http://www.sciencepublishinggroup.com/j/ajls doi: 10.11648/j.ajls.20180601.12
ISSN: 2328-5702 (Print); ISSN: 2328-5737 (Online)
A Quantitative Study of Nocodazole’s Effect on HeLa Cells’
Growth Rate and F-actin Structure
Jiayi Lu, Jorge De La Torre, Carson McCann, Maureen Madar, Qiongqiong Zhou
Department of Biological Sciences, Denison University, Granville, USA
Email address:
To cite this article:
Jiayi Lu, Jorge De La Torre, Carson McCann, Maureen Madar, Qiongqiong Zhou. A Quantitative Study of Nocodazole’s Effect on HeLa Cells’ Growth Rate and F-actin Structure. American Journal of Life Sciences. Vol. 6, No. 1, 2018, pp. 7-12. doi: 10.11648/j.ajls.20180601.12
Received: January 19, 2018; Accepted: March 2, 2018; Published: March 9, 2018
Abstract:
HeLa cells were derived from human cervical cancer, which has been widely used as research model to study mammalian cell functions. In this work, nocodazole, a microtubule destabilizer, was used to treat HeLa cells with different concentrations. The results showed that nocodazole was able to inhibit HeLa cell’s growth rate significantly at the concentration of 100 nM. It suggested that nocodazole may inhibit cell growth through an alternative impacting effect other than destabilizing microtubules, since the effect of nocodazole destabilizing microtubule is usually not seen at micromolar range. However, at nanomolar concentration, nocodazole was not able to induce any changes in F-actin structure. Whereas, at concentration of 1 µM, nocodazole induced significant alterations of F-actin structure in HeLa cells. It indicated the strong relationship between microtubule and actin dynamics. Our work suggested that a molecule may exhibit different impacting mechanisms at different concentrations. Since nocodazole has been used as a chemotherapy reagent in cancer treatment, it will be beneficial to re-evaluate the effective concentration in terms of cancer treatment.Keywords:
Nocodazole, F-actin, Microtubules, Growth Rate, HeLa Cells1. Introduction
HeLa cells – the first continuous cancer cell line - were isolated from human cervical cancer, which has been widely used as a research model to study mammalian cell functions. Almost every mammalian cellular process has been tested in HeLa cells [1, 2, 3].
Nocodazole, a benzimidazole derivative, was initially developed as a potential anticancer drug [4]. The drug became a popular treatment for cancer due to its ability to prevent mitosis. Besides it’s medical application, nocadazole has also been widely used as a biomedical research tool to many cellular functions. Nocodazole inhibits cell growth by binding to tubulin and thus suppressing microtubule dynamics or inducing microtubule depolymerization, thus leading to apoptosis in dividing cells [5]. Other studies argue that Nocodazole induces mitotic arrest due to a decrease in microtubule dynamic turnover, meaning the drug significantly hindered the elongation of microtubules [6]. This discrepancy may be resulted from different doses related function of nocodazole on mammalian cells, which has not been heavily investigated.
In this experiment, we tested the survival rate of the cultured HeLa cells by introducing different concentrations of Nocodazole. At concentration of 10 M, Nocodazole blocks microtubule polymerization and it does so by taking away α/β tubulin dimers in dictyostelium cells, thus inhibiting the growth of the dicty cells [7, 8]. However, work done by Blajeski et al showed that at 1 M, nocodazole functioned to destabilize microtubules and arrest cell cycle [9]. And work done by Vasquez et al showed that at nanomolar concentration, nocodazole was able to inhibit microtubule functions. [10]
The microtubule-actin interactions are constant and critical in cells, especially in events like cell division and cell migration [11, 12, 13]. During cell division, the structural integrity of the spindle apparatus is essential for the formation of actomyosin ring at the end of anaphase and the beginning of cytokinesis. The cell would not enter cytokinesis if the spindle apparatus is disrupted, but would not stop cytokinesis once the cleavage furrow formation starts, even if spindle apparatus is destroyed [14].
microtubules from the aster center until reaching the midpoint of two daughter cells. This process is especially important in asymmetrical cell division [15]
During mitosis, the plus end of the spindle apparatus binds to actin, which then helps to orientate the spindle and binds to specific regions in the cell cortex, to make sure that the future division plane is properly placed [16].
During cell migration, when cell is moving towards a certain direction, microtubules grow along the actin fibers at the leading edge of cell migration, push the cell forward. Microtubules are then bound to F-actin meshwork undergoing retrograde flow in lamellum. Microtubules are compressed and broken down in this process, which gives space to new microtubules to grow along the actin fibers and keeps the cell moving forward [17]
In another study that focuses on the quantitative regulation of microtubule and actin, both microtubule and actin are found in the Rho A signaling pathway. When microtubules in a cell are assembled, protein GEF-H1 is released, which can activate Rho A, the protein responsible for both the formation of Actin and the stabilization of microtubules. This mechanism implies that the disassembly of microtubules would trigger the Rho A pathway and increase the amount of actin in cells [18].
These wide varieties of actin-microtubule interaction raise the question of how the change in one can affect the other. In this experiment, we intended to find out what would happen to actin in cells if we use drugs to depolymerize the microtubules in a cell. Therefore, we hypothesized that disturbance of microtubule networks by nocodazole will lead to changes in F-actin structures and the growth rate of HeLa cells.
2. Materials and Methods
HeLa cells were obtained from ATCC (American Tissue Culture Center) and cultured in DMEM media supplemented with 10% FBS and antibiotics. Cell culture related reagents, DMEM media, PBS, Trypsin, FBS, were purchased from Life Technologies. Nocodazole was purchased from Sigma.
The growth rate of hela cells was first determined by setting four 2 cm plates and adding 2ml of media in each one of them, with nocodazole or DMSO. All four plates were incubated at 37°C. On the 1st, 3rd, 4th, and 6th day after plating, one of the 2 cm plates were taken out of the incubator and the cells were counted using a hemocytometer. The cell number was calculated using the following equation: After collecting data from all four 2cm plates, a time vs.
cell number equation (Y=ae^bX) was generated using JMP software, the doubling time (Xd) can be calculated as follow:
Doubling time (Xd)= ln2/b
F-actin in nocodazole treated cells and control cells were stained using a F-actin visualization kit from Cytoskeleton Inc. One day before the staining, HeLa cells were cultured in 2-well chambered slides. Cells were treated with nocodazole at 1M, 100 nM or DMSO for 24 hours.
The next day, cells were washed with warm PBS, fixed by 4% paraformaldehyde, and permeabilized by triton X 100. Cells were then incubated with Rhodamine-phalloidin (100 nM) for 20 min. Finally, slides with cells were mounted with mounting media with DAPI.
3. Results
3.1. The Growth Rate of HeLa Cells
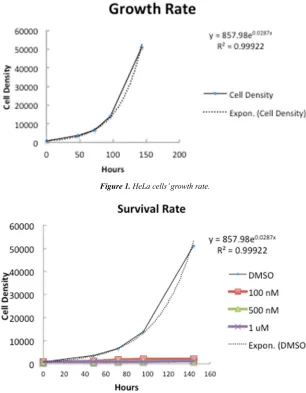
In order to study the effect of nocodazole on the growth rate of HeLa cells, HeLa cells growth rate without nocazole was initially determined (Fig 1). Cells growth dynamics was monitored in a period of 150 hours. Cell densities in dishes were determined every 24 hours. Cell density against time was plotted as figure 1. Data was then fit by an exponential curve: y=858e0.029x. The calculated growth rate was 0.029, which means cells divide in every 23.9 hours.
3.2. Nocodazole Inhibit the Growth of HeLa Cells
Next, we want to test how nocodazole can inhibit the growth of HeLa cells. Cells were again cultured in 2-cm dishes, nocodazole at different concentrations or DMSO were added to cells. Cell density again was determined by using hemocytometer daily. As low as 100 nM, nocodazole was able to inhibit most of the cell growth. At the same time, the 500 nM and 1 µM treatments exhibited the most growth inhibition (Figure 2). The control, the DMSO treatment, showed normal growth of HeLa cells according to the previously analyzed growth rates in figure 1.
In addition to the investigation on growth inhibition of nocodazole at different concentrations, cell viability was determined by MTT assay after cells incubated with nocodazole. HeLa cells were cultured in 2-cm dish and incubated with nocodazole at different concentrations for 48 hours. Cell viability data were shown in table 1 and figure 3. At concentration as low as 100 nM, nocodazole was able to impact cell viability.
Table 1. Cell viability assay by MTT absorptions.
Noc Concentrations Absorption 540 nm Relative Cell Viability
0 nM 0.869 100%
100 nM 0.549 63%
500 nM 0.274 32%
Figure 1. HeLa cells’ growth rate.
Figure 2. Nocodazole’s inhibitory effect on HeLa cell’s growth rate.
Figure 3. Cell viability assay by MTT absorptions.
3.3. The Effect of Nocodazole on F-Actin Structure in HeLa Cells
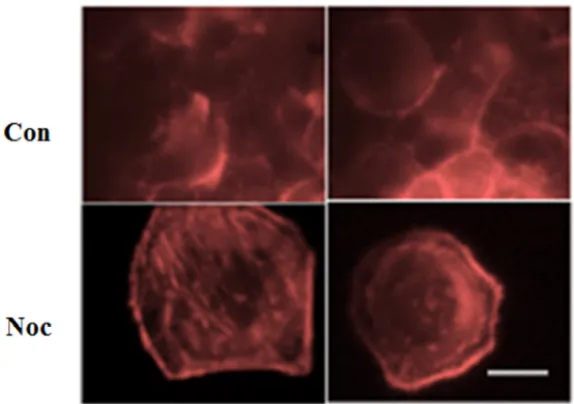
Next, we want to test how different concentrations of nocodazole may effect the structure and dynamics of F-actin in HeLa cells. HeLa cells were cultured in chambered slides. Nocodazole or DMSO were added to cells. After 24 hours of
incubation, cells were fixed and stained to visualize the F-actin structure (Figure 4). It was consistently observed that cells treated with 1 mM nocodazole showed stronger F-actin signal compare to the control cells. Therefore, it is possible that dissolved tubulin may have a positive effect on the polymerization of F-actin filaments.
0% 20% 40% 60% 80% 100% 120%
0nM 100nM 500nM 1000nM
R
e
la
v
e
ce
ll
v
ia
b
il
it
y
Noc
Conc.
Figure 4. F-actin staining by Rhodamine-Phalloidin. Control cells were incubated with DMSO. Treatment cells were incubated with 1 µM nocodazole for 24 hours. Scale bar: 10 µm.
Figure 5. Cell radius measurement. HeLa cells were treated with nocodazole at concentration of 1 µM for 24 hours. Cells were then fixed and stained with Rhodamine-phalloidin. Cell radius was quantified by measuring the cell area indicated by rhodamine-phalloidin.
4. Discussion
After cells treated with high concentration of nocodazole, we could observe clear structure and quantity difference in F-actin in nocodazole+/- conditions. The amount of F-actin appears to be higher when cells are exposed to nocodazole, it is possible that the microtubule depolymerization triggered Rho A pathway and synthesized more actin. Since Rho A can not stabilize microtubules when nocodazole is present, more and more actin were made during the process of Rho A trying to stabilize microtubules [18].
Structurally, we could see a clear circular distribution of F-actin in cells that were exposed to nocodazole, while F-actin in control group shows no clear pattern in their distribution. Actin serves an essential role in mitosis by help spindle apparatus to orientate and bind to specific regions in the cell, to make sure that the placement of future division plane is correct. It is possible that in the nocodazole treated cells the actin spreads out in the cell and prepares to bind to
microtubules during mitosis, but since microtubules were depolymerized by nocodazole, actin has nothing to binds to and stays in the circular distribution. However, in the control group, actin served their function by properly orientated microtubules and actin has moved out of this circle [16].
These clear differences in F-actin give us reason to conclude that our hypothesis, that if microtubules in cells are disassembled, we would be able to see a significant change in either the structure or the quantity of actin in the cell.
However, we are still uncertain of what caused these changes, and a lot of mechanisms involved in these changes remain unknown. Future research can be focused on looking for the exact mechanisms that caused these changes.
higher than 1 µM, it is reasonable for us to contemplate that how does nocodazole exert the inhibitory effect at the concentration of 100 nM. An alternative mechanism of nocodazole inhibit cell division may exist.
For the survival test of our HeLa cells when treated with Nocodazole, cell density counting revealed interesting data that supported our hypothesis. As expected, the 1 µM of nocodazole treatment exhibited the least cell density at the end of the treatment, followed by the 500 nM (Figure 3). These two results show that both of these Nocodazole concentrations were effective in reducing the HeLa cell populations. The control treatment (DMSO) exhibited a cell density that was expected according to the previously analyzed growth rate of our HeLa cells in regular conditions. The 100 nM Nocodazole treatment group showed the highest post-treatment value of cell density (Figure 3) and thus the most growth over the course of the experiment, supporting our hypothesis. This finding can be related to Darwinian Evolution as these cells were treated with enough Nocodazole to develop adaptions that allowed the cells to thrive when exposed to Nocodazole and as the treatment concentration was not effective nor lethal, it allowed the cells that developed adaptions, the “most fit” cells in Darwinian language [19], to proliferate and thus pass on their resistance against Nocodazole to the newly generated cells.
In the future experiment, nocodazole at concentration of 10 nM and 50 nM will be tested as well to see how effective these concentration may inhibit cell growth.
However, in a shorter incubation period, 100 nM of nocodazole showed 30% inhibitory effect on cell viability. All together, these results suggest cells may not able to metabolize nocodazole effectively. At low concentration, the drug effect may last for 4-5 days. It also suggests that in terms of treatment, low concentration in combination with long treatment period maybe an option.
The doubling time of HeLa cells was measured to be 23.9 hours. The previous study done by Meck et al [20] found that the doubling time of HeLa cells in vitro was about 28.8 hours. The result obtained in this experiment was close to but not exactly matches with Meck’s result. There are several possible reasons for the discrepancy. The HeLa cells have been cultured extensively, therefore HeLa cells from different lab or batch may have different features since they are at high risk of mutation. Secondly, the culturing condition maybe slightly different between lab to lab. Overall, knowing the growth rate is helpful, so that researchers could potentially use this number to assess their cell line’s growth condition.
It will also be interesting to assess nocodazole’s drug effect on other cancer cells. Nocodazole, although it has been used as an anti-cancer drug for a long time, is still used in some cases clinically. Determine the effective concentration and treatment period will be very helpful in determine the drug amount and treatment period clinically. Our results suggest that a low dose but longer treatment combination maybe considered to use nocodazole to have negative impact on cancer cell proliferation.
In summary, this series of experiments with HeLa cells serves as an example of how effective these cells are for culturing, treatment exposure, and characterization in vitro. There were some limitations to our experiments such as the short periods of observation of growth short, or the fact that we only experimented with three different concentrations of Nocodazole. For future studies with HeLa cells we recommend larger observation periods and replications of such in order to accurately monitor the growth rate of these cells. For studies that also aim at observing Nocodazole’s effect on cell structures we recommend testing with more concentrations as well as performing in vivo experiments in which Nocodazole was introduced to cancerous cells in a mice model, if plausible this experiment could give more insight into Nocodazole’s ability to disrupt the F-actin filaments in vivo.
5. Conclusion
In conclusion, our results indicate that nocodazole was able to inhibit cell growth in a nanomolar range concentration. At micromolar concentration, the disturbance of microtubule clearly has an impact on the F-actin dynamics and structure.
References
[1] Masters JR. Nat Rev Cancer. 2002 Apr;2(4):315-9. Review. [2] Beskow LM. Annu Rev Genomics Hum Genet. 2016 Aug
31;17:395-417. doi: 10.1146/annurev-genom-083115-022536. Epub 2016 Mar 3. Review.
[3] AccessScience Editors. (2014). Importance of HeLa cells. In AccessScience. McGraw-Hill Education.
[4] Blajeski, A. L., Phan, V. A., Kottke, T. J., & Kaufmann, S. H. (2002). G1 and G2cell-cycle arrest following microtubule depolymerization in human breast cancer cells. The Journal of Clinical Investigation, 110(1), 91–99.
[5] Jordan, M. A., Thrower, D., & Wilson, L. (1992). Effects of vinblastine, podophyllotoxin and nocodazole on mitotic spindles. Implications for the role of microtubule dynamics in mitosis. J cell Sci, 102(3), 401-416.
[6] Vasquez, R. J., Howell, B., Yvon, A. M., Wadsworth, P., & Cassimeris, L. (1997). Nanomolar concentrations of nocodazole alter microtubule dynamic instability in vivo and in vitro. Molecular biology of the cell, 8(6), 973-985.
[7] Ngo T, Miao X, Robinson DN, Zhou QQ. Acta Pharmacol Sin. 2016 Nov;37(11):1449-1457. doi: 10.1038/aps.2016.57. Epub 2016 Aug 29.
[8] Zhou Q, Kee YS, Poirier CC, Jelinek C, Osborne J, Divi S, Surcel A, Will ME, Eggert US, Müller-Taubenberger A, Iglesias PA, Cotter RJ, Robinson DN. Curr Biol. 2010 Nov 9;20(21):1881-9. doi: 10.1016/j.cub.2010.09.048. Epub 2010 Oct 14.
[10] Vasquez RJ, Howell B, Yvon AM, Wadsworth P, Cassimeris L., Mol Biol Cell. 1997 Jun;8(6):973-85.
[11] Poulter NS, Thomas SG. Int J Biochem Cell Biol. 2015 Sep;66:69-74. doi: 10.1016/j.biocel.2015.07.008. Epub 2015 Jul 23. Review.
[12] Akhshi TK, Wernike D, Piekny A. Cytoskeleton (Hoboken). 2014 Jan;71(1):1-23. doi: 10.1002/cm.21150. Epub 2013 Oct 29. Review.
[13] Cabrales Fontela Y, Kadavath H, Biernat J, Riedel D, Mandelkow E, Zweckstetter M. Nat Commun. 2017 Dec 7;8(1):1981. doi: 10.1038/s41467-017-02230-8.
[14] HIRAMOTO Y. J Cell Biol. 1965 Apr;25:SUPPL:161-7. [15] Sider, J. R., Mandato, C. A., Weber, K. L., Zandy, A. J., Beach,
D., Finst, R. J.,. Bement, W. M. (1999). Journal of Cell Science, 112(12), 1947.
[16] Lutz, D. A., Hamaguchi, Y. & Inoue, S. (1988). Cell Motil. Cytoskeleton 11, 83–96.
[17] Forscher, P., Bement, W. M., Waterman-Storer, C. M., Mandato, C. A., Schaefer, A. W., & Rodriguez, O. C. (2003). Nature Cell Biology, 5(7), 599-609.
[18] Palazzo, A. F., Cook, T. A., Alberts, A. S. & Gundersen, G. G. (2001). Nature Cell Biol. 3, 723–729.
[19] Peitzsch, C., Tyutyunnykova, A., Pantel, K., & Dubrovska, A. (2017, February). In Seminars in Cancer Biology. Academic Press.