Chromosome Pairing Does Not Contribute to Nuclear Architecture in Vegetative Yeast Cells
Full text
Figure

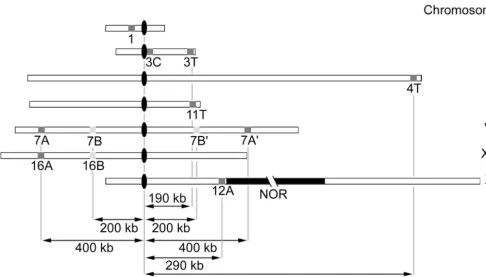

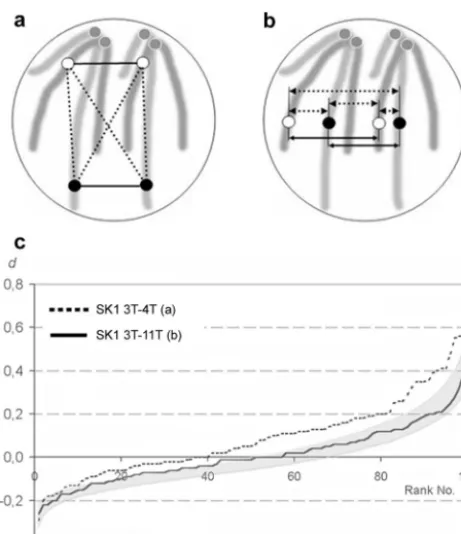
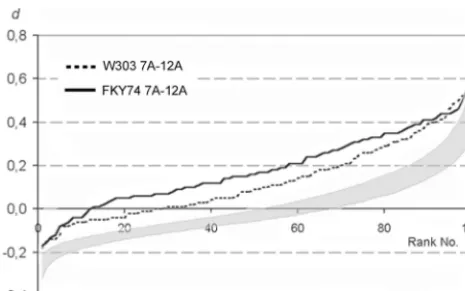
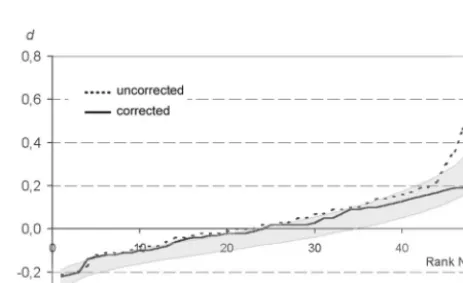
Related documents
If to look at the experience of modern gender equality, although Fatma Aliya khanum is not exactly a feminist writer, the fact that she brought the female
to be in favor of the President’s power to disregard the restrictions because the President is exercising a constitutional power, not a power granted under a statute passed
Such a process is different from the example we had in Figure 1, called a worker-paced line flow, as it does not allow for a build up of inventory between the process steps.. The
Utilizing ERP systems software, such as SAP R/3, allows information management for complex organizational forms such as, supply chain, virtual and extended enterprises that require
With di ff erent updating methods for the weights, two detectors are proposed: Hard Decision for MSTR (HD-MSTR) detector and Soft Decision for MSTR (SD-MSTR) detector, where
decimal point (A) to be between the sign and the most significant decimal digit. In single-precision, floating-point notation, the two digit positions'..
Note: When Shake Qmaster Admin Tools are installed, the Shake Qadministrator icon appears in the Applications folder and the Batch Monitor icon appears in the
Empat perkara utama yang menjadi teras kepada pembentukan program G7 selain daripada meningkatkan prestasi akademik pelajar sekolah-sekolah terlibat adalah perkongsian dalam
