Copyright1999 by the Genetics Society of America
A Quantitative Model of the Relationship Between Phenotypic Variance
and Heterozygosity at Marker Loci Under Partial Selfing
Patrice David
CEFE-CNRS, 34293 Montpellier Cedex 5, France
Manuscript received March 16, 1999 Accepted for publication July 14, 1999
ABSTRACT
Negative relationships between allozyme heterozygosity and morphological variance have often been observed and interpreted as evidence for increased developmental stability in heterozygotes. However, inbreeding can also generate such relationships by decreasing heterozygosity at neutral loci and redistribut-ing genetic variance at the same time. I here provide a quantitative genetic model of this process by analogy with heterozygosity-fitness relationships. Inbreeding generates negative heterozygosity-variance relationships irrespective of the genetic architecture of the trait. This holds for fitness traits as well as neutral traits, the effect being stronger for fitness traits under directional dominance or overdominance. The order of magnitude of heterozygosity-variance regressions is compatible with empirical data even with very low inbreeding. Although developmental stability effects cannot be excluded, inbreeding is a parsimonious explanation that should be seriously considered to explain correlations between heterozygos-ity and both mean and variance of phenotypes in natural populations.
N
EGATIVE relationships between allozyme hetero- Although mentioned in several instances (Eaneszygosity and morphological variance have been 1978;Chakraborty1987;Yezerinac et al. 1992), the
found in various species (Eanes 1978; Mitton 1978; role of genes other than the marker loci has not been
Zouroset al. 1980;MittonandKoehn1985;Yezeri- considered theoretically. This contrasts with
heterozy-nacet al. 1992;Davidet al. 1997) although not consis- gosity-fitness correlations, as various theoretical studies
tently (Handford 1980; McAndrewet al. 1982; Zink analyzed the apparent heterozygote advantage at
neu-et al. 1985; Booth et al. 1990; Livshits and Smouse tral markers due to correlations with fitness genes, i.e.,
1993;Bamshadet al. 1994; YampolskyandScheiner associative overdominance (Ohta1971;Ohtaand
Cock-1994;Gardner1995). With reference to concepts devel- erham 1974; Strobeck 1979; Charlesworth 1991;
oped byLerner(1954), these data have generally been Zouros 1993). Indirect effects on
heterozygosity-vari-interpreted as evidence for a higher developmental ho- ance correlations were not investigated as the relevant
meostasis in heterozygotes (Eanes1978;Mitton1978; models (Chakraborty and Ryman 1983;
Chakra-LivshitsandKobyliansky1985;MittonandKoehn borty1987) only considered infinite panmictic
popula-1985). Under this hypothesis, changes in phenotypic tions that lack correlations among loci. In this case
rela-variance among genotypes reflect changes in rates of tionships between phenotype and marker loci reduce
developmental error. Individual phenotypes are there- to their direct phenotypic contribution (Smouse1986).
fore considered as random variables with a fixed expec- Two factors promote genotypic correlations: (i) linkage
tation (the population mean) and different variances, desequilibrium due to small population size (Hilland
depending on heterozygosity. Alternatively, individuals Robertson 1968) and (ii) partial inbreeding, which
may have different expectations, their developmental generates identity desequilibria (i.e., heterozygosity
cor-variance being constant or negligible. This was first pro- relations; Bennett and Binet 1956; Weirand
Cock-posed byChakrabortyandRyman(1983), who mod- erham 1973). I here focus on partial inbreeding and
eled a phenotype determined by additive alleles. In this its consequences on heterozygosity-variance and
hetero-case, heterozygotes are phenotypically intermediate be- zygosity-mean correlations. I examine the effects of
dif-tween homozygotes. As homozygotes include two geno- ferent genetic determinisms of the phenotype
(additiv-types with different means, their phenotypic variance ity, dominance, and overdominance) and of the
exceeds that of heterozygotes. With several diallelic loci, number of marker loci.
this is amplified by a combinatory effect as many differ-ent genotypes are included in homozygotes for n loci
(ChakrabortyandRyman1983;Chakraborty1987). GENETIC VARIANCE AND MARKER GENOTYPES IN A PARTIALLY INBRED POPULATION
Distribution of phenotypic values among inbreeding
Address for correspondence: CEFE-CNRS, 1919 route de Mende, 34293
Montpellier Cedex 5, France. E-mail: [email protected] classes:I consider an infinite population at inbreeding
equilibrium. A proportion s of offspring are produced where the additive and dominance variance under ran-dom mating are VA5Ri2piqi[ai2di(pi2qi)]2and VD5
by selfing, whereas (12s) come from random matings.
Each individual belongs to a given inbreeding class k, Ri4p2iqi2d2i(FalconerandMackay1996), respectively.
Distribution of phenotypic values among genotypes: being the number of generations of selfing in his
pedi-gree, starting from the last outcrossing event (0# k# The genotype G also contains various inbreeding classes,
but the frequency Gk of class k within G differs from
∞). Class k has frequency (12s)skassuming no selection.
Although inbreeding depression usually occurs in par- the population frequency. For example, homozygous
genotypes contain high frequencies of inbred
individu-tially inbred populations (Charlesworthand
Charles-worth 1987), neutral frequencies computed using a als (large k). The mean and variance within G are there-fore
lower value s9(detailed inappendix a) provide a good
approximation of this case. s may be replaced by s9to φ
G5
o
kGkφk (6)
account for selection in the expressions below.
We assume a neutral phenotypeφdetermined by L
and quantitative trait loci (QTL). Each QTL i has two alleles
Aiand Biwith respective frequencies piand qi. Its contri- s2G(φ)5 s2(within inbreeding classes)
butions to the phenotype are ai, di, and2aifor genotypes 1 s2(among classes)
AiAi, AiBi, and BiBi, respectively, and are additive over
loci. Genotypes are assessed at a set of marker loci (ML) 5
o
k
Gks2k1
o
kGk(φk2 φG)2. (7)
different from the QTL. Linkage equilibrium is assumed
for all pairs of loci (QTL and ML) as a consequence of Using these general formulas, I now examine the
differences in phenotypic mean and variance among infinite population size. Throughout the article, I focus
on the simplest case of unlinked loci, as linkage merely genotypes at the ML and especially their relationship
to heterozygosity. I first consider the case of a single
reinforces the patterns (appendix d).
diallelic ML and then extend to multiple loci. Heterozygosity-variance and heterozygosity-mean
cor-The mean phenotype of a genotype G (Equation 6), relations appear if genotypes at QTL and ML are
corre-and the within-class component of its variance [first lated. In infinite, partially selfing populations,
correla-term of (7)], can be found from Equations 4 and 5, tions among loci result from the mixing of different
respectively, replacing fkby fG(the average inbreeding
inbreeding classes. Within each class, genotypes at
un-coefficient of genotype G) and f2
k by fG2 (the average
linked loci are not correlated. Therefore, phenotypic
squared inbreeding coefficient of G), defined as mean and variance within a genotype G at the ML are
determined by the relative proportions of the different
fG5
o
kGkfk (8a)
inbreeding classes within G.
Within class k, genotypic frequencies at QTL i are
f2
G 5
o
kGkfk2. (8b)
AiAi AiBi BiBi
p2
i 1 fkpiqi 2piqi(12fk) q2i 1 fkpiqi
, (1) The phenotypic mean of G is therefore
φG5φ02 fG
o
i(2piqidi) (9)
where fk5121/2kis the usual inbreeding coefficient.
The mean and variance of phenotypic contributions
and the phenotypic variance, adding the among-class from QTL i within class k are therefore
to the within-class component, is
φki5 ai(pi2 qi)1diHki (2) s2
G5 s201fG(
o
i(4piqia2i)2VA2fGVD) and
1 s2
f(G)
o
io
j.i(8pipjqiqjdidj), (10)
s2
ki52piqi(11fk)ai21 di2Hki(1 2Hki)
wheres2
f(G)5 fG22 (fG)2.
22aidi(pi2qi)Hki, (3)
One marker locus: Let us consider a ML with two
where Hki 5 2piqi(1 2 fk). As the contributions from
alleles M and m of respective frequencies P and Q. At
various QTL are independent within a given inbreeding equilibrium, the frequency of (Mm) is 2PQ[12
s/(22
class, the phenotypic mean and variance of class k are s)] in the population (Hartl and Clark 1997) and
simply φk5 Riφki andsk25Ris2ki, respectively. With ref- 2PQ(1/2)k within class k. Therefore the frequency of
erence to their values for outbred individuals, φ0 and class k among Mm individuals is
s2
0, and to classical terms of additive and dominance
variance,
[Mm]k5
2PQ(1/2)k
sk
(1 2s)
2PQ[12s/(22s)] (11)
φk5φ02 fk
o
i(2piqidi) (4)
5 (s/2)k(12 s/2).
s2
k5 s202 fkVA 2fk2VD1fk
o
i(4piqia2i), (5)
[MM]k5 sk(12 s)
12Q/2k
12Q[12s/(22 s)] (12)
and a similar expression for [mm]k, replacing Q by P in
(12). From these frequencies we obtain
fMm5s/(42s) (13)
f2
Mm5
s(41s)
(42 s)(82s) (14)
fmm5
s[21s1 2Q(12s)]
(42 s)[s12Q(12s)] (15)
f2
mm5
s[8(11Q)16s(22 Q)1s2(1 22Q)]
(42 s)(82s)[s12Q(12s)] (16)
and similar expressions for MM, replacing Q by P in
(15) and (16). Clearly, f and f2are minimal for
heterozy-gotes and maximal for the rarest homozygote.
Expressions (9) and (10) allow any kind of
determina-Figure1.—Ratio of phenotypic variances (pooled
homozy-tion of the phenotype by QTL. We focus on three cases gotes/heterozygotes) for additive QTL and one marker locus
of interest: additivity, directional dominance, and sym- as a function of the allele frequency at that locus (P).
metric overdominance.
Additivity: The genetic variance is entirely additive
when di50 for all QTL. In this case (8) gives Dominance: Dominance at multiple loci is modeled
by setting di5aifor all i (complete dominance). In this
φk5
o
iai(pi2qi)5 φ0. (17)
case the phenotypic mean of class k is a monotonic function of k:
Therefore the phenotypic mean does not vary among
inbreeding classes and/or genotypes. The phenotypic φk5 φ02 fk
o
i
(2piqiai). (21)
variance within G reduces to
An interesting case is directional dominance, when
s2
G5(11 fG)s20, (18)
the phenotype decreases following homozygosity for
re-whereas the population variance is cessive alleles. This may apply to fitness traits
(deleteri-ous recessives) or to any morphological trait subject to
loss-of-function mutations (Roff1997). In this case, all
s25 2
22 ss
2
0. (19)
aiare positive and the mean phenotype decreases with
inbreeding.
Remarkably, the ratio s2
Mm/s2 5(42 2s)/(42s) is
The phenotypic variance within class k is a function of the selfing rate only. As this ratio is always
,1, the phenotypic variance in heterozygotes is always s2
k5 s201fk(
o
i(4piqia2i)2 VA 2fkVD) less than the population variance. The variance ratio
between MM and Mm is 5 s2
01fk
o
i(4piqia2i)(12 2q2i 2 fkpiqi). (22)
s2
MM/s2Mm5
(42s)24Q(12s)
(422s) 24Q(12s). (20) When inbreeding k increases, the contribution of
lo-cus i to the change in phenotypic variance is
propor-From (20), the variance of homozygotes exceeds that tional to fk(122qi22f
kpiqi), an increasing function of
of heterozygotes by a maximal factor of 1.5. The differ- k for pi . 0.5. When the phenotype is a fitness trait,
ence is more pronounced for rare homozygotes as (20) recessive alleles are rare (pi ≈ 1), and the variance
in-increases with Q. creases with inbreeding.
The variance of heterozygotes is often compared to Phenotypic mean and variance for a genotype G are
that of pooled homozygotes (MM1mm) (Eanes1978;
φG5 φ02 fG(
o
i2aipiqi) (23)
Mitton1978;King 1985), which is simply a weighted
average ofs2
MM ands2mmas both have the same
pheno-and typic mean. The variance ratio between homozygotes
and heterozygotes is plotted in Figure 1. This ratio, for s2
G5 s201 fG
o
i[4a2
ipiqi(122q2i)]
a given s, varies little with the allelic frequency at the
ML and reaches its maximum value (11 s/2) at P5 2
(fG)2(
o
i2aipiqi)21 8f2G
o
io
j.iaiajpipjqiqj. (24)
As phenotypic means are negative linear functions
of fG, heterozygotes have the highest (and rare
homo-zygotes the lowest) phenotypic mean. This is the
well-known “associative overdominance” phenomenon (Ohta
andCockerham1974;Charlesworth1991). Pheno-typic means under partial selfing were previously
de-rived byHoule(1994), using the adaptive distance
for-malism invented by Smouse (1986). Mean fitness was
predicted to be a negative, linear function of adaptive distances (defined as 0, 1/P, and 1/Q for genotypes
Mm, MM, and mm, respectively). Houle’s (1994) ap-proximations included discrete inbreeding depression (the fitnesses of inbreds and outbreds are constants) and low selfing (negligible recurrent inbreeding). The present results allow for a variance in fitness among inbreds and among outbreds, arbitrary values of s, and
recurrent inbreeding. For small s, fGand adaptive
dis-tances are linearly related, in agreement with Houle
(1994).
Phenotypic variances also vary among genotypes. As-suming, for simplification, directional dominance, iden-tical allele frequencies (p, q), and phenotypic effect (a) for all QTL (24) becomes
s2
G5 s20 11
1
q(11 q)[fG(12 pq fG22q
2)
1pq(L 21)s2
f(G)]
. (25) zygotes (MMFigure2.—The ratio of phenotypic variance among homo-1 mm) to the variance among heterozygotes
(Mm) at a marker locus, with directional dominance. The
With small frequencies of recessives (q , 0.5), the graphs are given for two values of q, the frequency of recessive
alleles, and four values of L, the number of loci that contribute
variance increases with fG and s2f (G), both minimum
to phenotypic variation. The allele frequency at the marker
for heterozygotes (Mm), except for very large s. Figure
locus has little effect on variance ratios (data not shown) and
2 shows that for s,0.75, the variance among
homozy-is set at P50.5.
gotes exceeds that among heterozygotes, sometimes considerably (ratio of 3, with many QTL and/or rare recessive alleles).
s2
hom/s2het5
31(12 s)(124PQ)
11(12 s)(124PQ). (27)
So far, selection at QTL has been neglected. However, correlations between the phenotype and fitness have
two consequences: (i) The equilibrium frequency of This ratio stays between 2 and 3 (when P 5 0.5) in
deleterious alleles is small and (ii) it varies across in- agreement with Figure 2.
breeding classes, as purging selection occurs following The consequences of (ii) are complex. Although the
repeated selfing; (i) means that we need consider only computations assume constant allelic frequencies,
dele-the bottom part of Figure 2 (q50.01). In this case, the terious alleles decrease in frequency in selfed lineages.
homozygote-to-heterozygote ratio of phenotypic vari- However, this effect is small (appendix b).
ances takes high values (near 3) over a large range of Symmetric overdominance: Overdominance occurs
s and L. Indeed, assuming very small qi’s (relative to 1/ when di.|ai|. I focus on the simplest case of symmetric
L and s), the variance becomes proportional to the overdominance, i.e., ai50 for all i. Although this looks
average inbreeding level of G: unrealistic, the discrepancy with real situations may be
small for two reasons. First, very asymmetrical
overdomi-s2
G≈4fG
o
iqia2i. (26) nant polymorphisms are usually instable with partial
selfing (KimuraandOhta1971, pp. 190–196). Second,
This agrees withDavid’s (1997) prediction that the asymmetry results in extreme allele frequencies (near
variance in fitness traits increases linearly with adaptive 0 or 1) when a stable equilibrium exists. Such loci
con-distance for small s. Indeed, adaptive con-distances are lin- tribute little genetic variance.
early related to fGin this case. More generally (for any With symmetric overdominance, phenotypic means
s), the homozygote-to-heterozygote ratio in phenotypic decrease with inbreeding (Equation 4), and so do vari-ances:
s2
k5(1 2fk)(s021fkVD). (28) zygosity being retained in inbred lines than predicted
under neutrality. This introduces little change to the This is expected as heterozygotes for QTL are
progres-above conclusions (appendix c).
sively eliminated from increasingly inbred classes; only
Several marker loci:Most studies on
heterozygosity-homozygotes eventually remain, which have
homoge-variance relationships use several ML (typically 5–10; neous phenotypes.
Zouroset al. 1980;Yezerinacet al. 1992;Livshitsand The mean phenotype of genotype G (Equation 4,
Smouse1993;Bamshadet al. 1994;Davidet al. 1997).
replacing fkby fG) is maximal for heterozygotes (Mm)
As the number of possible genotypes increases rapidly and minimal for rare homozygotes. The variance is
(3Mwith M diallelic loci), multilocus genotypes are
usu-ally pooled into heterozygosity classes. The phenotype
s2
G5 s202fGVA2fG2VD1 s2f(G)(
o
i2piqidi)2. (29)
and its variance (or absolute deviation from the mean) are then regressed on multiple-locus heterozygosity Although the within-class component (three first terms)
(MLH). is maximized in heterozygotes, the among-class
compo-The general expression of phenotypic variance
nent (fourth term) is usually (for s, 0.8) minimized
(Equation 10) still applies for multilocus genotypes or in heterozygotes. Heterozygosity-variance relationships
MLH classes. Regressions of phenotypic mean and
vari-depend on L, the number of QTL. With L5 1, the
ance on MLH are completely determined by f(h) and
variance increases with heterozygosity. However, for
large L, the among-class component dominates the f2
(h), the average inbreeding coefficient and average
within-class one, and the variance decreases with hetero- squared inbreeding coefficient of class h, respectively.
zygosity. In the simple case of M ML with identical genetic
diver-Overdominance is mainly cited for fitness traits, and sity H0(5 2PQ), they turn out to be
selection must be taken into account. The first
conse-quence of overdominant selection is to keep allele fre- f
(h)5
s u(h,s)
o
M2h
i50
Ci M2h
(22H0)i (2h1i2
s)(2h1i112s) (31)
quencies constant. With symmetric overdominance, pi
and qiapproach 0.5 (VA≈0). Assuming constant
locus-and
specific effects (di5 d for all i’s), we obtain
s2
G5 s20[1 2( fG)2 1(L2 1)s2f(G)]. (30) f(h)2 5 s
u(h,s)
o
M2h
i50
Ci M2h
The homozygote-to-heterozygote ratio of phenotypic
variances depends on both s and L (Figure 3). In the · (22H0)i(2h1i111s)
(2h1i2s)(2h1i112s)(2h1i12 2s), (32)
usual range of s (0–0.3), the ratio is very slightly ,1
with one QTL. However, with two or more QTL, it is
where u(h,s) 5 RM2h
i50CiM2h((22H0)i/(2h1i2 s)) and
.1 and becomes quite large when L increases.
Cp
n5 n!/p!(n2 p)!.
Purging selection does not occur with
overdomi-Equations 31 and 32 are decreasing functions of h. nance. However, overdominance results in more
hetero-As expected from single-locus results, the relationship between heterozygosity and phenotypic mean is absent in the additive case and positive in the dominant and overdominant cases. Heterozygosity-variance relation-ships are negative under additivity and under domi-nance (Figures 4 and 5). Under overdomidomi-nance (Figure 6), this relationship may be positive, negative, or present a maximum at intermediate MLH values, depending on
L and s. The most realistic situation for empirical studies
is low s and L @ 1, in which case the relationship is
predominantly negative.
In all cases the differences in variance are higher than in the single-locus case, because a larger range of f and
f2is explored. The orders of magnitude depend on the
determinism. For low s (0.05–0.1), the variance ratio between extreme MLH classes is of the order 1–2 under additivity, 5–10 under overdominance, and 30–60 under dominance. In all cases, heterozygosity-mean and het-Figure3.—The ratio of phenotypic variance among homo- erozygosity-variance relationships are nonlinear. An
as-zygotes (MM 1 mm) to the variance among heterozygotes ymptote exists for high MLH as very heterozygous
geno-(Mm), with symmetric overdominance. The graphs are given
types contain no inbreds, and their phenotypic mean
for four values of L, the number of loci that contribute to
and variance approach those of a random-mating
popu-phenotypic variation. The allele frequency at the marker locus
Figure 4.—Relationship between heterozygosity at 10 marker loci (each with H05 0.5) and phenotypic variance
under additivity. The phenotypic variance is in units ofs2 0,
the phenotypic variance in the absence of inbreeding.
DISCUSSION
Our aim was to derive the relationships between het-erozygosity at neutral marker loci and the mean and variance of arbitrary phenotypes in infinite, partially inbred populations. In this section, I focus on three questions. First, how do the results on phenotypic means compare with and/or extend previous results? Second,
Figure5.—The change in mean (A) and variance (B) of
what are the main conclusions regarding heterozygosity- a phenotype under dominance with increasing heterozygosity
variance relationships? Third, what is their relevance to at 10 marker loci (each with H050.5). A is valid for arbitrary
empirical results? number, locus-specific effects, and allele frequencies of QTL.
The phenotypic mean under random mating (φ0) has been The relationship between heterozygosity and
pheno-arbitrarily set to 0 and the graphs are in units of R(2piqiai)
typic mean in partially inbred populations:This model
(see Equation 23). B is represented for 100 QTL loci with
explicitly relates the phenotypic mean with fG, the aver- identical effects on phenotype and allele frequency q50.01.
age inbreeding level of a genotype. This relationship is Graphs are in units of s2
0, the phenotypic variance under
linear, resulting in a simple prediction: whenever a trait random mating. Range of s values as in Figure 4.
is negatively affected by inbreeding (through overdomi-nance, directional domioverdomi-nance, or both), phenotypic
means at ML rank as heterozygote.common
homozy-loci byDavid(1997)] relied on restrictive assumptions:
gote.rare homozygote. This has long been established
small s and discrete inbreeding depression, neglecting
by simulations (Charlesworth1991;Pamiloand
Pals-the variance arising from Pals-the detailed genetic
architec-son1998) or analytically (OhtaandCockerham1974;
ture of the trait. This study agrees with previous results,
Houle1994), and has been repeatedly suggested as the
as the fitness predictors derived byHoule(1994;
adap-probable origin of heterozygosity-fitness correlations
tive distances) andDavid(1997;gcoefficients) are
lin-(Houle1989;Davidet al. 1997). However, alternatives,
early related with fG in their domain of applicability.
based on direct overdominance (see Mitton and
However, fGis more general, accounting for recurrent
Grant1984) or multiple-locus dominance (Dengand
inbreeding and arbitrary genetic architecture.
Fu1998), have also been cited. The occurrence of
posi-One of the predictions obtained using fGis that
hetero-tive heterozygosity-fitness correlations, resulting from
zygosity-phenotype relationships are nonlinear. They partial inbreeding, has indeed been referred to as a
saturate because very heterozygous genotypes are almost
general effect (Davidet al. 1995), a special case of the
exclusively outbred. Although intuitively predictable,
associative overdominance hypothesis (Ohta 1971;
this feature is usually ignored as most empirical studies
Zouros1993). The other case is the local effect arising
use linear regressions. Of course, individuals with ex-from linkage disequilibrium in small populations (see
treme MLH are rare and very likely lost from a sample
David1998 for review and definitions).
of reasonable size. However, saturation at high MLH This study extends previous results, providing a
sim-values is visually perceptible in some instances (Strauss
ple method (the computation of fG) to predict
genotype-1986;Davidet al. 1997).
phenotype relationships under partial inbreeding.
pheno-types evenly distributed among classes have higher vari-ance. The relative importance of (i) and (ii) depends on the genetic architecture.
Very generally, heterozygotes appear to have lower variance than homozygotes in partially inbred popula-tions. This applies to phenotypes coded by additive or directionally dominant alleles as well as by overdomi-nant alleles in large regions of the parameter space including the most realistic situations. Additivity is ex-pected for neutral phenotypes, directional dominance and/or overdominance being more common for fitness
traits (Roff1997). Negative heterozygosity-variance
re-lationships have been predicted by Chakraborty
(1987) in large, random-mating populations, but only when the marker genes themselves have additive effects on the phenotype (a quite unlikely situation). In non-random mating populations, negative heterozygosity-variance relationships have been predicted for fitness
traits by David (1997) but with restrictive conditions
(see above), and without evaluating the influence of the genetic architecture, the number of genes involved, and the inbreeding rate.
The genetic architecture largely affects the relation-ships. Directional dominance results in large differences in variance among genotypes, one order of magni-tude higher than the differences under overdominance, which in turn are larger than under additivity. Although details of genetic architecture are usually unknown, a broad consequence is that the influence of heterozygos-ity on variance will be larger for traits highly correlated with fitness (mostly under dominant or overdominant gene action) than for traits loosely or not correlated with fitness.
Figure 6.—Heterozygosity at 10 marker loci (each with The number of phenotype-coding loci (QTL) affects
H0 5 0.5) and phenotypic variance under symmetric over- the relative importance of the within- vs. among-class
dominance for three numbers of overdominant QTL: (A)
components of variance. When many genes affect a trait,
L51; (B) L510; and (C) L5100. Range of s values as in
the stochastic variation in the overall sum of their effects
Figure 4.
is buffered, and all classes have low phenotypic vari-ance. Increasing the number of QTL (L) thus reinforces the among-class component of variance compared to typic variance in partially inbred populations:Until now,
the within-class component [see (25) and (30)]. This changes in phenotypic variance with inbreeding have
effect is nonexistent under additivity (the among-class been mostly envisaged in artificial, homogeneously
in-component is zero) and weak for dominant fitness traits
bred lines (Robertson 1952; Falconer andMackay
1996). Recently, it has been recognized that the pat- [because of low frequencies of recessives—see (25)].
tern of variation of mean and variance of fitness traits However, it is important under overdominance.
An-upon controlled inbreeding provides information on other consequence under overdominance is that when
the underlying determinism such as selection coeffi- enough polymorphic marker loci are considered, the
cients and dominance (Deng1998). This method was variance no longer decreases uniformly with
heterozy-designed for either outcrossing or selfing populations gosity. However, with an average scoring effort (10
poly-at mutpoly-ation-selection equilibrium. However, npoly-atural
morphic loci or less), this is perceptible only in largely populations are often heterogeneous because of partial
inbred populations. inbreeding. This generates genotype-phenotype
cor-Inbreeding rate also influences the patterns. When s relations based on the distribution of genotypes among
increases, the decrease in variance with heterozygosity various inbreeding levels. These distributions determine
is steepened under additivity and dominance. However, phenotypic variances, because (i) inbreeding classes
under overdominance, increasing s rather tends to shift have different variances (within-class component),
the maximum variance toward more heterozygous therefore genotypes concentrated in high-variance
classes (Figure 6), although heterozygosity-variance re-classes have higher variance and (ii) re-classes have
Moreover, the initial increase may be overlooked be-cause very homozygous classes are often pooled to-gether. Although higher s may yield variable outcomes, highly selfing species are rarely examined, being essen-tially devoid of heterozygosity and inbreeding
depres-sion (CharlesworthandCharlesworth1987).
The relevance of the model to empirical studies in
natural populations:This study involves usual
assump-tions of quantitative genetics and inbreeding depression models. The assumption of homogeneity in locus-spe-cific effects is unrealistic. However, mixtures of different
Figure7.—Relationship between heterozygosity at nine
al-determinisms have not been studied because of the
lozyme loci and variance in growth, measured as t1/2, the age number of situations to explore (Equation 10 provides
at half the maximum size in years, in the marine bivalve Spisula
a general formula). Negative heterozygosity-variance
re-ovalis (seeDavidet al. 1997). Classes MLH50 and MLH5
lationships are expected in mixed situations, because 1 have been pooled to increase sample sizes. Error bars are
each determinism separately produces such relation- 95% confidence intervals (slightly underestimated due to the
nonnormality of distributions). The dotted line indicates
theo-ships. The assumption of linkage equilibrium is not
retical predictions under partial inbreeding and directional
significantly violated in large populations, such as
ma-dominance (Equation 25) with L5100 and q50.01. A value
rine bivalves or trees (often used in empirical studies; of s50.05 has been chosen because this species has separate
cf.Britten1996), even if QTL may be physically linked. sexes and natural levels of inbreeding must be small. The
The main difference between our model and actual number of marker loci has been set to the observed value (9)
and, for simplicity, all loci were assumed to have identical
situations lies with the environmental variance VE. The
heterozygosity (Equations 31 and 32) equal to the geometric
model considers only genetic variance, although all
mean of observed values (geometric rather than arithmetic
traits display various levels of nongenetic variation. VE was used as heterozygosities interact multiplicatively rather
will reduce relative changes in variance among geno- than additively in Equations 31 and 32). The graph has been
types, adding a constant to all. Furthermore, sampling scaled to the observed values by settings2
050.02. variance must also be considered. Extreme
heterozygos-ity classes have low sample sizes, hampering the
estima-tribution of genetic variance upon inbreeding are not tion of phenotypic variance. A robust procedure is to
easily distinguished. In practice, genotypic replicates, pool extreme heterozygosity classes up to reasonable
such as clonemates, are needed. Within-clone variance
sample sizes (King1985;Davidet al. 1997).
increased in inbred lines of the facultative parthenogens Despite these potential problems, negative
heterozy-Daphnia pulex and D. pulicaria (Deng1997), suggesting gosity-variance relationships have been repeatedly
ob-that inbreeding enhances developmental instability. Yet served in natural populations (see references in
Intro-YampolskyandScheiner(1994) found no relationship duction). Most of them have been interpreted as
between MLH and within-clone variance in D. magna, evidence for a direct role of enzyme heterozygosity in
either because natural inbreeding is too low or because maintaining developmental stability without reference
the results on pulex and pulicaria do not extend to magna. to a possible genetic variation of the traits. As shown by
The two sides of a given individual are often used as our model, partial inbreeding is a plausible alternative,
substitutes for clonemates (e.g.,Learyet al. 1983).
How-especially for fitness traits. This is illustrated by a data
ever, the overall evidence for negative correlations be-set on heterozygosity at nine enzyme loci and growth
tween heterozygosity and fluctuating asymmetry ap-in the bivalve Spisula ovalis (Figure 7, modified from
pears weak (Britten 1996). In conclusion, partial
Davidet al. 1997). Although the similarity between
ex-inbreeding can generate MLH-variance relationships by perimental and theoretical graphs obviously depends on
two nonexclusive ways: altering developmental stability an adequate choice of parameters, the observed pattern
and redistributing genetic variance. The former relies (shape of the heterozygosity-variance relationship,
or-on assumptior-ons whose generality and quantitative im-ders of magnitude) is consistent with the inbreeding
portance remain to be properly established. The latter hypothesis under dominance. In this instance,
overdom-always occurs, and, in a variety of realistic situations, inance or additivity cannot generate effects of the
ob-may produce detectable patterns qualitatively and quan-served magnitude.
titatively similar to empirical results obtained to date. Although the redistribution of genetic variance upon
I thank P. Jarne for helpful comments on the manuscript.
inbreeding is sufficient to explain large changes in va-riance with MLH, developmental stability could also vary. Even if direct effects of allozymes on
developmen-LITERATURE CITED
tal stability might be negligible, inbreeding itself may
Bamshad, M., M. H. Crawford, D. O. O’RourkeandL. B. Jorde,
influence developmental stability by increasing
non-1994 Biochemical heterozygosity and morphologic variation in
genetic variance (VE;Lerner 1954;FalconerandMac- a colony of Papio hamadryas hamadryas baboons. Evolution 48:
1211–1221.
redis-Bennett, J. H.,andF. E. Binet,1956 Association between mende- heterozygosity, growth rate, and developmental homeostasis. Annu. Rev. Ecol. Syst. 15: 479–499.
lian factors with mixed selfing and random mating. Heredity 10:
51–56. Mitton, J. B.,andR. K. Koehn,1985 Shell shape variation in the
blue mussel, Mytilus edulis L. and its association with enzyme Booth, C. L., D. S. WoodruffandS. J. Gould,1990 Lack of
signifi-cant associations between allozyme heterozygosity and pheno- heterozygosity. J. Exp. Mar. Biol. Ecol. 90: 73–80.
Ohta, T.,1971 Associative overdominance caused by linked detri-typic traits in the land snail Cerion. Evolution 44: 210–213.
Britten, H. B.,1996 Meta-analyses of the association between multi- mental mutations. Genet. Res. 18: 277–286.
Ohta, T.,andC. C. Cockerham,1974 Detrimental genes with par-locus heterozygosity and fitness. Evolution 50: 2158–2164.
Chakraborty, R.,1987 Biochemical heterozygosity and phenotypic tial selfing and effects on a neutral locus. Genet. Res. 23: 191–200. Pamilo, P.,andS. Palsson,1998 Associative overdominance, het-variability of polygenic traits. Heredity 59: 19–28.
Chakraborty, R.,andN. Ryman,1983 Relationship of mean and erozygosity and fitness. Heredity 81: 381–389.
Robertson, A.,1952 The effect of inbreeding on the variation due variance of genotypic values with heterozygosity per individual
in a natural population. Genetics 103: 149–152. to recessive genes. Genetics 37: 189–207.
Roff, D. A.,1997 Evolutionary Quantitative Genetics. Chapman & Hall, Charlesworth, D.,1991 The apparent selection on neutral marker
loci in partially inbreeding populations. Genet. Res. 57: 159–175. London.
Smouse, P. E.,1986 The fitness consequences of multiple-locus het-Charlesworth, D.,andB. Charlesworth,1987 Inbreeding
de-pression and its evolutionary consequences. Annu. Rev. Ecol. erozygosity under the multiplicative overdominance and inbreed-ing depression models. Evolution 40: 946–957.
Syst. 18: 237–268.
David, P.,1997 Modeling the genetic basis of heterosis: tests of Strauss, S. H.,1986 Heterosis at allozyme loci under inbreeding and crossbreeding in Pinus attenuata. Genetics 113: 115–134. alternative hypotheses. Evolution 51: 1049–1057.
David, P.,1998 Heterozygosity-fitness correlations: new perspectives Strobeck, C.,1979 Partial selfing and linkage: the effect of a heter-otic locus on a neutral locus. Genetics 92: 305–315.
on old problems. Heredity 80: 531–537.
David, P., B. Delay, P. BerthouandP. Jarne,1995 Alternative Weir, B. S.,andC. C. Cockerham,1973 Mixed self and random mating at two loci. Genet. Res. 21: 247–262.
models for allozyme-associated heterosis in the marine bivalve
Spisula ovalis. Genetics 139: 1719–1726. Yampolsky, L. Y.,andS. M. Scheiner,1994 Developmental noise, phenotypic plasticity, and allozyme heterozygosity in Daphnia. David, P., B. DelayandP. Jarne,1997 Heterozygosity and growth
in the marine bivalve Spisula ovalis: testing alternative hypotheses. Evolution 48: 1715–1722.
Yezerinac, S. H., S. C. LougheedandP. Handford,1992 Morpho-Genet. Res. 70: 215–223.
Deng, H.-W.,1997 Increase in developmental instability upon in- logical variability and enzyme heterozygosity. Individual and pop-ulation level correlations. Evolution 46: 1959–1964.
breeding in Daphnia. Heredity 78: 182–189.
Deng, H.-W.,1998 Estimating within-locus nonadditive coefficient Zink, R. M., M. F. SmithandJ. L. Patton,1985 Associations be-tween heterozygosity and morphological variance. J. Hered. 76: and discriminating dominance versus overdominance as the
ge-netic cause of heterosis. Gege-netics 148: 2003–2014. 415–420.
Zouros, E.,1993 Associative overdominance: evaluating the effects Deng, H.-W.,andY.-X. Fu,1998 Conditions for positive and negative
correlations between fitness and heterozygosity in equilibrium of inbreeding and linkage desequilibrium. Genetica 89: 35–46. Zouros, E., S. M. Singh andH. E. Miles, 1980 Growth rate in populations. Genetics 148: 1433–1440.
Eanes, W. F.,1978 Morphological variance and enzyme heterozygos- oysters: an overdominant phenotype and its possible explana-tions. Evolution 34: 856–867.
ity in the monarch butterfly. Nature 276: 263–264.
Falconer, D. S.,andT. F. C. Mackay,1996 Introduction to
Quantita-Communicating editor:P. D. Keightley tive Genetics, Ed. 4. Longman, Harlow, United Kingdom.
Gardner, J. P. A.,1995 Developmental stability is not disrupted by extensive hybridization and introgression among populations of
the marine bivalve molluscs Mytilus edulis (L.) and M. galloprovin- APPENDIX A: APPROXIMATIVE DISTRIBUTION OF cialis (Lmk.) from South-West England. Biol. J. Linn. Soc. 54: INBREEDING LEVELS WITH SELECTION 71–86.
Without selection, inbreeding classes 0, 1, . . . , k have
Handford, P.,1980 Heterozygosity at enzyme loci and
morphologi-cal variation. Nature 286: 261–262. frequencies (12s), s(12s), . . . , sk
(12s). However,
Hartl, D. L.,andA. G. Clark,1997 Principles of Population Genetics.
mean fitness wkusually decreases when inbreeding (k)
Sinauer Associates, Sunderland, MA.
increases. Therefore the equilibrium distribution of k
Hill, W. G.,andA. Robertson,1968 Linkage desequilibrium in
finite populations. Theor. Appl. Genet. 38: 226–231. has lower frequencies of inbred classes (k.0) than the Houle, D.,1989 Allozyme-associated heterosis in Drosophila melano- neutral distribution. Therefore, the effect of selection
gaster. Genetics 123: 789–801.
resembles a decrease in selfing rate s to a lower value
Houle, D.,1994 Adaptive distance and the genetic basis of heterosis.
Evolution 48: 1410–1417. s9. Let us compute s9 values that provide reasonably Kimura, M.,andT. Ohta,1971 Theoretical Aspects of Population Genet- accurate distributions of k.
ics. Princeton University Press, Princeton, NJ.
Let the frequency of class k be lk. Assuming w05 1,
King, D. P. F.,1985 Enzyme heterozygosity associated with
anatomi-cal character variance and growth in the herring (Clupea harengus the change in thelk’s during one generation is
L.). Heredity 54: 289–296.
Leary, R. F., F. W. AllendorfandK. L. Knudsen,1983 Develop-mental stability and enzyme heterozygosity in rainbow trout.
Na-ture 301: 71–72.
1
l0
l1
… lk
…
2
t11
51 wt
1
(12s) (12s) … (12s) (12s)
sw1 0 0 0 0
0 … 0 0 0
0 0 swk 0 0
0 0 0 … 0
2
·
1
l0 l1 … lk …2
t , Lerner, I. M.,1954 Genetic Homeostasis. Oliver and Boyd, London.Livshits, G.,andE. Kobyliansky,1985 Lerner’s concept of devel-opmental homeostasis and the problem of heterozygosity level
in natural populations. Heredity 55: 341–353. (A1)
Livshits, G.,andP. E. Smouse,1993 Relationship between
fluctuat-or mfluctuat-ore concisely,
ing asymmetry, morphological modality and heterozygosity in an elderly Israeli population. Genetica 89: 155–166.
McAndrew, B. J., R. D. WardandJ. A. Beardmore,1982 Lack of L
t11 5 1
wt
S ·Lt, (A2)
relationship between morphological variance and enzyme hetero-zygosity in the plaice, Pleuronectes platessa. Heredity 48: 117–125.
Mitton, J. B.,1978 Relationship between heterozygosity for enzyme where w
tis the mean fitness at time t. The population
loci and variation of morphological characters in natural
popula-eventually reaches a steady state, the increase in k
tions. Nature 273: 661–662.
elimination of inbred individuals. The equilibrium q(k) in inbreeding class k. The deficiency in
heterozy-mean fitness w∞ is the real eigenvalue of S and the gous (AB) genotypes within class k compared to
random-distribution of k at equilibrium is the corresponding mating expectations is noted f(k). Without selection,
unitary eigenvector L∞. Although w∞ cannot be com- q(k)5q for all k, and f(k)5121/2k. However, if the
puted analytically, numerical values can be obtained, fitnesses of AA, AB, BB are 1, 1, 12z, respectively, and
restricting S to a reasonable dimension (say, kmax510) ignoring mutation, q(k) and f(k) change from class (k)
and setting values to the wk’s according to a selection to class (k1 1) (one generation of selection followed
model. L∞ can then be computed using lk,∞ 5 ((1 2 by selfing),
s)sk/wk∞11)
p
k j50wj.The approximation should conserve the crucial char- q(k11)5q(k) 211w(k)
w(k) (B1)
acteristics of the distribution of k for
heterozygosity-variance relationships, i.e., the mean and heterozygosity-variance of fk.
The former is f∞5 R∞k50lk,∞(1 21/2k) at equilibrium. f(k1 1)512[1 2f(k)] q(k)
2q(k1 1), (B2)
We need only the first few terms for numerical estima-tions.
where w(k)5 12 z[q(k)21f(k)p(k)q(k)] is the mean
As the approximate distribution of k is constructed
fitness of class k. As expected, q(k) decreases when k
as the neutral distribution under a selfing rate s9, its
increases, and selection slows down the increase in f(k).
mean f is the classical s9/(22s9). This will be equal to
Although recurrent mutation must exist to maintain the
the true f∞provided s9 52f∞/(11f∞).
B allele, it only tends to restore neutral frequencies.
The accuracy of this approximation was tested under
Therefore, ignoring mutation leads only to
overestima-a simple multiplicoverestima-ative selection model: wk5(12 a)fk
tion of purging effects. (0, a ,1). s9was determined numerically as explained
Plugging (B1) and (B2) into (4) and (5) does not
above for a variety of s (0.05–0.8) anda (0–1) values.
yield a simple expression of phenotypic mean and
vari-Stronger inbreeding depression (a z 1) generates
ance within class k. I therefore obtained numerical
esti-larger reduction (s9 , s) in apparent inbreeding,
al-mates (iterating B1 and B2) and compared them to though the absolute value of s has little influence on
neutral predictions. The expected homozygote-to-het-this reduction (data not shown). The variance and third
erozygote ratio of phenotypic variances (s2
hom/s2het) was central moment of f for the approximate distribution
calculated numerically (neglecting inbreeding classes are compared to their numerical values under selection
with k.25) with and without selection. P was set at 0.5
in Table A1. The approximate distribution is generally
and q at 0.01. All combinations of three s (0.05, 0.1, very close to the actual one. The largest discrepancy
0.3) and three z (0.05, 0.1, 0.3) values were considered. is obtained in an unlikely situation (high inbreeding
The simulations without selection perfectly matched an-depression and high selfing rate).
alytical predictions (Equation 25). Selection reduced the ratio (s2
hom/s2het) by an amount ranging from 0.2%
(s5 0.05, z 5 0.05) to a maximum of 8.3% (s5 0.3,
APPENDIX B: THE CHANGE IN ALLELIC
FREQUENCIES AND HOMOZYGOSITY OF A z 5 0.3, an unlikely situation). The same parameter
DELETERIOUS ALLELE WITH INBREEDING combinations were tested with several QTL (L510 and UNDER PURGING SELECTION
100) in linkage equilibrium, and the effect of
selec-tion on (s2
hom/s2het) always remained within the same
Consider a single locus with two alleles A, B of
respec-tive zygotic frequencies p, q in the population, and p(k), range (0 to28.5%). Therefore purging selection does
TABLE A1
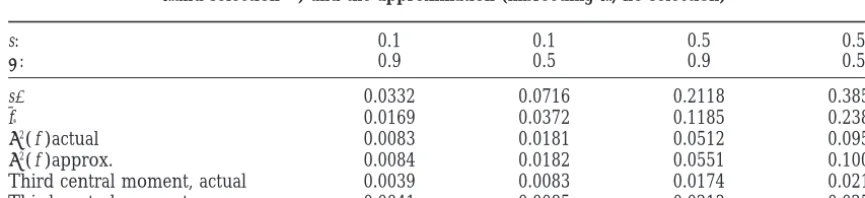
Comparison of the variance and third central moment offbetween the actual situation (inbreeding sand selectiona) and the approximation (inbreedings9, no selection)
s: 0.1 0.1 0.5 0.5
a: 0.9 0.5 0.9 0.5
s9 0.0332 0.0716 0.2118 0.3855
f∞ 0.0169 0.0372 0.1185 0.2387
s2( f )actual 0.0083 0.0181 0.0512 0.0958
s2( f )approx. 0.0084 0.0182 0.0551 0.1006
Third central moment, actual 0.0039 0.0083 0.0174 0.0217 Third central moment, approx. 0.0041 0.0085 0.0213 0.0252
not considerably affect heterozygosity-variance relation- additively. Thus, relationships between genotypes and phenotypic means [(4) and (9)] stay unchanged, ships.
provided ML and QTL remain unlinked.
Charles-worth (1991) predicted slight differences (a few
APPENDIX C: THE CHANGE IN HOMOZYGOSITY percent) in associative overdominance between un-AT SYMMETRICALLY OVERDOMINANT LOCI
linked and completely linked (l 51) QTL, probably
WITH INBREEDING
because QTL effects were considered multiplicative
Allele frequencies at symmetrically overdominant loci instead of additive and purging selection was not
are p5q51⁄
2, irrespective of inbreeding. Without selec- neglected. Linkage among QTL affects the
within-tion, heterozygosity is halved following every generation class component of variance more seriously,
of selfing. Under overdominance, the decrease in
het-s2
G(linked)5 s2G(unlinked) (D3)
erozygosity will be slower as heterozygotes have higher
viability than homozygotes (say, 11z for genotype AB 18
o
i
o
j.ipipjqiqjdidjhij,G,
and 1 for AA and BB),
where
f(k1 1)512[1 2f(k)](11z)
2w(k) , (C1) h
ij,G5
o
kGkhijk (D4)
where w(k)5 11 z[12 f (k)]/2 and f(k) is the zygotic
ands2
G(unlinked) is given by (10).
heterozygote deficiency within class k. As inappendix
From D3, linkage among QTL has no effect on
b, numerical simulations (up to k5 25) were used to
purely additive phenotypes (di50). However, it does
estimate the influence of selection as (C1) does not
affect heterozygosity-variance relationships under
di-yield a tractable expression of (s2
hom/s2het). Simulations
rectional dominance or overdominance. The exact using (C1) and parallel simulations using neutral f
distri-expressions for thehij,G’s, obtained from D4, D2, 11,
butions were performed for all combinations of three
and 12, are complicated. First-order approximations
s (0.05, 0.1, 0.3) and z (0.05, 0.1, 0.3) values, using 1, 2,
are
5, 10, and 50 QTL and P50.5. The neutral simulations
perfectly matched analytical predictions from (30). The
hij,Mm5 l2ijs/8 10(s2) (D5)
other results (not shown) indicate that (30) tends to
exaggerate absolute changes in variance between het- and
erozygotes and homozygotes. Thus, (s2
hom/s2het) is
under-hij,MM5[(22Q)/(12Q)]l2ijs/810(s2). (D6)
estimated when ,1 (one QTL) and is overestimated
when.1 (several QTL). However, the effect of selection
Therefore,hij,Gis positive and more than twice as
on (s2
hom/s2het) is always small (from11.3% for L5 1, large in homozygotes than in heterozygotes. It will
s50.3, z50.3; to26.5% for L550, s50.1, z50.3).
increase the difference between them. For fitness traits under directional dominance, this increase may
be negligible as qiand qjare small. However, under
APPENDIX D: THE EFFECT OF PHYSICAL LINKAGE
overdominance, with several QTL, the variance is
Physical linkage increases identity disequilibria in mainly sensitive to s2
f(G), which is of order s/8.
selfed lineages (Charlesworth1991), although it does Therefore, tight linkage among QTL (l
ij ≈ 1) may
not generate linkage disequilibria in infinite popula- appreciably increase the negative
heterozygosity-vari-tions. Two cases are envisaged: (1) linkage among QTL ance correlation.
and (2) linkage between ML and QTL. 2. I now consider linkage between ML and QTL.
Con-sider an indicator variable IG, which equals 1 for
indi-1. Heterozygosities at linked QTL are correlated within
viduals of genotype G and 0 otherwise. Because of a given inbreeding class. Consequently, phenotypic
physical linkage, IG and phenotypic values covary
contributions of loci i and j covary within class k by
within class k. This is quantified by two parameters, an amount
aG,k5covk(IG,φ)/p(G|k) andbG,k5covk(IG,φ2)/p(G|k),
Covk(φi,φj)54pipjqiqjdidjhijk. (D1) where cov
kmeans the covariance and p(G|k) the
fre-quency of G within class k. Introducing covariance
The identity disequilibriumhijkdepends on
inbreed-terms into (6) and (7) yields
ing (k) and linkagelij(51 minus twice the
recombi-nation fraction between i and j): φ
G5φG,unl1 aG (D7)
and
hijk5
1
11 l2
ij
4
2
k
2
1
14
2
k
(D2)
s2
G(φ)5 s2G,unl(φ)1 bG1 a2G2 2aGφG, (D8)
(WeirandCockerham1973). Covariation of QTL
(Equa-tions 6 and 7),aG5RkGkaG,k, andbG5RkGkbG,k. For ships are not affected when the phenotype is additive
a single QTL, we obtain (d 5 0). However, under dominance, the average
phenotypic difference between heterozygotes and
homozygotes increases. For small s and P5 Q51⁄
2,
aMm5
4pqd(22 s)sl2
(4 2s)[42s(11 l2)] (D9)
the increase is approximately proportional to (1 1
l2) (always,2), as inCharlesworth’s (1991)
nu-and
merical results. Similar effects are observed under
bMm 5daMm, (D10) overdominance. Moreover, under dominance or
overdominance, linkage also increases differences in
lbeing the linkage between the ML and the QTL.
variance between heterozygotes and homozygotes
Values for pooled homozygotes (MM 1 mm) are
(the last term of D8 being negative for the former
identical except for an opposite sign fora.