JOURNRL
OF
MNGE MNVGEMENT
Published bimonthly-January, March, May,July, September, November-by the Society for Range Management 2120 South Birch Street Denver, Colorado 80222
Copyright @ 1976 by the Society for Range Management Managing Editor
DAVID A. SMITH
2120 South Birch Street Denver, Colo. 80222 Editor
ELBERT H. REID
624 South Shields Street Fort Collins, Colo. 80521 Book Review Editor PAT 0. CURRIE
Rocky Mountain Forest and Range Experiment Station 240 W. Prospect Street Fort Collins, Colo. 80521 Copy Editor
PATRICIA G. SMITH 2120 South Birch Street Denver, Colo. 80222 Editorial Board 1974-76
PHILLIP L. SIMS, Fort Collins, Colo. REX D. PIEPER, Las Cruces, N. Mex. MINORU HIRONAKA, Moscow, Ida. MARK R. KILCHER, Swift Current, Sask. 1975-77
JAMES B. NEWMAN, Lincoln, Neb. WALTER F. MUEGGLER, Logan, Utah DARRELL N. UECKERT, Lubbock, Tex. 1976-78
JOHN MALECHEK, Logan, Utah C. J. SCIFRES, College Station, Tex. CLIFFORD E. LEWIS, Marianna, Fla. RAYMOND EVANS, Reno, Nev.
INDIVIDUAL SUBSCRIPTION IS by membershrp rn the Socretv for Range Management
LIBRARY or other INSTITUTIONAL SUBSCRIP TIONS, on a calendar year basis, are 822 00 postpaid to all countraes Payments from outslde the Unlted States should be remltted I” US dollars by lnternatlonal money order or draft on a New York bank.
CHANGE OF ADDRESS notrces should be sent to the Managing Edrtor, 2120 S. Brrch St , Denver, CO 80222, no later than the frrst day of the month of Issue Copres lost due to a change of address cannot be replaced unless adequate notrce IS grven. To assure unrnterrupted ser vice. provrde your local postmaster wrth a Change of AddressOrder (POD Form 35751, rndrcatrng thereon to guarantee forwardrng postage for second class marl. BUSINESS CORRESPONDENCE, concernrng subscrrp trons, advertrsing, reprcnts, back issues, and related matters, should be addressed to the Mdnagrng Editor, 2120s. Btrch St., Denver, CO 80222.
EDITORIAL CORRESPONDENCE, concerning manu- scrrpts or other edrtorral matters, should be addressed to the Edrtor, 624 S. Shtelds St., Fort Colfrns, CO 80521. INSTRUCTtONS FOR AUTHORS appear each year cn the March issue, copres of these rnstructions are available from the editor.
SECOND CLASS POSTAGE patd at Denver, Colorado.
TABLE OF CONTENTS: Vol. 29, No. 4, July 1976 ARTICLES 268 272 278 281 284 286 290 294 299 301 304 306 309 311 313 316 320 322 326 328 333 338 342
Cultural Energy Expended in Range Meat and Fiber Production by C. Wayne Cook
The Effects of Weather Modification on Northern Great Plains Grasslands: A Preliminary Assessment by David A. Perry
The Abortifacient and Toxic Effects of Big Sagebrush (Artemisiu tridentata) and Juniper (Juniperus osteosperma) on Domestic Sheep by A. Earl Johnson, Lynn F. James, and Juan Spillett
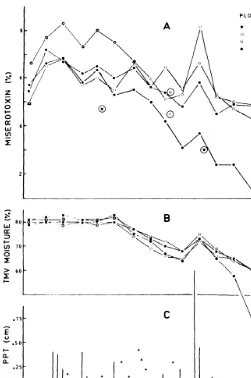
The Effect of Rainfall on Columbia Milkvetch Toxicity by W. Majak, R. J. Williams, A. L. van Ryswyk, and B. M. Brooke
Toxicity of Bassiu hyssopifolia to Sheep by L. F. James, M. C. Williams, and A. T. Bleak
Twenty Years of Changes in Grass Production Following Mesquite Control and Re- seeding by Dwight R. Cable
Botanical Composition of Eland and Goat Diets on an Acacia-grassland Community in Kenya by John C. Nge’the and Thadis W. Box
Effect of Prescribed Burning on Sediment, Water Yield, and Water Quality from Dozed Juniper Lands in Central Texas by Henry A. Wright, Francis M. Churchill, and W. Clark Stevens
Grazing and Debris Burning on Pinyon-Juniper Sites-Some Chemical Water Quality Implications by John C. Buckhouse and Gerald F. Gifford
Responses of Herbage, Pines, and Hardwoods to Early and Delayed Burning in a Young Slash Pine Plantation by Harold E. Grelen
Effects of Cattle Grazing and Wildfire on Soil-Dwelling Nematodes of the Shrub-Steppe Ecosystem by J. D. Smolik and L. E. Rogers
Range Burning and Fertilizing Related to Nutritive Value of Bluestem Grass by Leland J. Allen, Leniel H. Harbers, Robert R. Schalles, Clenton E. Owensby, and Ed. F. Smith
Impact of Wildfire on Three Perennial Grasses in South-Central Washington by D. W. Uresk, J. F. Cline, and W. H. Rickard
Clipping Effects on Dry Matter Yields and Survival of Basin Wildrye by L. J. Perry, Jr., and S. R. Chapman
Soil Moisture Response to Several Levels of Foliage Removal on Two Utah Ranges by John C. Buckhouse and George B. Coltharp
Bahiagrass Regrowth and Physiological Aging by E. V. S. B. Sampaio, E. R. Beaty, and D. A. Ashley
Variable Germination Response to Temperature for Different Sources of Winterfat Seed
by J. L. Moyer and R. L. Lang
Estimating Potential Downy Brome Competition after Wildfires by James A. Young, Raymond A. Evans, and Ronald A. Weaver
Expressing the Competitive Relationship between Wyoming Big Sagebrush and Crested Wheatgrass by L. R. Rittenhouse and F. A. Sneva
An Evaluation of Barrel Medic (Medicago truncatula) as an Introduced Pasture Legume for Marginal Cropping Areas of Southeastern Australia by D. L. Michalk and J. A. Beale
Influence of Date of Planting on Emergence of Cultivars of Trifolium hirtum All. and T. subterraneum L. by Raymond A. Evans, Burgess L. Kay, and James A. Young
Subsurface Herbicide Applicator for Brush Control by R. W. Bovey, T. 0. Flynt, R. E. Meyer, J. R. Baur, and T. E. Riley
Remote Sensing for Optimum Herbicide Application Date for Rabbitbrush by James A. Young, Raymond A. Evans, and Paul T. Tueller
TECHNICAL NOTES
344 Caloric Content of Rocky Mountain Subalpine and Alpine Plants by Douglas C. Andersen and Kenneth B. Armitage
346 Perennial Grasses Respond Inconsistently to Alternate Year Seasonal Rest by S. Clark Martin and Donald E. Ward
347 Foods of Free-Roaming Horses in Southern New Mexico by Richard M. Hansen 348 A Supporting Device for Use with Stepwise Thermal Sensors by George E. Probasco,
Ardell J. Bjugstad, and Roy W. Pierce
349 Fast Filter for In Vitro Studies by Donald R. Dietz and Harold E. Messner
BOOK REVIEWS
351 Readings in Natural Resource Economics edited by John E. Reynolds, J. Martin Redfern, and Robert N. Shulstad; Africa: the Devastated Continent? by A. de Vos; The State of
Food and Agriculture 1974 by Food and Agricultur Organization of the United Nations
Cultural Energy Expended in Range Meat and
Fiber Production
C. WAYNE COOK
Highlight: Range livestock production requires more cultur- energy in heat when steam is used to produce power to turn al energy than commonly believed. However the cultural the -turbines. This does not include the cultural energy energy expended for range meat and fiber is considerably less
than that required in confined fattening procedures. Comple-
required to mine the coal and transport it to the generating sites or the loss in transmission lines. In the Central Plains area menting rangelands with dryland forages offers great promise
in decreasing the cost of fossil fuel to produce a pound of red it was determined that about 80% of the electricity comes meat for human consumption, compared to feedlot fattening. from steam generators and 20% from hydroelectric sources. Sorghum produced on summer-fallowed ground for grazing
Oil and gas in the United States are finite resources and required 495.7 Meal per acre and feeds used in supplements predictions are that there are only a SO-year supply of gas and and rations varied from 350 to 500 kcal per pound (Cook et a 75-year supply of oil at the present rate of usage. It is true al., 1976).
that we have several hundred years’ supply of coal and a rather
vast energy reserve in oil shale and nuclear material, but they Range Sheep Production in the Intermountain Great Basin too are finite and expensive to develop as substitutes for gas The sheep ranches included in the study were typical of and oil. Therefore the public is becoming conscious of the use many operations in Utah, Nevada, and southern Idaho, where of fossil fuels, whether it be for pleasure or for the production sheep graze desert ranges during the winter from about of food. In the latter case, a close correlation will develop November 1 until about April 1, after which they are trailed between the cost of a product and the expenditure of cultural
energy to produce it.
It is of interest to evaluate the cultural energy expended in the production of food and particularly meat because meat comes from the second feeding level in the food chain and therefore requires more subsidized energy in the conversion of solar energy than do plant foods.
Measures of Cultural Energy
Cultural or subsidized energy is the energy other than solar energy needed to produce food and fiber. Cultural energy includes labor, manufacture of machinery, transportation, tillage, production of fertilizers and herbicides, processing material, and finally marketing the product (Cook et al., 1976).
Calculations of cultural energy input for labor were 2,500 kcal per hour, for depreciation and maintenance of large machinery were 45,000 kcal per hour of operation, gasoline per gallon contains 3 1,056 kcal, transportation requires 4 kcal per pound per mile (Cook et al., 1976). Electricity was calculated to be about 860 kcal per kilowatt hour (Table 1). Electricity was calculated as being only 40% efficient because it requires 2.5 more BTU’s of coal to produce the equivalent kilowatt hours of electricity. This is a result of the loss of
Author is head, Department of Range Science, Colorado State University, Fort Collins, 80523.
Manuscript received January 15, 1976.
onto foothill range where they lamb on crested wheatgrass and/or native grass-sagebrush types. About the middle of June they are moved into mountain brush types and subsequently into higher mountain zones.
In this study all lambs were black-face crosses. All cultural energy expenditures were made on the basis of a 1,800 mature ewe herd with 200 replacements, 36 bucks, and 3 to 4 horses. From these operations about 70% of the lambs were sold as grass-fat choice lambs weighing about 90 lb with a 50.9 dressing percentage. The dressed carcass was 16.2% protein and 23.4% fat. The remaining 30% of the lambs weighed 75 lb and sold as feeders which dressed 47.5% with 17.1% protein and 18.8% fat.
All carcass protein was considered to possess 5.7 kcal of gross energy per gram and all fat was calculated as having 9.4 kcal per gram.
The sheep, while on the desert ranges, were supplemented from December 15 until March 15 at the rate of one-half pound per head every other day. The supplement consisted of 45% alfalfa, 25% barley, and 30% soybean meal. The cultural energy in the alfalfa, barley, and soybean meal was 272 kcal, 411 kcal, and 658 kcal per pound of material respectively. The mixture was fed in pelleted form and was purchased at a feed mill and transported to the desert range about 48 miles away. It was calculated that 2.24 kcal of cultural energy was expended for each kilocalorie in dressed carcass meat and 1.54 Meal of cultural energy was used for each pound of live weight of lamb produced (Table 2).
There were 130,410 lb of choice lamb, which sold for 44 & a pound, totaling $57,380.00; 46,575 lb of feeder lamb, which sold for 42 6 for a total gross income of $19,561 .OO; and 20,000 lb of wool, which brought 72 b or a total of $14,400.00. Thus 84% of the sales came from lambs and 16% from wool. Therefore 16% of the total Meal of cultural energy (16 X 271,705 Meal) or 43,473 Meal can be assessed to wool production. The Meal of cultural energy for shearing was calculated as follows: 184 man hours amounting to 460 Meal, machinery totaling 4,259 Meal, the transportation of wool to market amounted to 14,400 Meal, and finally the scouring using 3,600 Meal. The total expenditure of cultural energy for shearing, handling, and processing wool was 22,7 19 Meal. Thus the addition of 43,473 Meal of cultural energy resulting from 16% of the general operational energy costs and the 22,719 Meal expended in actual shearing, transportation, and scouring of the wool would be 66,192 Meal for 8,000 lb of clean wool or 8.27 Meal/lb of scoured wool.
In a recent report by the National Cotton Council of America (Gatewood, 1973) it was estimated that cultural energy consumed in a pound of cotton lint was 3.22 Meal and the cultural energy cost for noncellulosic and cellulosic synthetic fibers per pound was 15.17 and 20.38 Meal, respectively. The noncellulosic fiber included polyester, nylon, and acrylic material, which are petro-chemicals or derived from petroleum or natural gas.
Cotton has a significant advantage in the amount of cultural energy required for fiber production since the cotton plant not only produces fiber but also produces cotton seed oil and cotton seed meal. Wool, like cotton, is a by-product or a dual product of the sheep industry and requires considerably less cultural energy than synthetic fibers.
Range Cattle Production in the Intermountain Great Basin
The 250-head cattle ranches that were analyzed in the
Intermountain Great Basin area operate somewhat similarly to sheep ranches of the region. Typical cow-calf operations throughout Utah, Nevada, and southern Idaho, like sheep operations, run on desert ranges during the winter from about November 1 to April 1 and are grazed on crested wheatgrass and/or native range during calving and until about the third week in June, when they move to mountain ranges until about October 1.
The supplement for cattle consisted of 30% alfalfa, 20% barley, and 50% soybean meal. This was fed to cattle in pelleted form for 90 days at the rate of 2 lb/day per animal. The supplement was hauled about 48 miles from the feed mill to the range where it was fed.
The percent calf crop was 93% and all calves were Hereford X Angus crosses, which sold at weaning time as good grade grass-fat calves weighing 405 lb. They dressed at 57.9% with the carcass containing 11.2% fat and 16.6% protein that totaled 2 12,437 kcal.
The expenditure of cultural energy per pound of weaning weight was 2.15 Meal; the ratio of cultural energy used per increment of food energy produced in the dressed carcass was 4.09 (Table 2).
Range Cattle Production in the Central Plains
In the central plains states of Nebraska, Kansas, Oklahoma, northern New Mexico, and eastern Colorado, a typical cow-calf operation remains on the range all or most of the year with only a few pounds of supplement fed during the winter months. Calves are dropped in mid-April generally. Cows, during calving, continue to receive a supplement until green growth starts on the ranges, which occurs during early May. The cow-calf unit is grazed on rangelands until about the middle of October, when the calves are weaned and sold.
In some cases forage sorghums are planted for summer grazing from about July 15 until weaning time. This is referred
Table 2. Average output of meat and energy compared to cultural energy input from various range sheep and cattle ranches and typical fefdot operations in the western range area.
Central
Intermountain Great Basin Central Great Plains Plains
Range sheep Range cows Range-sorghum Range cows Range-sorghum 15,000 2,000 female 250 female 250 cow 250 female 400 steer feedlot
herd herd operation herd operation operation
Market weight or gain (lb) 85.5’ 4052 445 3 414 396 586
Calf or lamb crop (%) 103.5 93 93 92 984 99
Total energy in carcass (Meal) 121,382 49,498 54,423 50,578 138,138 13,186,800
No. marketed 2,070 233 233 230 392 14,85 0
Live weight marketed (lb) 176,985 94,365 103,685 95,220 155,232 8,702,100
Cultural energy/food energy 2.24 4.09 5.25 4.58 2.77 5.33
Meal cultural energy/lb of live weight 1.54 2.15 2.76 2.44 2.47 8.07
’ 70% sold as choice fat lambs at 90 lb and 30% sold as feed lambs at 75 lb. ‘All sold as good grass fat calves.
3 70% sold as choice grass fat calves.
‘Percent death loss was 2%, therefore 98% was marketed, all sold as good grade grass fat steers weighing 810 lb.
’ Percent death loss was l%, therefore 99% was marketed, 70% sold as choice and 30% as good grade fed beef weighing an average of 1,000 lb.
to as a range-sorghum operation for producing grass-fat weaner calves that sell as choice grade feeder calves and weigh about 445 lb. For practical purposes the dressing percentage and the percent fat and protein in the carcass were considered the same as for the calves produced to choice grass-fat feeders in the Intermountain Great Basin area discussed previously.
The ratio of cultural energy used to produce an increment of food energy in meat was 5.25 for the cow-calf units that grazed range in conjunction with forage sorghum pastures for fat weaner calf production and 4.58 for the typical cow-calf range operation in the Central Plains region. The range- sorghum grazing system expended 2.76 Meal of cultural energy per pound of weaner calf compared to 2.44 Meal/lb of calf gain for the conventional range cow-calf operation (Table 2). The development of the forage sorghum pastures increased the expenditure of cultural energy because of the tillage and plantings of these pastures.
In all cases winter supplement was fed to the breeding cow herd from about the middle of December until about May 1 at the rate of 2 lb per head per day. The supplement used in the calculations consisted of 60% alfalfa, 20% soybean meal, and 20% milo, which averaged 348 kcal of cultural energy per pound of pelleted feed mixture.
The weaning weight from the range-sorghum grazing system produced calves weighing 445 lb at weaning about October 1.5; the conventional range cow-calf operation produced weaner calves weighing 414 lb. All of the weaner calves in the range-sorghum feeding system were graded as choice grass fat feeder calves, whereas only 70% of the calves produced solely from range forage sold as choice.
Steer Operations in the Central Plains
Many ranchers of the western range region have shifted from a cow-calf operation to an enterprise of growing steers. These animals are purchased as weaner calves and sold in the spring and summer as feeders. However, more and more operators are fattening such steers on mountain summer ranges or summer-fallowed forage sorghum pastures in the Central Great Plains area. If properly managed, the steers can be sold from these systems weighing over 800 lb at 17 to 18 months of age and grading as good for block meat. Such meat is now marketed by many chain stores under various trade names.
Experiments in Colorado showed that these steers gained from 414 lb as weaners to 8 10 lb when sold from the forage sorghum pastures in late summer. These 800-lb steers dress at 58.0% with 20.1% fat and 14.4% protein in the carcass. Again it was calculated that fat contained 9.4 kcal/g and protein contained 5.7 kcal/g.
In this study range steers were fed 3 lb of supplement per day from December 15 to May 15. The pelleted supplement contained 60% alfalfa, 20% soybean meal, and 20% barley, which had a cultural energy cost of 348 kcal/lb.
From the 400~steer operation analyzed it was calculated that 2.77 kcal of cultural energy were required for each kilocalorie of dressed meat produced and 2.47 Meal of cultural energy for each pound of steer gain (Table 2).
Feedlot Operations
A typical 15,000 feedlot capacity operation in the plains area near Denver, Colorado, was analyzed in this study. The animals fed consisted of 62% steers and 38% heifers. They were fed for 255 days, starting with an average weight of 414
lb and finishing at 1,000 lb. Thus the overall daily gain was 2.3 lb/day.
The ration the first 122 days was about 60% roughage, consisting of corn silage, chopped alfalfa, and beet pulp along with 40% dry rolled corn and soybean meal. During the next 133 days the ration was about 65% concentrates, consisting of dry rolled corn and soybean meal, along with 30% corn silage and 5% molasses. All silage was raised near the feeding operation and 40% of the corn was raised on the owned land. The remainder of the feed was hauled onto the farm feedlot from feed mills 15 miles away.
The average cultural energy for the ration during the first 122 days was 389 kcal of cultural energy per pound of feed and for the ration fed during the next 133 days was 495 kcal of cultural energy per pound of feed. The average for the entire 255-day feeding period was 444 kcal/lb of feed. The average daily intake per animal over the 255-day feeding period was 35 lb. During the early feeding period, daily consumption was 22 lb but was as high as 39 lb during the latter part of the feeding period.
The heifers dressed at 60.5% and the steers dressed at 62.5%, which was a weighted average of 61.7% for the 14,850 head marketed. Approximately 79% graded choice and 21% graded good.
It was calculated that an average 1 ,OOO-lb animal dressed at 62% with a 4% shrink and the carcass containing 35% fat and
11.8% protein, which yielded 1,108 Meal of energy per dressed carcass. The dressed carcass of the 414-lb weaner range calf had 220 Meal; therefore the 255-day feeding period produced 888 Meal per carcass from the 586 pounds gained per animal. With a 1% death loss the total Meal from the dressed carcass was 13,186,800 for 14,850 animals marketed (Table 2).
Under a fattening program under confinement feeding, 5.33 kcal of cultural energy were required to produce a kilocalorie of dressed carcass meat. Each pound gained required 8.07 Meal of cultural energy (Table 2). These figures were considerably higher than those for a combination of range and forage- sorghum pastures, which were 2.77 and 2.47 Meal of cultural energy for a Meal of meat and per pound of gain, respectively.
Complete Systems from Conception to Marketing
The total expenditure of cultural energy including the cow-calf and ewe-lamb operations until animals reached various marketable weights and ages is shown in Table 3. Sheep are considerably more efficient than cattle. This is a result of multiple births, faster growth rate, and ability to fatten to choice grade on range forage without grain. A ewe produces almost 62% of her weight as weaner lamb and a cow produces only about 45% of her weight as weaner calf in the same period. The cow-calf unit can be most competitive with the ewe-lamb unit in the production of red meat from forage by finishing steer calves to good grade on sorghum pastures.
The expenditure of cultural energy to produce a choice grade beef from conception to a 1 ,OOO-lb animal was of course the most inefficient in the use of cultural energy because of the use of harvested roughage and grain.
In the present study it required 5.74 Meal of cultural energy to produce 1 lb of fattened choice grade beef. Comparable figures by Lockeretz (1975) showed that 4.54 Meal per pound of beef from birth to feedlot fattening was required. In the same study for use of irrigated pastures along with feedlot finishing in Oklahoma, 11.6 Meal of cultural energy were required to produce a pound of finished beef.
Table 3. Average production of live weight along with expenditure of cultural energy per increment of energy produced and per pound of live weight gain for various systems of producing marketable lamb and beef on an individual basis at the source of production.
Weaner Steer3
Lamb’ calf2 production Feedlot production production on forage fattening
Weight produced (lb)
Energy in dressed carcass (Meal) Cultural energy
expended (Meal) Cultural energy
in/energy out Cultural energy in/lb of live weight (Meal)
85.5 414 810 1000
58.6 220 527 1108
131.3 1010 1987 5740
2.24 4.5 8 3.47 5.18
1.54 2.44 2.45 5.74
’ Ewes were white-face crosses weighing an average of 139 lb.
‘Cows were good grade Hereford cows weighing an average of 9 14 lb. 3Calculated for an animal including the cow-calf operation until
finished either on forage or in the feedlot.
Summary
The sheep industry has the advantage of producing choice grade meat directly off the range at a cost of only 2.24 kcal of cultural energy per kilocalorie of dressed carcass compared to cattle which required approximately 4.5 kcal per kilocalorie of dressed meat for weaner calves.
By using range along with forage-sorghum pastures, a good grade animal that was raised from a calf to a steer weighing 810 lb can be produced for an expenditure of about 3.5 kcal of cultural energy for each kilocalorie of dressed carcass and a choice weaner calf weighing 445 lb can be produced on range and forage sorghums for a cost of 5.25 kcal of cultural energy per kilocalorie of dressed meat. By intensive management of a cow-calf operation in the Intermountain Great Basin area, where crested wheatgrass is used for early spring grazing and good mountain range is used from June 15 to September 15, a
kilocalorie of dressed choice weaner-calf meat can be produced at a cost of 4.09 kcal of cultural energy.
Thus it appears that the lowest expenditure of cultural energy per kilocalorie of table meat is from lamb and meat from steers produced to good grade largely from range and forage-sorghum pastures.
The expenditure of cultural energy per pound of live weight gain of marketable animals was only 1.54 kcal/lb of gain for a ewe-lamb operation and 2.15 kcal/lb of gain for a cow-calf operation in the Intermountain Great Basin area, compared to 2.76 and 2.44 kcal/lb of gain in the Central Great Plains where forage sorghums are used for summer grazing and the conventional range operation for weaner calf production, respectively.
The steer operation that starts from birth and makes use of range and forage sorghums to a weight of 8 10 lb required 2.45 kcal of cultural energy per pound of body gain. Feedlot operations that start from birth and feed the weaner calf for 255 days to a final weight of 1,000 lb required 5.74 kcal of cultural energy per pound of live weight gain.
The relative cost of cultural energy per pound of body gain under the various livestock operations follows the same ranking as the kilocalories of cultural energy expended per kilocalorie of dressed meat. A ewe-lamb operation is most efficient, with a steer operation next. These are followed by the conventional range cow-calf operation, the range-sorghum pasture grazing system, and finally the feedlot fattening procedure.
Literature Cited
Cook, C. Wayne, A. H. Denham, E. T. Bartlett, and R. Dennis Child. 1976. Efficiency of converting nutrients and cultural energy in various feeding and grazing systems. J. Range Manage. 29:186-191. Gatewood, L. B., Jr. 1973. The energy crisis: can cotton help meet it?
Nat. Cotton Count. Amer. Jan. Rep.
THESIS: THE UNIVERSITY OF ARIZONA
Effect of Mesquite Control on Small Game Populations, by Dale P. McCormick, PhD, Range Sciences. 1975.
Mesquite has invaded extensive portions of the desert grass- land and where it has become a dominant plant, livestock forage has declined. As a result, range managers have often con- demned mesquite and advocated its control.
This study was conducted to determine the effect of mesquite control on the relative abundance of seven species of small game animals inhabiting the desert grassland.
To meet the study objectives, comparisons of ground cover and small game abundances were made between a mesquite- controlled and a mesquite-invaded pasture. Both areas, located on the Santa Rita Experimental Range near Tucson, Arizona, supported relatively good stands of native perennial grasses and forbs overtopped by varying degrees of woody plants.
While control of medium dense stands of mesquite ( 1% 100 trees per acre) did not significantly increase total perennial grass ground cover over a IO-year period, it decreased use by moum- ing and whitewinged doves, Gambel’s and scaled quail, and desert cottontail rabbits. Antelope and blacktailed jackrabbit abundances were relatively unaffected. Therefore, ranges in this condition should be maintained by controlled grazing for the benefit of both livestock and wildlife.
Mesquite control is recommended only when the density exceeds 100 trees per acre on upland sites. Small dense stands ( 100-335 trees per acre) of mesquite occurring in the drainages should be preserved.
The Effects of Weather Modification on North-
ern Great Plains Grasslands: A Preliminary
Assessment
DAVID A. PERRY
Highlight: Possible effects of weather modification on Northern Great Plains Grasslands are examined using published reports on community-water relations. It is concluded that (1) long-term incremental forage production will be governed by the effect of added water on nutrient cycling rates; (2) community composition will change, but the nature of the change will depend on the timing of added precipitation; and (3) increased forage in the absence of increased nitrogen may have a neutral or negative effect on livestock weight gains.
The use of cloud seeding to increase growing season rainfall is receiving a great deal of attention in the semiarid West. In the Northern Great Plains, both North and South Dakota are seeding clouds on a large scale operational basis, and the Bureau of Reclamation recently initiated a major experimental program in Montana. Not only is purposeful change in the region’s weather a growing possibility, but inadvertent weather modification will likely emerge as one of the probable spinoffs from increasing industrialization in the area.
In cultivated crops, which are relatively simple ecological systems, and in which man has a large number of management options, the results of changing weather can be anticipated with fair (though not exact) certainty (cf. Caprio and Williams,
1973). The complexity of the rangeland ecosystem, however, suggests that its response to added water will not be so easily predicted.
This paper reviews, for the Northern Great Plains, the literature on the relation between grasslands and water. The objective is an estimation of the effect added water will have on these range ecosystems. Based on this literature, I will deal
During the research the author was range ecologist. Montana Department of Natural Resources and Conservation; I-Iigh Plains Program, Miles City. At present he is research forester, U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, Ogden, Utah 84401, stationed in Bozeman, Mont., at the Forestry Sciences Laboratory, maintained in cooperation with Montana State University.
This work was funded by the Atmospheric Water Resources Division, U.S. Bureau of Reclamation, and Intermountain Forest and Range Exp. Sta., U.S. Dep. Agr., Forest Service. The author wishes to thank Don Ryerson, Kris Aase, Frances Siddoway, Ross Wight, Al Black, Earl Neff, Walt Mueggler, Arlin Super, Tad Weaver, Wyman Schmidt I and Joe Basile for reading and commenting on the manuscrrpt.
Manuscript received July 29, 1975.
with four questions: (1) What is the effect of increased rainfall on primary and secondary productivity? (2) What factors limit the response of grasslands to added water, and how will they be affected by increased rainfall? (3) How do different species respond to added water with various seasons of application, and how will the interaction of species response and season- ality affect the composition of the range? (4) How will changes in the weather affect community resistance to drought, plant and animal pathogens, and soil loss associated with runoff?
Effect of Added Water on Primary Productivity
Primary ,productivity of grasslands has been extensively studied in relation to increases in soil water. Two experimental techniques have been most often used: (1) comparison of rainfall and forage production using regression and correlation, and (2) irrigation, either by water spreading or sprinkling. Results have been mixed, making it difficult to generalize about rangeland response to added water.
Because regression techniques simulate plant response to increments in natural rainfall, they might be expected to give the most realistic estimate of the effect of weather modifi- cation on forage production. A number of studies have been conducted on northern grasslands. These have produced variable results, with differences in yield attributable to growing season rainfall ranging from 84% to less than 25% (Rogler and Haas, 1947; Smoliak, 1956; Dahl, 1963; Rauzi, 1964; Noller, 1968; Mueggler, 1972; Ballard and Ryerson, 1973). Where rainfall and forage production are significantly related, estimates of precipitation use efficiency have varied from approximately 50 lb dry forage/acre-inch of available water (Thomas and Osenbrug, 1964) to 110 lb dry forage/ acre-inch of water (Wight and Black, 1972).
The differences seen in these studies probably represent both experimental noise and biological variation. For example, if rain gauges are located separately from production sampling points, significant errors may enter into the analysis. On the other hand, the capacity to respond to added water certainly varies with time, space, and community type. It is the degree of this latter variation which is of interest.
Irrigation studies, although normally adding more water than might be expected from cloud seeding, have the advantage of being controlled experiments. They therefore presumably minimize experimental noise and allow a more accurate estimate of the true variability in the response of rangeland to additions of water. A number of irrigation experiments have produced significant results. Two water spreading studies in southeastern Montana resulted in grass forage increases of approximately 300% (Monson and Queensberry, 1958; Branson, 1956). Sprinkling in sufficient amounts to double growing season rainfall resulted in yield increases of 100% to 200% in western Montana (Klages and Ryerson, 1965) and 50% in North Dakota (Smika et al., 1965). Lauenroth and Sims (1973) found that yields of shortgrass prairie in Colorado were increased approximately
100% when soil water stress at 10 cm was kept above -0.8 bars.
However, an impressive array of experiments have shown little or no increased production resulting from irrigation. Cosper and Thomas (1961) found yield increases of only 16% on a water-spreading study in South Dakota. In New Mexico, water spreading on sandy sites did not affect vegetative cover (Valentine, 1947). Weaver (1975) found that additions of water ranging from 140% of normal rainfall to maintaining a constantly wet soil increased production of Idaho fescue
(Festuca idahoensis) meadows slightly when water was added
in the spring, but not at all for summer additions. Bleak et al. (1974) compared yields on Utah grasslands receiving 150% and 67% of normal rainfall, and found no difference. Pettit and Fagan (1974) irrigated buffalograss (Buchloe dactyloides) in combination with various nitrogen treatments, and found lower yields in irrigated than in nonirrigated plots in all but one nitrogen treatment. In two studies of the effect of irrigation on below-ground biomass, no increased root growth was found as a result of added water (Lauenroth and Sims, 1973; Garwood, 1967). Garwood attributed this to a faster rate of decay of dead roots.
These results indicate that the response of grasslands to added water is complex and highly variable. An evaluation of the role which weather modification will play on’ western rangelands requires an understanding of the factors affecting this variability. Several of these are discussed below. They are: (1) nutrient limitations, (2) pattern of water application, and (3) genetic limitations.
Factors Affecting the Grassland Response to Water
Nutrients
Available forms of nutrients, especially nitrogen (N) and phosphorus (P), are commonly in short supply in grasslands (Harmsen and Van Schreven, 1955). Range fertilization often results in dramatic increases in water use efficiency and forage production (Smika et al., 1965; Klages and Ryerson, 1965; Wight and Black, 1972; Lauenroth and Sims, 1973), and a number of investigators have concluded that N is more limiting than water on Northern Great Plains rangelands (White and Moore, 1972; Wight and Black, 1972; Lauenroth and Sims, 1973). Even where N isn’t the most limiting factor, it may quickly become so when water availability is increased. Thus, rather than being a linear relation, forage yield as a function of water availability may asymptotically approach some maxi- mum determined by available N (cf. Van Dyne, 1975). This would perhaps explain some cases of poor correlation between rainfall and forage production.
The effect which added precipitation has on nutrient availability is directly related to its effect on microbial activity. Soil CO* evolution and/or nitrification are inhibited at 0 bars soil water tension, rise quickly at - .15 to -.50 bars, and then decline with further drying, although the decline may be slow and significant activity may continue at tensions as low as -15 bars (Parker and Larson, 1962; Justice and Smith, 1962; Miller and Johnson, 1964; Wiant, 1967; Nyhan and Doxtader, 1974). Koepf (1952) reported that the optimum water content for the activity of soil bacteria was 60-70% of field capacity.
If microbial activity is largely confined to a certain range in soil water tension, nutrient release should be correlated with the length of time soil water tension is within these ranges, and therefore to the pattern rather than simply the amount of rainfall. This theory is supported by Soulides and Allison’s (1961) observations that decomposition rate was significantly increased by alternate wetting and drying of the soil. DeJong et al. (1974) found that the number of wetting and drying cycles in the soil was responsible for a large part of the annual variation in soil respiration. Wildung et al. (1973) found soil respiration rate highly correlated with soil moisture and temperature one year, but not the next. They suggested that differences in rainfall pattern between the two years might be an explanation. However, “few experiments have been de- signed to determine the influence of fluctuating soil water on microbial activity” (Nyhan and Doxtader, 1974).
Soil water may also affect microbial activity through its influence on soil temperature, which generally enters as an important variable in regressions of decomposition rate on native grasslands (Nyhan and Doxtader, 1974; DeJong et al., 1974). At least certain kinds of microbial populations are quite sensitive to temperature changes. Parker and Larson (1962) found that, in the temperature range 16°C to 2O”C, a 2°C change had a measurable effect on nitrification. At temperatures of 25°C to 30°C, this wasn’t observed. Water added during spring months may keep soil temperatures cooler and less favorable for microbial activity. In summer, added water may reduce extremely high soil temperatures and create more favorable decomposition conditions.
Increased rainfall will probably result in higher microbial activity and consequent higher rate of nutrient release. However, this may not be seen as increased nutrient concentra- tion in the soil. Power (1972) has shown that as much as 350 kg/ha of added N can be immobilized by grasslands. Much of this immobilization is done by roots, however, and increased nutrient turnover leading to increased root growth will surely be beneficial to the rangeland community.
Rainfall Pattern
The effect of timing, frequency, and amount of rainfall from single storms has generally been neglected in range research (cf. Smith, 1972) although these factors have been acknowledged as important in water use efficiency (Wight and Black, 1974), nutritive levels of grass (Rogler and Haas, 1947), community composition (Albertson and Tomanek, 1965), phenological events (Sundberg, 1974; Beatley, 1974), and insect response (Cooper and Jolly, 1969). Several authors have suggested that year-to-year differences in pattern of rainfall may explain the poor correlation between it and primary productivity (Wight and Black, 1974; Ballard and Ryerson, 1973). No experiment has been designed to specifically test this theory, and the role which rainfall pattern plays in
grassland response remains largely speculative.
Genetic Limitations
Genetic factors apparently play a significant role in the ability of a grassland to respond to improved water or nutrient supplies. Wight and Black (1974) found that mixed prairie standing crop in eastern Montana peaked in mid-July regard- less of the amount of soil water available at that time. They concluded that peak standing crop was genetically controlled and that maximum water use by this type of community was about 25 cm. Weaver (1975) suggested that genetic control of the period during which the plant makes its growth may be a factor in the failure of Idaho fescue meadows to respond to summer irrigation. Late summer dormancy is apparently common in many grass species (Laude, 1953), and suggests that cloud seeding to increase rainfall in the late summer may either be nonproductive or result in significant shifts in species composition. This will be discussed in more detail later.
Many grass species apparently do not respond to additions of water. Ballard and Ryerson (1973) found that only four out of ten species in southeastern Montana were significantly influenced by precipitation, and in only one of these, western wheatgrass (Agropyron smithii), was more than 30% of variation in production explained by rainfall. In Kansas, Shiflet and Dietz (1974) found good correlations between rainfall and production in only one of four grass species.
Bradshaw et al. (1960) and Bradshaw (1964) postulated that grasses with low yields have a selective advantage in nutrient-poor situations. The evidence indicates that evolution in the rigorous environment of the Great Plains has similarly produced at least some species whose forage production is genetically fixed at fairly low levels. There are ample data to show that the grassland community will respond to improved growth conditions, especially greater nitrogen availability. But this response is not uniform among species, and such an amelioration of the environment would almost certainly result in shifts in species composition. The direction which these shifts may take are explored below.
Effect of Added Water on Composition of the Range
Species Change
The exact nature of changes in composition due to increased precipitation will undoubtedly vary among commu- nities. Spring and early summer rains will probably favor the development of cool-season over warm-season grasses (Ballard and Ryerson, 1973). Late summer rains will perhaps do the opposite (Klages and Ryerson, 1965; Lauenroth and Sims, 1973); however, no experiment has specifically tested this. Forb production also appears to be favored by late summer and fall rains (Noller, 1968). Increased N and P availability will probably favor cool-season grasses and forbs (Williams, 1953; Cosper and Thomas, 1961; Goetz, 1969).’
Species composition of Great Plains grasslands normally changes with changing weather. The response of rangeland to drought has been thoroughly studied (Weaver and Albertson,
1939), but only one publication has documented species changes resulting from a several-year period of high rainfall. Coupland (1959) found that, during a period of unusually favorable weather on grasslands in southern Saskatchewan and Alberta, percent composition contributed by wheatgrasses and porcupine needlegrass (Stipa spartea) increased markedly,
while blue grama (Bouteloua gracilis) and needleandthread
(&pa comata) decreased. Porcupine needlegrass actively in- vaded sites which had formerly been too dry.
Experiments which modify water availability give further insights into possible shifts in species composition due to weather modification. Pitting and scalping studies in northern Montana, which increased both water and N availability, resulted in shifts of dominance from needleandthread to western wheatgrass. In some cases, these treatments have also resulted in marked increase in fringed sagewort (Artemisia
frigida), and in the appearance of grasses such as squirreltail
(Sitanion hystrix) (Ryerson et al., 1970). The long-term
effects of an environmental perturbation are well illustrated by these experiments. Fifteen years after pitting or scalping, species change is still dynamic, and fringed sagewort now appears to be decreasing.*
Availability of N and P is a major factor in determining species distribution (Lunt, 1972; t’Harte, 1949; Sonnevold et al., 1959; Bradshaw et al., 1960), and higher nutrient availability resulting from increased water will play a large role in compositional changes. Fertilizer experiments in the North- ern Plains have generally resulted in increases in western wheatgrass and decreases in blue grama (Rogler and Lorenz,
1957; Lorenz, 1970; Wight and Black, 1972).
Genotypic Change
It is likely that an altered water-nutrient regime would result in genetic change through natural selection within populations on a particular site. Genotypic shifts in response to an altered environment can occur very rapidly (Levins, 1968). The work of Bradshaw and his colleagues on adaptation of grasses to mine spoils has demonstrated this elegantly (see Gregory and Bradshaw, 1965, for a review). Harung (1974) has linked water availability to incorporation of phosphorus into nucleic acids of the desert plant, Anastatica hierochuntica L., providing. another possible mechanism for water-linked genetic change.
Little is known concerning ecotypic variation of grasses with respect to water and nutrient requirements. Genotypic differences in water requirement have been shown in orchard grass (Dactylis glomerata) (Keller, 1953; McKell et al., 1960), pubescent (Agropyron trachophorum) and intermediate wheatgrasses (Agropyron intermedium) (Baker and Hunt,
1961). Differential response to magnesium and calcium has been found among populations of bluebunch wheatgrass
(Agropyron spicatum) (Main, 1974). However, the extent of
such differentiation in nature remains to be determined. Blue grama, for example, appears to vary in its water requirements. In Colorado, it is noted for its ability to respond to favorable growing conditions anytime during the year (Turner and Klipple, 1952), but Ballard and Ryerson (1973) could find no correlation between rainfall and blue grama production in Mont ana.
Genetic change (including shifts in species) is a common response of nature to a changing environment. It is not a cause for concern unless it results in poorer forage producers or a less stable community. The evidence presented above indicates that community change resulting from increased rainfall in the spring and early summer would be toward higher yielding, more desirable forage species.
’ Also from personal communications with J. Ross Wight. ‘Personal communication with Harold A. R. Houlton.
Stability is directly related to the degree of success which weather modification operations have. If rain is increased during normally wet years and not affected during drought, the result will be an increase in the amplitude of climatic variation. What this means in an already liighly variable environment is not known. If periods of favorable weather result in dominance by more mesic species or genotypes, the ability of the community to respond successfully to drought will involve a much higher compositional flux than at present. Effects of a drought could be more severe; recovery could take longer.
Klages and Ryerson (1965) demonstrated quite clearly some problems which may be encountered. They added water and nitrogen to native range in 1958, 1959, and 1960 with significant increases in yield. 1960 and 1961 were drought years, and by 1962, grasses in previously fertilized and watered plots had decreased much more than in control plots. Needleandthread was virtually eliminated from experimental plots, while forbs increased. Apparently 3 years of exception- ally favorable growing conditions had reduced this commu- nity’s ability to withstand unfavorable conditions.
Effect of Added Water on Secondary Productivity
Cooper and Jolly (1969) suggest that there would be no major insect pest outbreaks as a result of weather modifi- cation. Data from irrigated and fertilized range plots in Colorado show insect biomass increases which are generally consistent with increases in forage production (Lavigne and Kumov, 1974).
Among arthropods, response to an ecosystem perturbation is dependent on trophic level and successional status of the community (Hurd and Wolf, 1974). There is significant variation in the response of closely associated grasshopper populations to moisture, temperature, and starvation (Hastings and Pepper, 1964) and there are year-to-year fluctuations in grasshopper populations “which are not the direct result of vegetational changes alone, nor probably of any single environ- mental factor” (Anderson, 1964). However, Edwards (1960) did find a weak relationship between grasshopper numbers in Saskatchewan and April-August precipitation in the preceding 2 years. Riegert (1968) found grasshopper infestations in Saskatchewan increase in warm, dry weather, whereas cool, moist conditions caused population declines.
Unexplained fluctuations in the populations of small mammals make correlation of then numbers with weather variables very difficult. Okulora and Myskin (1974) found that numbers of the northern red-backed vole
(Clethrionomys
nutilus)
correlated principally with temperature; however, there was an inverse correlation with spring precipitation. Lauenroth and Sims (1973) found shifts in small mammal species composition coupled with significantly smaller biomass in plots which had been irrigated for 2 years.The response of large herbivores to change in primary production could probably be generalized from cattle data, if cattle data allow a generalization. Ballard and Ryerson (1973) found that in southeast Montana, calf weights were significant- ly correlated (positively) to growing season and previous year precipitation, but gain of wet cows was not. In a higher rainfall zone in central Montana, there was no relationship between precipitation and cattle weight gains. In a North Dakota study, weaning weights of calves were lowest for years which received the greatest amounts of rainfall from April through July (Johnson et al., 1974). In Colorado, it was found that 1 inch
or more rainfall in July or August enhanced steer weight gains significantly (Bose et al., 1975; Sims et al., 1976; cited in Cook, 1975). Whether this would be true in more northerly grasslands, with a smaller component of warm-season grasses, is not clear.
The effect of increased forage on cattle weights will depend largely on its nutritive value. The effects of increased water on the protein content of grasses is quite variable. In many parts of Montana, optimum nutritive content is reached at precipita- tion levels slightly under average.3 Willard and Schuster (1973) found that crude protein of six grasses from the Southern Great Plains was highest when rainfall was adequate for vegetative growth. In the Northern Great Plains, Wight and Black (1974) reported that nitrogen concentration in above- ground biomass declined throughout the growth period in- dependently of rainfall, while phosphorus concentration fluc- tuated as a function of precipitation. Thomas and Osenbrug (1964) found that the nitrogen content of fertilized grasses was diluted by added water.
Relationship between nutrient content of grasses and precipitation is probably dependent on a number of factors, including grass species and timing of the rainfall. Crude protein percent increases in blue grama, but not in western wheatgrass, following a summer thunderstorm (Rauzi et al., 1969). In grasslands with a significant component of blue grama, rains in the late summer would perhaps be the most productive from the standpoint of maintaining an adequate nutrient supply. Where western wheatgrass predominates, the evidence indi- cates this would not be the case; however, it is a question that needs further research. The possibility exists that, given rain in the proper season, warm-season grasses could assume a more important role in northern grasslands.
Other Potential Effects
Plant Diseases
Broadbent (1967) reviewed the research on the influence of weather on plant virus diseases, and concluded that it is “considerable and very complex (and) has been adequately studied in relation to very few diseases. . . .”
Aphids and leafhoppers are the principal vectors of plant diseases. Aphids multiply faster in warm, dry summers than in cool, moist ones; however, in dry areas, wet periods which favor the growth of plants may also favor the proliferation of the insects that feed on them (Bradbent, 1967). Curly top virus of beet is increased by spring rains because of increased germination of annual weeds which harbor the virus (Severin, 1939) and in California, rain encourages the growth and subsequent infestation of grasses by aphids which carry cereal yellow dwarf virus (Oswald and Houston, 1953).
Temperature effects on aphids and leafhoppers are mixed. In general, warm temperatures increase aphid movement and thus infection rates (Bald et al., 1950); however, very warm temperatures may result in a cessation of aphid activity altogether (van der Plank, 1944). Hot weather may either increase or decrease the infectivity of leafhoppers, depending on species (Kunkel, 1937; Webb, 1956).
The possible effects of increased moisture on soil-borne pathogens are mixed. Fungi are killed by prolonged conditions that favor activity of anaerobic bacteria. Those without adequate dormant structures may be controlled to a certain extent by low water tensions in surface soils (Bruehl, 1969).
3 Personal communication with Gene F. Payne.
added water may produced by added rainfall will probably result in higher livestock gains. At higher rainfall levels, however, the effect on livestock may be minimal and perhaps even negative. The response will largely be governed by the nutrient content of the forage. Further research is needed on this question.
Once again, the implication is that timing of be the important factor in determing effects.
Parasites
The liver fluke is an important parasite of cattle and sheep in many areas. Two conditions are necessary for survival and spread of flukes: (1) an adequate population of the snail,
Lynnaea truncatula, which is an essential host during a portion
of the fluke’s life cycle; and (2) saturated soil for movement of the fluke from host to host (Ollerenshaw, 1967). A concern has been voiced that enhanced rainfall may increase that incidence of liver fluke disease in the Northern Great Plains (now virtually nonexistent). However, Ollerenshaw (1967) has determined that, in England, 3 consecutive months in which rainfall exceeds evaporation are necessary for the fluke to complete its life cycle. In addition, areas of standing water are necessary for suitable snail habitat. It seems unlikely that weather modificaton in the semiarid Great Plains will be successful enough to fulfill either of these conditions.
Runoff
The amount of runoff produced by increased rsinfall will depend on soil type, vegetative cover, and the way in which storm intensity is affected by precipitation modification. On mixed prairie, grassland condition exerts a greater control over infiltration than soil type. On semiarid sites, because of sparse vegetation, the opposite is true (Wolf, 1970).
Wolf (1970) reported that the average 1 -hour duration storm was capable of being infiltrated on practically all range sites which he studied. The average lo-minute duration storm would be infiltrated where range conditions were good to excellent, but not when they were fair to poor. Branson and Owen (1970) found that 74% of the variation in runoff from watersheds underlain by Mancos shale was explained by the proportion of bare soil in the watershed.
The above data indicate that in the short run, runoff in semiarid areas may be increased by precipitation enhancement. However, if increased rainfall results in increased vegetative cover, runoff will decrease proportionately and the long-term effect will be beneficial.
Summary and Conclusions.
1. Over the short term, aboveground primary production may be increased slightly by spring and early summer rains, but without fertilization maximum yields per inch of added rainfall will probably not exceed 50- 110 lb/acre. Increases from added precipitation will probably be considerably higher in years of low than in years of high total growing season precipitation.
2. The long-term effect of added water on primary production will depend on the degree to which decomposition rates are increased, and therefore, the rate at which nutrients are released. Higher rainfall will probably increase nutrient cycling rates; however, research is needed to quantify this effect.
3. Compositional changes will almost certainly occur in the community, the exact nature of which will be highly depen- dent on the pattern of rainfall increases. Enhanced precipita- tion in the spring (April and May) will probably result in increased proportions of more mesic, high producing species. Summer and fall rains will probably affect mainly warm-season grasses and forbs; however, research is needed on this point.
4. The effect of increased forage production on livestock weights is not clear. In periods of deficient water, the forage
5. Changes in consumer populations will probably not be dramatic, although small mammal numbers may decline. The effect such a decline would have on community function is not known.
6. Cool, moist weather during the growing season could in declines in grasshopper populations.
Enhancement of precipitation could result result
7. in communi-
ties which are less able to- withstand adverse conditions. If drought periods cannot be affected by weather modification, more extreme fluctuations in the vegetation may result.
8. In the long run, runoff will be reduced in an amount proportionate to increased vegetative cover, providing there isn’t a significant increase in storm intensity.
9. There will be both positive and negative effects on diseases of plants. The net effect cannot be predicted.
Literature Cited
Albertson, F. W., and G. W. Tomanek. 1965. Vegetation changes during a 30-year period in grassland communities near Hays, Kansas. Ecology 46:714-720.
Anderson, Norman L. 1964. Some relationships between grasshoppers and vegetation. Annu. Entomol. Sot. Amer. 5 7:736-742.
Baker, John N., and 0. J. Hunt. 1961. Effects of clipping treatments and clonal differences on water requirements of grasses. J. Range Manage. 14:216-219.
Bald, J. G., D. 0. Norris, and G. A. Helson. 1950. Transmission of
potato virus diseases. VI. The distribution of the aphid vectors on sampled leaves and shoots. Aust. J. Agr. Res. 1:18-32.
Ballard, William, and D. E. Ryerson. 1973. Impacts of induced rainfall on the Great Plains of Montana-Section 1:Range livestock produc- tion. Mont. Agr. Exp. Sta. Res. Rep. 42.
Beatly, Janice C. 1974. Phenological events and their environmental triggers in Mojave Desert ecosystems. Ecology 55 :85 6-8 63. Bleak, Alvin T., Wesley Keller, and A. C Hull, Jr. 1974. Range plant
yield and species relationships in natural and partially controlled environments. J. Range Manage. 27 : 396-399.
Bose, D. R., Herb Mann, and C Wayne Cook. 1975. Grazing research in southeast Colorado 1974. Colo. State Univ. Exp. Sta. Progr. Rep. No. 2.
Bradshaw, A. D. 1964. Experimental investigations into the mineral nutrition of several grass species. IV. Nitrogen level. J. Ecol. 52:665-676.
Bradshaw, A. D., J. J. Chadwick, D. Jowett, and R. W. Snaydon. 1960.
Experimental investigations into the mineral nutrition of several grass species. III. Phosphate level. J. Ecol. 48: 631-637.
Branson, F. A. 1956. Range forage production changes on a water- spreader in southeastern Montana. J. Range Manage. 9: 187-191. Branson, F. A., and J. R. Owen. 1970. Plant cover, runoff, and
sediment yield relationships on Mancos shale in western Colorado. Water Resources Res. 6:7 83-790.
Broadbent, L. 1967. The influence of climate and weather on the incidence of plant virus diseases. p. 99-104. In: James A. Taylor (Ed.), Weather and Agriculture. Pergamon Press, London.
Bruehl, G. W. 1969. Factors affecting the persistence of fungi in soil. p. 11-13. In: Roy A. Young and William C. Snyder (Ed.), Nature of the Influence of Crop Residues on Fungus-Induced Root Diseases. Wash. Agr. Exp. Sta. Bull. 7 16.
Caprio, Joseph M., and Jack S. Williams. 1973. Impacts of induced rainfall on the Great Plains of Montana-Section 4: Agricultural production. Mont. Agr. Exp. Sta. Res. Rep. 42.
Cook, C W. 1975. Possible benefits of weather modification on rangeland production. p. 13-14. In: Lewis 0. Grant and John D. Reid (Ed.), Workshop for an Assessment of the Present and Potential Role of Weather Modification in Agricultural Production. Colo. State Univ., Dep. Atmospheric Sci., Fort Collins, Atmospheric Sci. Pap. No. 236.