., Wente, S. R. and Chen, W.(2013). Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system.Proc. Natl. Acad. Sci. USA
110, 13904-13909.
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A. and Charpentier, E.
(2012). A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity.Science337, 816-821.
Jinek, M., East, A., Cheng, A., Lin, S., Ma, E. and Doudna, J.(2013). RNA-programmed genome editing in human cells.Elife2, e00471.
Joris, M., Schloesser, M., Baurain, D., Hanikenne, M., Muller, M. and Motte, P.
(2017). Number of inadvertent RNA targets for morpholino knockdown inDanio
reriois largely underestimated: Evidence from the study of Ser/Arg-rich splicing
factors.Nucleic Acids Res.45, 9547-9557.
Joung, J. K. and Sander, J. D.(2013). TALENs: a widely applicable technology for targeted genome editing.Nat. Rev. Mol. Cell Biol.14, 49.
Juntti, S. A., Hilliard, A. T., Kent, K. R., Kumar, A., Nguyen, A., Jimenez, M. A., Loveland, J. L., Mourrain, P. and Fernald, R. D.(2016). A neural basis for control of cichlid female reproductive behavior by prostaglandin F2α.Curr. Biol. 26, 943-949.
Katow, H., Suyemitsu, T., Ooka, S., Yaguchi, J., Jin-nai, T., Kuwahara, I., Katow, T., Yaguchi, S. and Abe, H.(2010). Development of a dopaminergic system in sea urchin embryos and larvae.J. Exp. Biol.213, 2808-2819.
Kennerdell, J. R. and Carthew, R. W.(1998). Use of dsRNA-mediated genetic interference to demonstrate thatfrizzledandfrizzled2 act in the wingless pathway. Cell95, 1017-1026.
Kim, Y.-G., Cha, J. and Chandrasegaran, S.(1996). Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain.Proc. Natl. Acad. Sci. USA93, 1156-1160.
Kishimoto, K., Washio, Y., Yoshiura, Y., Toyoda, A., Ueno, T., Fukuyama, H., Kato, K. and Kinoshita, M.(2018). Production of a breed of red sea bream
Pagrus majorwith an increase of skeletal muscle muss and reduced body length
by genome editing with CRISPR/Cas9.Aquaculture495, 415-427.
Kok, F. O., Shin, M., Ni, C.-W., Gupta, A., Grosse, A. S., van Impel, A., Kirchmaier, B. C., Peterson-Maduro, J., Kourkoulis, G., Male, I. et al.(2015). Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish.Dev. Cell32, 97-108.
Kolosov, D. and Kelly, S. P.(2017). Claudin-8d is a cortisol-responsive barrier protein in the gill epithelium of trout.J. Mol. Endocrinol.59, 299-310.
Kolosov, D., Donini, A. and Kelly, S. P. (2017). Claudin-31 contributes to corticosteroid-induced alterations in the barrier properties of the gill epithelium.
Mol. Cell. Endocrinol.439, 457-466.
Kumai, Y. and Perry, S. F.(2011). Ammonia excretion via Rhcg1 facilitates Na+
uptake in larval zebrafish,Danio rerio, in acidic water.Am. J. Physiol. Regul.
Integr. Comp. Physiol.301, R1517-R1528.
Labun, K., Montague, T. G., Gagnon, J. A., Thyme, S. B. and Valen, E.(2016). CHOPCHOP v2: a web tool for the next generation of CRISPR genome engineering.Nucleic Acids Res.44, W272-W276.
Lawson, N. D.(2016). Reverse genetics in zebrafish: mutants, morphants, and moving forward.Trends Cell Biol.26, 77-79.
Li, T. and Folkesson, H.(2006). RNA interference for a-ENaC inhibits rat lung fluid absorption in vivo.Am. J. Physiol. Cell. Mol. Physiol.95, 649-660.
Li, T., Huang, S., Jiang, W. Z., Wright, D., Spalding, M. H., Weeks, D. P. and Yang, B.(2010). TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain.Nucleic Acids Res.39, 359-372.
Li, J., Yue, Y., Dong, X., Jia, W., Li, K., Liang, D., Dong, Z., Wang, X., Nan, X., Zhang, Q. et al. (2015). Zebrafish foxc1a plays a crucial role in early somitogenesis by restricting the expression ofaldh1a2directly.J. Biol. Chem.
290, 10216-10228.
Li, R., Tian, J.-Z., Zhuang, C.-H., Zhang, Y.-C., Geng, X.-Y., Zhu, L.-N. and Sun, J.-S.(2016). CHHBP: a newly identified receptor of crustacean hyperglycemic hormone.J. Exp. Biol.219, 1259-1268.
Lim, Y., Bak, S. Y., Sung, K., Jeong, E., Lee, S. H., Kim, J.-S., Bae, S. and Kim, S. K.(2016). Structural roles of guide RNAs in the nuclease activity of Cas9 endonuclease.Nat. Commun.7, 13350.
Lin, X., Ruan, X., Anderson, M. G., McDowell, J. A., Kroeger, P. E., Fesik, S. W. and Shen, Y.(2005). siRNA-mediated off-target gene silencing triggered by a 7 nt complementation.Nucleic Acids Res.33, 4527-4535.
Liu, Q., Segal, D. J., Ghiara, J. B. and Barbas, C. F.(1997). Design of polydactyl zinc-finger proteins for unique addressing within complex genomes.Proc. Natl.
Acad. Sci. USA94, 5525-5530.
Liu, Y., Sepich, D. S. and Solnica-krezel, L.(2017). Stat3/Cdc25a-dependent cell proliferation promotes embryonic axis extension during zebrafish gastrulation.
PLoS Genet.13, e1006564.
Lu, Z.-C. and Wan, F.-H.(2011). Using double-stranded RNA to explore the role of heat shock protein genes in heat tolerance inBemisia tabaci(Gennadius).J. Exp.
Biol.214, 764-769.
Lupton, C., Sengupta, M., Cheng, R.-K., Chia, J., Thirumalai, V. and Jesuthasan, S. (2017). Loss of the habenula intrinsic neuromodulator kisspeptin1 affects learning in larval zebrafish. eNeuro 4, ENEURO-0326-16.2017.
Mahfouz, M. M., Li, L., Shamimuzzaman, M., Wibowo, A., Fang, X. and Zhu, J.-K.
(2011). De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-strand breaks.Proc.
Natl. Acad. Sci. USA108, 2623-2628.