warwick.ac.uk/lib-publications
A Thesis Submitted for the Degree of PhD at the University of Warwick
Permanent WRAP URL:
http://wrap.warwick.ac.uk/109952
Copyright and reuse:
This thesis is made available online and is protected by original copyright.
Please scroll down to view the document itself.
Please refer to the repository record for this item for information to help you to cite it.
Our policy information is available from the repository home page.
Propylene O xide in M e th a n e -o x id isin g B a c t e r ia
by
Motoshl Suzuki
T his t h e s is i s p re s e n t e d fo r the d e g re e o f Doctor o f P h ilosop h y
Department o f B i o l o g i c a l Sciences
U n iv e r s ity o f Warwick
ACKNOWLEDGMENTS
SUNNARY
LIST OP FIGURES
LIST OF TABLES
CHAPTtat 1 G e n e r a l In trodu ction
1.1 The concept o f a e th y lo tro ph y
1.2 O ccu ren ce, ecology and h i s t o r y o f
th e i s o la t i o n o f a e t h a n e -o x id is in g b a c t e r i a
1.3
1.4
1 .4 .1
1 .4 .2
1 .4 .3
1 .4 .4
1 .4 .5
1 .4 .6
C l a s s i f i c a t i o n o f a e t h a n e -o x id is in g b a c t e r i a
P h y s io lo g y and b lo c h e a la t r y o f o b li g a t e a e t h a n e -o x id is in g b a c t e r i a
B a s i c growth re q u ir e s e n ts
C arb o n a e t a b o lis a
C^-coapound a s a i a ll a t i o n pathways
R lb u lo s e aonophosphate pathway (RKP)
S e r i n e pathway
R i b u lo s e diphosphate pathway o f C0_ a s s l a i l a t i o n
1.5
1
.
5 .21 .5 .3
1 .6
1.10
1.11
Methane oxid ation
TWo types o f aethane aonooxygenase. s o lu b le - a n d p a rt lc u la te -IM O
S o l u b l e aethane aonooxygenase
P a r t i c u l a t e aethane aonooxygenase
M ethanol o x id a tio n
G row th on aethanol and IMO a c t iv i t y
Secon dary a lc o h o l o x id a t io n
Formaldehyde o x id a tio n
F o rn a te oxid ation
The in d u s t r i a l a p p lic a t i o n s o f a e th y lo tro p h s
PAGE NO.
oxygenated compounds
22
1.13 I s »e th a n o l dehydrogenase a b le to su p ply
e le c t ro n s to the UNO? 27
1.14 The s t a b i l i t y o f b i o c a t a ly s t 28
1.14.1 The s t a b i l i t y o f the s o lu b le MMO 28
1.14.2 The s t a b i l i t y o f the p a r t ic u la t e MMO 31
1.15 In h i b it io n and I n a c t iv a t io n o f b io c a t a ly s t 32
1.15.1 I n h i b it io n o f NMO by c h e la t o rs 32
1.15.2 In h i b it io n o f c e l l growth by n i t r i t e 33
1.16 I n a c t iv a t io n o f NNO by ace tyle n e 34
1.17 I n a c t iv a t io n o f b i o c a t a l y s t by epoxides 36
1.16
I n a c t iv a t io n o f b i o c a t a l y s t by r a d i c a ls 381.19 S t a b i l i s a t i o n o f b i o c a t a ly s t 39
CHAPTER 2 M a te r ia ls and Methods
2.1
O rganisas 412.2
Media 412 .3 Maintenance and growth 41
2.4 Dry «»eight e stim atio n s o f c u ltu re s and
c e l l suspensions 44
2 .5 P re p a ra tio n s o f c e l l e x t r a c t s 44
2.6
Whole c e l l NNO a c t i v i t y 442.7 Methanol dehydrogenase a ssa y 45
2.8
Methane aonooxygenase a ssa y 462.9 Im m obilized c e l l - b l o f l l m re a c to r
(IB P -r e a c t o r ) 46
2.10
In a c t iv a t io n o f c e l l s u n der the co n d itio n so f P0 produ ction and I t s r e a c t iv a t io n 47
2.11
N i t r i t e determ ination 492.12
P r o t e in determ ination 502.13 Oases 50
CHAPTER 3
3 .1 .1
3 . 1. 2
3 .1 .3
3-2
3.2 .1 3.2 .2
3 .2 .3
3 .2 .*
3 -2 .5
3 .2 .6
3 -4 .1
3 -4 .2
3.*».3
3 .4 .4
3-5
3 -5 .1
The Produ ction o f Oxygen C on tain in g Compounds by M e th a n e -o x id isin g B a c t e ria
Reactions by s o lu b le MHO c e l l s 51
In tro d u c tio n
51
Experimental
52
R esu lts and d is c u s s io n
32
The e v a lu a tio n o f re a c tio n s and compounds
38
In tro d u c tio n
38
The produ ction o f prim ary a lc o h o ls
58
The produ ction o f secondary a lc o h o ls and
aethyIketones 62
The produ ction o f epoxides
63
The produ ction o f hydroxy-cyclic-com pounds
63
The produ ction o f aldehydes from halogensted
compounds and o th e rs 64
Comment 64
S e le c tio n o f t a r g e t compound and d is c u s s io n
66
The energy supply f o r the produ ction
o f PO 69
In tro d u c tio n
69
9
A lc oh ols as e le c t ro n donor
70
Methane as an e le c t ro n donor
73
Hydrogen a s an e le c t ro n d on or
75
D isc u ssio n
77
The c u lt iv a t io n o f h igh a c t i v i t y
MNO c e l l s
78
In tro d u c tio n
78
The vh o la c e l l MK) a c t i v i t y o f
aethanotrophs
8
lD isc u ssio n
8
*MonooxvKenase in Whole C e l l s
4.1 In t ro d u c tio n 91
<
1.2
The s t a b i l i t y o f c e l l s u n der the co n d itio n so f non-PO production 91
4 .2 .1 Experim ental 92
4 .2 .2 R e s u lt s 92
4 .2 .2 .1 C e ll s 92
4 . 2 . 2
.2
The e f f e c t o f tem peratures on the s t a b i l i t yo f p a r t ic u la t e MHO c e l l s . 92
4.3 The e f f e c t o f PO c o n c e n tra tion o f the growth
o f Nethylococcus c a p s u la tu s (Bath) and
M eth y lo cy stis parvus (OBBP) 94
4.4 I n a c t iv a t io n o f c e l l s u n d e r the c o n d itio n s o f
PO p ro du ctio n w ith PO accum ulation
97
4 .4 .1 Experim ental 99
4 .4 .2 R e s u lt s 99
4.5 D isc u ssio n 103
C hapter 5 In a c t iv a t io n o f Whole C e l l s under the
C on ditio n s o f PO P ro d u c tion without PO Accumulation
5-1 In t ro d u c tio n 109
5-2 The o ptim izatio n o f r e a c t i o n c o n d itio n o f
I B F -r e a c t o r H O
5-3 In t e r p r e t a t io n o f k i n e t i c s o f in a c t iv a t io n
on im m obilized c e l l - b l o f l l m r e a c to r 115
5 . * Use o f the IB F -r e a c t o r 116
5 .4 .1 Comparison o f peak p r o d u c t i v it y and whole
c e l l a c t i v i t y
116
5 .H .2 E f f e c t o f pH on the i n a c t i v a t i o n 118
5 .4 .3 E f f e c t o f temperature on th e in a c t iv a t io n 120
5 .4 .4 C assants on r e s u lt s co n ce rn in g the
X B F -reac tor 120
5-5 E f f e c t o f e x t r a c e ll u l a r PO con ce n tration
on the in a c t iv a t io n un der the co n d itio n s
5 - 5 .1 Experimental 123
5 - 5 - 2 R e s u lt s 124
5 - 6 D isc u ssio n 127
CHAPTER
6
The R ea ctiv ation o f C e l l s which havebeen In a c tiv a te d by a c e ty le n e or P ropylen e Oxide
6 .1 In tro d u c tio n 129
6 .2 The in a c t iv a t io n o f MHO u s in g ace tyle n e
and i t s r e a c t iv a t io n i n v iv o
130
6 . 2 . 1 In tro d u c tio n
130
6 . 2 . 2 The e f f e c t o f carbon s o u rc e s in the
r e a c t i v a t i o n process 131
6 . 2 . 2 . 1 Methane 131
6 . 2 . 2 . 2 Methanol 137
6 . 2 . 2 . 3 Formaldehyde 140
6 .2 . 2 . 4 Formate 140
6 .3 The e f f e c t o f oxygen on the re a c tiv a tio n
o f NMO 143
6 .4 The e f f e c t o f n itro g e n so u rc es on
r e a c t iv a t io n 143
6 .5 The e f f e c t o f su lph u r sou rc es 146
6 . 6 The e f f e c t o f tem perature on re a c t iv a t io n 146
6 .7 The re a c tiv a tio n o f c e l l s which are
in a c t iv a t e d by e x t r a c e l l u l a r PO 150
6 . 7 . 1 Experimental 152
6 . 7 . 2 R e s u lt s and d is c u s s io n 152
6 .8 Repeated r e a c t iv a t io n o f c e l l s fo llo w in g
in a c t iv a t io n 155
6 .9 P r o t e in s y st h e s is d u rin g the re a c t iv a t io n 155
6 .1 0 D isc u ssio n 158
CHAPTER 7
o f PO Production and i t s R e a c tiv a tion
the co n d itio n s o f PO produ ction 166
7 . 3 E f f e c t s o f pH, te a p e ra t u re and carbon
so u rc es on the r e a c t i v a t i o n o f p ro p y le n e -
in a c t iv a t e d c e l l s 177
7 - 3 . 1 The e f f e c t o f pH on the r e a c t i v a t i o n o f
p ro p y le n e -in a c tiv a te d c e l l s 177
7 - 3 . 2 The e f f e c t o f te a p e ra t u re on the
r e a c t i v a t i o n o f p r o p y le n e -in a c tiv a te d
c e l l s 177
7 . 3 . 3 The e f f e c t o f carbon sou rc es on the
r e a c t iv a t io n o f p r o p y le n e -in a c tiv a te d
c e l l s 177
7 .3 .* » The e f f e c t o f copper on the r e a c t iv a t io n
o f p r o p y le n e -in a c tiv a te d c e l l s 183
7 . 3 . * . l Experim ental 185
7 . 3 - * . 2 R e s u lt s and D isc u ssio n 185
CHAPTER
8
E lu c id a t io n o f In a c t iv a t io n and R e a c tiv a tio nMechanisms under the C on ditions o f PO Produ ction
8 . 1 In tro d u c tio n 191
8 . 2 PO con ce n tration i n the c e l l under the
c o n d itio n s o f PO produ ction 191
8 . 2 . 1 In tro d u c tio n 192
8 . 2 . 2 Experim ental 192
8 . 2 . 3 R e s u lt s and d is c u s s io n 193
8 . 3 P o s s i b le in a c t iv a t io n mechanisms o f MMO
by I n t r a c e ll u l a r PO 197
8 . 3 . 1 Experim ental 197
8 . 3 . 2 The e f f e c t o f methane on in a c t iv a t io n
o f c e l l s 197
8 . 3 . 3 The e f f e c t o f ammonia on in a c t iv a t io n 199
Î . 3 . * D isc u ssio n 199
8 . * P o s s ib le in a c t i v a t i o n mechanisms o f
co n cealed in a c t iv a t io n 203
8
. * . 3 . 18
. 4 .3.2
8
.
8.8
8
.8
.8.1
8 .5
CHAPTER 9
9
.
2 .19
.2.2
9 .2 .2 .1
9
.2
.2.2
9 .2 .3
9 -8
9
.8.1
9
.8.2
By-product form ation and in a c t iv a t io n
d u rin g the o x id a tio n o f propylene
205
How does P0 a c t as an in a c t i v a t o r w it h in the c e l l s under c o n d itio n s o f
h igh P0 production? 206
Experimented. 208
R e s u lt s and d is c u s s io n 208
The e f f e c t o f e le c t r o n donors on
in a c t i v a t i o n
212
Formaldehyde 212
Comments 119
The I n h i b it io n o f R e a c tiv a tio n
In tro d u c tio n ???
The e f f e c t o f n i t r i t e on in a c t iv a t io n
and r e a c t iv a t io n
2
??The e f f e c t o f n i t r a t e on r e a c t iv a t io n 222
The in h ib i t io n o f r e a c t i v a t i o n by n i t r i t e 223
Experim ental 223
R e s u lt s 223
The accumulation o f n i t r i t e 226
Experim ental 226
N i t r i t e accum ulation fo llo w in g the
in a c t iv a t io n
227
The e f f e c t s o f oxygen and an en e
rgy-supply on the accum ulation o f n i t r i t e
227
The e f f e c t o f n i t r i t e on the produ ction
o f P0 238
The in h ib i t io n o f r e a c t i v a t i o n by
h i s t i d i n e
236
I s r e a c t iv a t io n c o n t ro lle d ?
238
Experim ental 239
The e f f e c t o f methanol on the r e a c t i v a t i o n
o f MMO 241
9 .4 .4 D isc u ssion 243
CHAPTtjt 10 Process E valu atio n and the
Economics o f P0 Produ ction
10.1 In tro d u c tio n 247
10.2 S e le c t io n o f s u i t a b le o rg a n isa s 248
10.2 .1 In tro d u c tio n 248
10 .2 .2 Experim ental 230
1 0 .2 .3 R esu lts 230
1 0 .2 .3 .1 The r e a c t iv a t io n p a t t e r n s o f
th erm otolerant aethanotrophs 230
1 0 .2 .3 -2 The r e a c t i v a t i o n p a t t e r n s o f a e s o p h ilic
aethanotrophs
233
10.3 Which o rg a n is a i s b e t t e r as a b io c a t a ly s t
f o r the produ ction o f P07
236
10.4 The stoic h iom etry o f th e P0 p rodu ction and
methanol consumption
239
10.3 S u bstra te s consumption f o r r e a c t iv a t io n 264
10 .3 .1 In tro d u c tio n 264
10 .3 .2 Experim ental 264
10 .3 .3 The nec essa ry amount o f carbon source
f o r the r e a c t iv a t io n
263
10 .3 .4 The nec essa ry amount o f n itrogen
source f o r the r e a c t i v a t i o n 268
10 .3 .3 The nec essa ry amount o f sulphur source
f o r the r e a c t iv a t io n
271
10 .3 .6 D isc u ssio n 271
10.6 P rocess economics 273
10.6.1 The b i o l o g i c a l c h lo ro h y d rin p ro
cess-Cetus P ro cess
273
10.6 .2 The produ ction o f P0 u s in g the
g r o w i n g - c e lls process
277
10
.6
.2.2
Experimental1 0 .6 .2 .3 P0 produ ction under grow in g c e l l
co n d itio n s u sin g M eth y lo cy stis pai
1 0 .6 .2 .4 Process economics o f g r o w in g -c e ll
process
10.6.3 Flow sheet o f the tw o -st a g e
r e a c t iv a t io n process
10.6.4 The economics o f p ro du cin g P0
u sin g the tw o -stage r e a c t iv a t io n process
278
279
284
287
CHAPTER
11
General ConclusionR eferences
$
l
I should l i k e to thank s in c e r e ly ey s u p e r v is o r . P r o fe s s o r Howard
D alton f o r h i s encouragement and a d v ic e over the p a s t two y e a r s. I
would a l s o l i k e t o thank D r.S .H . S t a n le y , N r.A .O . R ich ards f o r the
experim ental a d v ic e and M r.J. Carney and M r.a. MacDonald f o r t h e ir
e x c e lle n t te c h n ic a l a s s is ta n c e and a l l o f the o t h e r members o f
M ic ro b io lo g y 1 a t B i o lo g i c a l S c ie n c e s over the l a s t '4 y e a r s .
F i n a l ly , I would l i k e to thank M r.T. Yamamoto, D r .T . Y o r l f u j l ,
M r.T. T etsu ka, M r.N. Murakami, M r.S . Kamata and a l l o f th e members o f
Biochem ical Research L aboratory and a ls o H E members f o r t h e i r support
The a l a o f t h is p r o je c t I s to develop a p ro c e ss f o r the production
o f p ro p y le n e o x id e (P O ), u s in g m eth an e-oxidisin g b a c t e r i a . A t the
b e g in n in g , a d i f f i c u l t p r o b le e needed to be s o lv e d f o r the development o f t h is p ro c e s s l . e . the s h o r t li f e - s p a n o f the b io c a t a ly s t . E x p e ria e n ts showed th at the c e l l s o f the e e t h a n e -o x id is in g bacterium, M ethylococcua capsulatu s (B a th ) l o s t t h e ir c a t a l y t i c a c t i v i t y w ith in 30
minutes un der the con d ition s o f h igh PO p rodu ction . The in a c tiv a tio n o f
the b i o c a t a l y s t was la r g e ly Independent o f exte rn a lly -a c c u m u la te d PO but was t o t a l l y dependent on PO produced in v iv o under c o n d itio n s o f high PO
p ro d u c tio n . The c e l l s l o s t t h e i r a c t iv i t y w ith ou t any e x te rn a l
accum ulation o f PO under those con d ition s where PO was produced. P r io r
to the re se a rc h o f the p re se n t w r i t e r , i t had been concluded that
e x t e rn a l PO in a c tiv a te d the b io c a t a ly s t . A s p e c i f i c PO p ro d u c tiv it y o f
■o re than 700 n ao l PO produced/aln/ag c e l l s was ob ta in e d in the work
re po rted h e r e . However, by in c r e a s in g the PO p r o d u c t iv it y more than 200
mU/mg c e l l s , the c e l l s l o s t t h e i r a c t iv i t y r a p i d ly and t h e ir h a l f - l i f e
la s t e d
7
m inutes.In o r d e r to overcome the s h o r t l i f e - s p a n o f the b io c a t a ly s t , a
r e a c t i v a t i o n o f the in a c tiv a te d c e l l s had to b e d e v is e d . The
m e th a n e -o x id isin g b a c t e r ia c o n t a in an enzyme, methane monooxygenase
(IM
0
) which o x id is e s methane to methanol and a ls o o x id is e s propylene toP0. The UNO was ir r e v e r s i b l y in a c tiv a te d by a c e ty le n e o r by P0, however
t h is in a c t i v a t e d MN
0
was r e a c t i v a t e d by s u b je c t in g the c e l l s tor e a c t i v a t i o n treatm ent. T h is r e a c t iv a t io n p ro c e ss i s a phenomenon not
p r e v io u s ly known about. I n o r d e r to r e a c t iv a t e the In a c tiv a te d MM0, the
c e l l s re q u ir e d carbon, n itro g e n and sulphur so u rc e s . In ad d ition , a
s u i t a b l e oxygen and temperature regime was re q u ir e d f o r the re a c tiv a tio n
p r o c e s s . The re q u ir emen t o f n u trie n t s fo r r e a c t i v a t i o n and the
in h i b i t i o n o f re a c tiv a tio n by th e ad dition o f chloram phenicol le d to the co n c lu sion t h a t p ro te in s y n th e sis was a sso c ia te d w ith the re a c tiv a tio n
p ro c e s s . Furthermore, i t was found that UNO s y n th e sis was completely
in h ib i t e d by a d e te c ta b le amount o f methanol in the c e l l suspensions. Copper was n o t re qu ired f o r the re a c tiv a tio n o f c e l l s which contained
p a r t i c u la t e NN
0
.Two ty p es o f in a c t iv a t io n mechanism were assumed under the
c o n d itio n s o f P0 production. These are the in a c t i v a t i o n o f NN0 and the
i n a c t i v a t i o n o f the b io c a t a ly s t by a means not y e t i d e n t i f i e d . When the
MIO o n ly was In a c tiv a te d , i t was re a c tiv a te d q u ic k ly . However when
th ese c e l l s w ere in a c tiv a te d un der con dition s o f h igh P0 production, they r e q u ir e d th ree times as lo n g a period f o r com plete re a c tiv a tio n
than d id th ose c e l l s which had been in a c tiv a te d by a c e ty le n e . This
d e la y in th e re a c tiv a tio n p ro c e s s was thought to be due to a concealed
i n a c t i v a t i o n (u n id e n t ifie d in a c t i v a t i o n ) fa c t o r . The l a t t e r was thought
to be caused by the accum ulation o f P0 w ith in the c e l l s . The
i n t r a c e l l u l a r P0 concentration was c a lc u la t e d on the b a s is o f the r e t e n t io n tim e o f P0 in the c e l l s , and i t s con c e n tra tion appeared to be
r e la t e d t o P0 p ro d u c tiv it y . The concealed in a c t i v a t i o n was assumed to
be due to a s o lv e n t -lik e e f f e c t o f P0 in the c e l l s and not from an a lk y la t i o n e f f e c t .
I n o r d e r to develop a P0 produ ction p ro cess u s in g the re a c tiv a tio n systam , H e th y lo c y stls parvus (0BBP) was s e le c te d as the b e s t organism
from 25 methanotrophs. The r e a c t iv a t io n system, the g r o w in g -c a ll
p ro c e ss ( s i n g l e sta g e ) and the tw o -sta g e r e a c t i v a t i o n p ro cess were
d esign e d and o pe rated. U sin g th e g r o w in g -c e ll p r o c e s s , continuous P0
p ro du ctio n was achieved a t a r a t e o f
12
g/l/da y f o r a p e rio d o f more2 .9 -1 The schematic diagram o f the IB F -r e a c t o r 1»8
3 -4 .1 The a c t iv e s i t e o f aethanol dehydrogenase 74
3 -5 .1 The e f f e c t o f co p per concentration s on
whole c e l l MMO a c t i v i t y
83
3 - 5 .2 The e f f e c t o f a e th a n o l con ce n tration on
whole c e l l MMO a c t i v i t y
87
3 -5 -3 The e f f e c t o f tem perature on whole c e l l
MMO a c t i v i t y
88
4 .2 .1 The s t a b i l i t y o f C e l l s under c o n d itio n s
o f non-PO p ro du c tio n
93
4 .2 .2 The s t a b i l i t y o f c e l l s under ana erobic
con d ition s
95
PAGE NO.
1 . 5 - 1 T h e o x i d a t i o n o f a e t h a n e b y m e t h a n o tro p h s 10
4 .2 .3 The decay o f whole c e l l a c t i v i t y d u rin g
s to ra g e in phosphate b u f f e r
96
4 .4 .1 In a c tiv a tio n o f c e l l s under the c o n d it io n
4 .4 .2 I n a c t iv a t io n o f c e l l s under the c o n d it io n
o f PO produ ction w ith PO accum ulation
High biom ass-m oderate c on ce n tration o f PO
accumulation
101
4 .4 .3 I n a c t iv a t io n o f c e l l s under the c o n d it io n s
o f PO p ro du ctio n w ith PO accum ulation
High b io m ass-h igh con ce n tration o f PO
accumulation
102
5 -2 .1 The time co u rse o f PO produ ction i n the
IB F -r e a c t o r
111
5 - 2 .2 The e f f e c t o f oxygen con ce n tration on
the peak p r o d u c t i v it y in the IB F -r e a c t o r 112
5 * 2 .3 The e f f e c t o f methanol c on ce n tration on
the peak p r o d u c t i v it y in the I B F -r e a c t o r 114
5 - 3 .1 The i n v i s i b l e I n a c t iv a t io n d u rin g PO
produ ction under lim it in g c o n d it io n s in
the IB F -r e a c t o r H
7
5 .4 .1 The e f f e c t o f pH on the in a c t i v a t i o n o f
5 . * . 2 The e f f e c t o f temperature on the
in a c t iv a t io n o f c e l l s in the I B F -r e a c t o r 121
5.1».3 The r e la t io n s h i p between peak p r o d u c t iv it y
and the time e la p se d u n t il the a c t i v i t y
decreased to h a l f o f the peak a c t i v i t y
122
5 .5 *1 The e f f e c t o f th e e xtern a l PO con c e n tra tion
on the i n a c t i v a t i o n o f c e l l s
125
5 .5 * 2 The e f f e c t o f th e e xtern a l PO con c e n tra tion
on the i n a c t i v a t i o n and on the p r o d u c t iv it y
o f the c e l l s 126
6 .2 .1 The e f f e c t o f methane as a carbon so u rc e
in the r e a c t i v a t i o n process
133
6 .2 .2 The e f f e c t o f methanol as a carbon source
in the r e a c t i v a t i o n process 134
6 .2 .3 The r e a c t i v a t i o n o f Hethylococcus
capsu latu s (B a t h ) c e l l s in which UNO was
com pletely in a c t iv a t e d by a c e ty le n e
135
6 .2 .4 The r e a c t i v a t i o n o f a c e t y le n e -in a c t iv a t e d
c e l l s by the endogenous energy in
6 .2 .5 The e f f e c t o f aethane con ce n tration in a i r
d u rin g the r e a c t i v a t i o n p ro cess in
Methylococcus cap su la tu s (B ath )
138
6 .2 .6 The e f f e c t o f aeth an ol on the r e a c t i v a t i o n
ra t e o f the in a c t iv a t e d c e l l s in
Methylococcus cap su la tu s (B ath ) 139
6 .2 .7 The e f f e c t o f fo ra ald eh y d e on the
r e a c t iv a t io n a s a carbon and energy
source in M ethylococcus capsu latu s (B a t h ) l 4 l
6 .2 .8 The e f f e c t o f f o r a a t e on the r e a c t i v a t i o n
as a carbon and energy source in
Methylococcus cap su latu s (B ath ) 142
6 .3 *1 The e f f e c t o f oxygen on the r e a c t i v a t i o n
o f MMO i n M ethylococcus c ap su latu s (B a th ) 144
6
.*1.1 The e f f e c t o f n itro g e n sources on ther e a c t iv a t io n o f MMO in Methylococcus
cap su la tu s (B a t h ) IU
5
6 .4 .2 The e f f e c t o f th e n itro g e n sources on
the r e a c t i v a t i o n o f NNO in M eth y lo cy stis
6 . 5 . 1
6
.
5.2
6
.6.1
6 .7 .1
6.7.2
6 .7 .3
6
.8.1
6 .9 .1 «
r e a c t iv a t io n o f MMO in Methvlococcus
capsu latu s (B a t h ) 148
The e f f e c t o f c y s te in e as a s u lp h u r so u rc e
on the r e a c t i v a t i o n o f MMO in Methylococcus
capsu latu s (B a t h ) 149
The e f f e c t o f tem perature on the r e a c t i v a t i o n
o f MMO in M ethylococcus c a p su la tu s (B a th ) 150
The r e a c t i v a t i o n o f the P O -in a c tiv a te d c e l l s
and both P 0 - and a c e t y le n e -in a c t iv a t e d c e l l s
o f M ethylococcus capsu latu s (B ath ) 153
The e f f e c t o f e x t e r n a l PO c o n c e n tra tion on
the in a c t i v a t i o n ' and the r e a c t i v a t i o n o f
c e l l s o f M ethylococcua cap su la tu s (B a th ) 154
The r e a c t i v a t i o n o f whole c e l l MMO a c t i v i t y
o f the P O -in a c tiv a te d c e l l s measured u s in g
d i f f e r e n t e le c t r o n donors
156
Repeated r e a c t i v a t i o n o f c e l l s f o llo w in g
in a c t iv a t io n b y ace tyle n e and e x t e r n a lly
added PO 157
P ro te in s y n t h e s is d u rin g the r e a c t i v a t i o n
o f MHO in M ethylococcus capsula t u s (B a th ) The e f f e c t o f t h e s u l p h u r s o u r c e o n t h e
6 . 9 . 1 b
7.2 .1
7 .2 .2
7 .2 .3
7 -2 . *
7 .2 .5
o f MHO i n M eth y lo cy stis parvu s (OBBP) l60
The r e a c t i v a t i o n o f c e l l s In a c tiv a te d
under the c o n d it io n o f PO p ro du ctio n -
low PO p r o d u c t i v it y ( I ) 168
The r e a c t i v a t i o n o f c e l l s In a c t iv a t e d
under the c o n d it io n o f PO p ro du ctio n
-low PO p r o d u c t i v it y ( I I ) 169
The r e a c t i v a t i o n o f c e l l s In a c t iv a t e d
under th e c o n d it io n o f PO p ro du ctio n -
moderate p r o d u c t iv it y
170
The r e a c t i v a t i o n o f c e l l s in a c t iv a t e d
under the c o n d it io n o f PO p ro du ctio n -
high PO p r o d u c t i v it y
171
The r e la t io n s h i p between r e s i d u a l
a c t i v i t y a t th e begin n in g o f the
r e a c t iv a t io n p ro c e ss and the time
re q u ir e d f o r complete r e a c t i v a t i o n
173
r a t e i n the e x p o n e t la l phase and the peak
p r o d u c t iv it y
17
*»7 * 2 .7 The r e a c t i v a t i o n o f c e l l s in a c t iv a t e d
under the c o n d it io n o f h igh PO p ro du c tio n
176
7 .3 .1 The e f f e c t o f pH on the r e a c t i v a t i o n o f
p ro p y le n e -in a c tiv a te d c e l l s I
78
7 * 3 .2 The e f f e c t o f temperature on the
r e a c t i v a t i o n o f propylene in a c tiv a te d
c e l l s o f Methylococcua cap au latua ( B a t h ) ,
a : r e a c tio n tem perature 45*C, b : r e a c t io n
temperature 40*C 179
7 .3 .3 The e f f e c t o f methanol as the carbon
source on the r e a c t iv a t io n o f p ro p y le n e
-in a c t iv a t e d c e l l s o f Methylococcua
c ap su latu s (B a t h ) 182
7 - 3 - 1» The e f f e c t o f peak p ro d u c tiv it y on the
l a g p e rio d observed d u rin g the i n i t i a l
p e rio d o f r e a c t i v a t i o n in Methylococcua
cap su la tu s (B a t h ) 183
7*3*5 The e f f e c t o f v a r io u s carbon so u rc es on
the I n i t i a l r e a c t i v a t i o n r a t e in
Methylococcua cap su la tu s (B ath ) 184
r e a c t i v a t i o n o f a c e t y le n e -in a c t iv a t e d
c e l l s o f M ethylococcus c a p su la tu s (B a th ) 186
7 - 3 - 7 The e f f e c t o f copper ions on the
r e a c t i v a t i o n o f p ro p y le n e -in a c tiv a te d
c e l l s o f Me th ylococcu s c a p su la tu s (B a th ) 187
7 - 3 - 8 The e f f e c t o f copper ions on the
r e a c t i v a t i o n o f a c e ty le n e -in a c t iv a t e d
c e l l s o f Me th y lo c y s t i s parvus (OBBP) 188
7 - 3 - 9 The e f f e c t o f copper ions on the
r e a c t i v a t i o n o f p ro p y le n e -in a c tiv a te d
c e l l s o f Me th y lo c y s t i s parvus (OBBP)
189
8 .2 .1 E stia a te d i n t r a c e l l u l a r PO con c e n tra tion
d u rin g the b io t r a n s fo r a a t i o n o f p ropylen e
195
8 .2 .2 S e qu e n tia l PO produ ction and th e change o f
e s t i a a t e d i n t r a c e l l u l a r PO c o n c e n tra tion
196
8 .3 - 1 The e f f e c t o f aethane on the i n a c t i v a t i o n
o f c e l l s un der con d ition s o f PO produ ction
i n the IB F -r e a c t o r
7 - 3 - 6 T h e e f f e c t o f c o p p e r i o n s o n t h e
o f c e l l s u n der conditions o f PO produ ction
in the I B F -r e a c t o r
8 - 3 -3 P o s s ib le ro u te s o f in a c t i v a t i o n o f MMO
a c t iv e s i t e by PO
8 .4 .1 In te rm itte n t re a c tio n and the in a c t iv a t io n
o f c e l l s o f Methylococcus c a p su la tu s (B ath )
8 .4 .2 The i n a c t i v a t i o n o f c e l l s f o llo w in g the
o x id a tio n o f ethylene in Methylococcus
c ap su latu s (B ath )
8 .4 .3 The i n a c t i v a t i o n o f enzymes under the
c o n d itio n o f PO production i n M ethylocytia
parvus (OBBP)
8 .4 .4 I n a c t iv a t io n o f enzymes by th e a d d itio n o f
ace ty le n e o r propylene o x id e and t h e ir
r e a c t i v a t i o n in M eth ylocystis parvus (OBBP)
8 .4 .3 The i n a c t i v a t i o n o f enzymes fo llo w in g the
a d d itio n o f h igh PO con ce n tration s and
t h e ir r e a c t i v a t i o n in M eth y lo cy stis parvus
8 . 3 - 2 The e f f e c t o f a u o n l a o n t h e i n a c t i v a t i o n
200
202
204
207
209
210
8 .4 .6 The i n a c t i v a t i o n o f enzymes fo llo w in g the
a d d itio n o f benzene and t h e i r r e a c t i v a t i o n
in M e th y lo c y stis parvus (OBBP) (1 ) 213
8 .4 .7 The In a c t iv a t io n o f enzymes fo llo w in g the
a d d itio n o f benzene and t h e i r r e a c t i v a t i o n
in H e th y lo c y s tis parvus (OBBP) (2 ) 214
8 .4 .8 The e f f e c t o f formaldehyde as an e le c t r o n
donor on th e in a c tiv a tio n o f Methylococcus
c ap su latu a (B ath ) in IB F -r e a c t o r 216
8 .4 .9 The i n a c t i v a t i o n o f c e l l s f o llo w in g the
a d d it io n i s formaldehyde and t h e ir
r e a c t i v a t i o n in Wethylococcus cap su latu a
(B ath ) 217
8 .4 .1 0 The i n a c t i v a t i o n o f enzymes fo llo w in g the
a d d it io n o f formaldehyde and t h e ir
r e a c t i v a t i o n in M eth y lo cy stis parvu s (OBBP) 218
8 .4 .1 1 The e f f e c t o f formate as an e le c t r o n donor
on the i n a c t i v a t i o n o f c e l l s i n the IB F
-r e a c t o -r
220
9 * 2 .1 The e f f e c t o f n i t r a t e con c e n tra tion on
r e a c t i v a t i o n in Hethylococcua cap su latu a
o f n i t r i t e and r e a c t iv a t io n in M eth y lo cy stis
parvus (OBBP) 228
9 .2 .3 a The accum ulation o f n i t r i t e f o llo w in g the
a d d it io n o f PO in M eth y lo cy stis p arvu s
(OBBP) 229
9 .2 .3 b The accum ulation o f n i t r i t e f o llo w i n g the
p ro du ctio n o f PO in M eth y lo cy stis p arvu s
(OBBP) 230
9 .2 .4 The accum ulation o f n i t r i t e f o llo w in g
the a d d i t i o n o f ace tyle n e under the
c o n d it io n o f low d is s o lv e d oxygen
c o n c e n tr a tio n s in M eth y lo cy stis p arvu s
(OBBP) 232
9 .2 .9 The accum ulation o f n i t r i t e fo llo w in g
the treatm en t o f high d is s o lv e d oxygen
c o n c e n tr a tio n con dition i n M e th y lo c y stis
parvus (OBBP) 233
9 .2 .6 The e f f e c t o f n i t r i t e on th e p ro du c tio n o f
PO in M ethylococcus c ap su latu a (B a th ) 235
9 .4 .1 The r e p r e s s io n o f r e a c t i v a t i o n MMO by
methanol i n M eth ylocystis p a r ''. - (OBBP) 240
9 .*.2
9 - * . 3
10.2.1
1 0.2 . 2
1 0 .2 .3
1 0 .2 .4
1 0 .3 -1
10.3 -2
o f MHO i n M ethylocystis p a rv u s (OBBP) 242
A d a p ta tion o f methane-grown c e l l s to the
methanol c u lt u r e by u sin g a c e ty le n e
-i n a c t -i v a t -i o n techn-ique 245
R e a c tiv a tio n o f a c e ty le n e -in a c t iv a t e d
c e l l s o f thermo to le ra n t methanotrophs 251
R e a c tiv a tio n o f p ro py le n e -in a c tiv a te d
c e l l s o f thermo to le ra n t methanotrophs
252
R e a c tiv a tio n o f a c e t y le n e -in a c t iv a t e d
c e l l s o f a e s o p h ilic methanotrophs 254
R e a c t iv a t io n o f p ro p y le n e -in a c tiv a te d
c e l l s o f m esoph ilic methanotrophs 255
The e f f e c t o f re action tem peratures
on the In a c t iv a t io n and r e a c t i v a t i o n
o f c e l l s 257
The e f f e c t o f re action tem peratures on
the in a c t i v a t i o n and r e a c t i v a t i o n o f
c e l l s I n Methylococcus c a p s u la tu s (B ath ) T he e f f e c t o f fo rm a te o n t h e r e a c t i v a t i o n
10 .4 .1
1 0 .5 .1
1 0 .5 .2
1 0 .5 .3
10 .5 .4
1 0 .5 .5
10.5.6
The s t o ic h lo a e t r y o f the b u t a n o l consumption
and P0 p ro du c tio n in M e th y lo c v stis parvus
(0B8P) 2*3
The n e c e s sa ry amount o f methanol f o r the
r e a c t i v a t i o n o f a c e t y le n e -in a c t iv a t e d
c e l l s i n Methylococcus c a p s u la tu s (B ath ) 266
The n e c e s sa ry amount o f methanol f o r the
r e a c t i v a t i o n o f p ro p y le n e -in a c tiv a te d
c e l l s in Methylococcus c a p s u la tu s (B ath ) 267
The n e c e ssa ry amount o f n i t r a t e f o r the
r e a c t i v a t i o n o f a c e t y le n e -in a c t iv a t e d
c e l l s 269
The n e c e s sa ry amount o f n i t r a t e f o r the
r e a c t i v a t i o n o f p r o p y le n e -In a c t iv a t e d
c e l l s 270
The n e c e ssa ry amount o f s u lp h a te f o r the
r e a c t i v a t i o n o f a c e t y le n e -in a c t iv a t e d
c e l l s 272
The n e c e s sa ry amount o f s u lp h a te f o r the
r e a c t i v a t i o n o f p r o p y le n e -in a c tiv a te d
10.6 .1 PO p ro du c tio n under g r o w i n g - c e ll c o n d itio n
u s in g M eth y lo cy stis p arvu s (OBBP) - low
biom ass c o n d itio n 280
1 0 .6 .2 P0 p ro du c tio n under g r o w i n g - c e ll c o n d itio n
u s in g M eth y lo cy stia p arvu s (OBBP) - h igh
biom ass c o n d itio n
281
1 0 .6 .3 The con c e p tu a liz e d p ro c e ss f lo w -s h e e t o f
the g r o w i n g - c e ll process 285
The con c e p tu a liz e d p ro c e s s f lo w -s h e e t o f
the tw o -st a g e r e a c t i v a t i o n p ro c e ss 289
The e f f e c t o f a d d it iv e s on the p reven tion
o f In a c t iv a t io n and the a c c e le r a t io n o f
r e a c t i v a t i o n 301
1 1 .1 .2 The e f f e c t o f sodium c arb o n ate on the P0
p r o d u c t i v it y in v a r io u s m e th a n e -o x id isin g
b a c t e r i a
303
1 1 .1 .3 The e f f e c t o f melamine on the prev en tio n
o f i n a c t i v a t i o n and the a c c e le r a t io n o f
1
.3.1
C la s s i f i c a t i o n o f o b li g a t e methane-o x i d i s i n g b a c t e ria *
1 .11 .1 S u b s tra te s p e c i f i c i t y o f MMO in v iv o
and i n v i t r o 23
1
.17.1
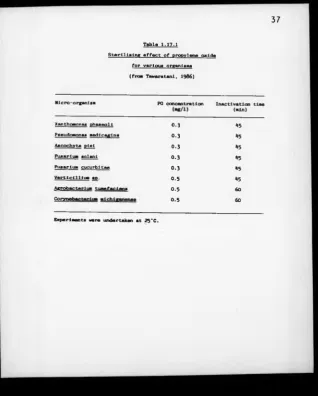
E f f e c t o f gaseous PO on v a r io u s microorganism s 37
2 .2 .1 Com position o f n i t r a t e m in eral s a l t (NMS)
medium <12
3 .1 .1 O x id a t iv e products produced by whole c e l l s
o f Methylococcua c a p su la tu s (B ath ) which
con tain ed soluble-MNO 53
3 .2 .1 Param eters f o r e v a lu a t in g compounds 59
3 .2 .2 G rou ping o f the r e a c tio n s by MMO 60
3 .2 .3 E v a lu a t io n o f p rodu cts by MHO 65
3 .3 .1 Economie comparison o f produ cts 67
3 .3 .2 D ir e c t e p o x idation p ro c e s s e s o f propylene
3.1». 1 E f f e c t o f a lc o h o ls as e le c t r o n donors 71
3-1».2 Whole c e l l a c t i v i t y »e n su re d u s in g
■ethane as the e le c t ro n donor 76
3.5*1 MMO a c t i v i t y in c e l l e x t r a c t s and whole
c e l l MMO a c t i v i t i e s 80
3 .5 *2 Whole c e l l MMO a c t i v i t y o f v a r io u s
■ethan otrophs 82
3 -5 -3 R e la t io n s h ip between b io a a s s concentration
and w h ole c e l l MMO a c t i v i t y
85
4 .3 •1 E f f e c t o f PO con cen tration in the c u lt u re
on c e l l growth
98
4 .5 .1 I n a c t iv a t io n o f whole c e l l s by e x t e r n a lly
added PO in Nethylococcus c a p su la tu s (B ath ) 105
9 *2 .1 The i n h i b i t i o n o f r e a c t i v a t i o n o f a c e ty le n e
-in a c t i v a t e d c e l l s by the a d d it io n o f n i t r i t e
225
10 .3 .1 The t y p i c a l p ro p e rtie s o f the th ree ■ethane-
o x i d i s i n g b a c t e r i a , M ethylococcus capsu latu s
(B a t h ), M eth ylocystls p arvu s (OBBP) and
10 .4 .1
10.6.1
10.6 .2
10 .6 .3
The s t o ic h io a e t r y o f the »e t h a n o l
consumption and P0 p ro du c tio n 26l
The b a s i c a l parameters f o r th e c a lc u la t io n
o f P0 p ro du c tio n c o st i n t w o -st a g e
r e a c t i v a t i o n process
291
The P0 p ro du c tio n c o st in tw o -st a g e
r e a c t i v a t i o n process
293
The c o s t a n a ly s is o f to w e rs, v e s s e ls and
In 1972, Colby and Zataan proposed and d e fin e d the tern
'm e th y lo tro p h y '. A cc o rd in g to t h e ir d e f i n i t i o n , m ethylotrophs a r e
o rg a n is e s c a p ab le o f o b t a i n in g energy by the o x id a tio n o f C j-g ro w th
s u b s tr a te s and the a s s im i la t io n o f carbon such as formaldehyde o r as a
m ixture o f formaldehyde and carbon d io x id e , b u t always by pathways that
a r e d is t in g u is h a b le from the C a lv in c y c le . T h is group o f organism s can
be fu r t h e r s u b -d iv id e d in t o two g r o u p s :- o b li g a t e and fa c u lt a t iv e
m ethylotrophs, the l a t t e r group h aving th e a d d i t i o n a l c a p a b i li t y to grow
and r e p li c a t e on a v a r i e t y o f oth e r c a r b o n -t o -c a r b o n bond compounds.
M ethylotrophs, as d e fin e d above, a r e organism s which can u t i l i z e
methane, methanol, N-m ethyl compounds, S -m eth yl compounds as t h e i r s o le
sources o f carbon f o r grow th and r e p l i c a t i o n . The e x te n siv e and v a rie d
p h y sio lo gy and b io ch em istry o f a l l the d i f f e r e n t types o f methylotrophs
have been com prehensively w ritt e n about i n a number o f re view s by
Quayle, 1972, Colby e t a l . . 1979; H ig g in s e t a l . . 1981a; Anthony, 1982;
Large and Bamforth, 1988. The re a d e r, t h e r e f o r e , should use th ese
sources o f re fe re n c e f o r more d e t a ile d in fo rm a tio n on m ethylotrophy.
1 .2 Occurrence , E c o lo g y and H isto ry o f th e I s o la t i o n o f Methane-
o x id is in g B a c t e r ia
The e x te n t o f tu rn o v e r o f methane w it h in the bio sp h e re as th e major
carbon source o f m e th an e-o x id isin g b a c t e r i a (methanotroph) i s n o t w idely
a p p re c ia te d . In 1976, E h h alt reported ap pro xim ately 50% o f a l l org a n ic
carbon, decomposed by a n a e ro b ic m ic r o flo r a i s converted in to methane.
The methane amounts to 5 - 3 “ 8.1 mega tonnes p e r y e a r.
B io gen ic methane I s the major so u rc e o f atmospheric methane and i s
methane gen erated by methanogenic b a c t e r i a i s due m ainly to the
a c t i v i t i e s o f m e th an e-o x id isin g b a c t e r i a which a r e w id e ly d is t r ib u t e d in
the environment. In c lu d in g the p o s s i b i l i t y th at some methane may be
o x id is e d a n a e r o b ic a lly (Pan gariban e t a l . . 1979; Reeburgh,
1981
) .Although m e th a n e -o x id isin g b a c t e r i a a r e now known to b e w id ely
d is t r ib u t e d in n a t u r e , p r i o r to 1970, o n ly th ree s p e c ie s o f
m eth an e-o xid isin g b a c t e r i a had been I s o la t e d and c h a r a c t e r iz e d (P o s te r
and D a v ie s.
1966
; Brown e t a l . , 1 9 ^ . S t o c k s and N cCleskey, I96
M)d e s p ite the f a c t th a t th e f i r s t i s o l a t e had been found e a r l y in th is
century by Sohngen (1 9 0 6 ). The d i f f i c u l t y o f i s o l a t i n g p u re c u lt u r e s ,
p a r t i c u la r ly o f m e th an e-o x id isin g b a c t e r i a was c e r t a i n l y due to the la c k
o f a r e l i a b l e enrichm ent and i s o la t i o n te ch n iqu e. When en rich ed
c u lt u re s were p la t e d on a g a r p la t e s un der an atmosphere o f methane and
a i r , many scaven gers formed c o lo n ie s w h ich , when r e p la t e d on fr e s h aga r
p la t e s , formed f u r t h e r c o lo n ie s even w it h o u t methane.
The o th e r problem encountered was sy m b io s is . I t was sometimes found
th at m e th a n e -o x id isin g b a c t e r i a grew w e l l i n a n ix e d c u lt u r e (Lin ton and
Buckee, 1977- Imai e t m l . . 1986). The enrichm ent and i s o l a t i o n
technique was d r a m a t ic a lly transform ed i n 1969-1970 by W hittenbury and
h is c o lle a g u e s . They d e v is e d sim ple and e f f e c t i v e tech n iqu es f o r the
is o l a t i o n o f m e th a n e -o x id isin g b a c t e r i a (W h itten bu ry , 1969; Whittenbury
e t a l . , 1970b) from which they is o la t e d more than a hundred d i f f e r e n t
s t r a i n s . The succ ess o f t h e i r techniqu es d e riv e d from th e employment o f
a s h o r t e r enrichment p e r i o d . This li m i t e d lo s s e s o f b a c t e r i a
experienced as a r e s u l t o f p red atio n and overgrow th g row in g on
s u b s tr a te s o th e r than methane.
W hittenbury e t a l . (1970a) c l a s s i f i e d t h e ir i s o l a t e s In t o f i v e
grou ps; M ethylococcus. Methylomonaa. Me th y lo b a c t e r . M eth ylosln u s and
1.3 C la s s i f i c a t i o n o f M eth a n e-oxidisin g B a c t e r ia
Confusion r e l a t i n g to the nomenclature and taxonomy o f
■ e th a n e -o x id is in g b a c t e r i a has not y e t been c o n c lu s iv e ly re so lv e d ,
however the b e s t scheme produced to d a te i s W hittenbury e t a l . (1970a)
who found a c o r r e la t i o n between the type o f aeabrane arrangement, and
the «ean s employed f o r carbon a s s im i la t io n . Type 1 organisms a ssim ilate
carbon by the r i b u l o s e monophosphate pathway, whereas organisms
p o sse ssin g Type I I membrane systems used the s e r i n e pathway.
T a b le 1 .3 -1 shows the c l a s s i f i c a t i o n o f o b li g a t e m ethane-oxidising
b a c t e r i a proposed b y the above au th o rs. However, s in c e the adoption o f
t h is scheme o f c l a s s i f i c a t i o n a number o f r e p o rt s have appeared with
su gg est th a t the two groups are not as c l e a r l y d e fin e d as was f i r s t
th ought. I t has been shown th at some o f Type I and I I methanotrophs can
p ossess both h e x u lo se phosphate synthase and hydroxypyruvate reductase
a c t i v i t y and th is i s in d ic a t iv e o f the presen c e o f the r i b u lo s e
monophosphate c y c le and the s e rin e pathway r e s p e c t iv e ly which has le d to
the p ro p o sa l o f th e t h ir d group o f methanotrophs, the Type X group
(W hittenbury and D a lt o n , 1981).
The c r i t e r i a u se d to re so lv e the problem o f c l a s s i f i c a t i o n o f
■ethanotrophs a r e , a t p resen t, u n s a t is fa c t o r y s in c e the n atu re o f the
organisms makes t h i s ta sk d i f f i c u l t . I n 198**. Galchenko and Andreev
d evised a scheme f o r the c l a s s i f i c a t i o n o f methanotrophs which was based
[image:35.362.12.338.11.402.2]:-C la s s i f i c a t i o n to O b lig a t e M e t h a n e -u t i li s i n g B a c t e ria
C haracter
Membrane arrangement
R estin g s ta g e
Carbon A s sim ila tio n
TCA c y c le
G lucose-6-phosph ate
and 6-ph osphogluconate
dehydrogenase
Examples
Type I
Bundles o f v e s i c u l a r
d is c s
Cysts (A z o t o b a c t e r
-11km)
R ibu lose monophosphate
pathway
Incomplete (la c k s
2-o x 2-o g lu ta ra te
dehydrogenase)
Type I I
P a ire d membranes in
la y e r s around
p eriph e ry
Exospores o r " l i p i d -
c y s t s "
S e rin e pathway
Complete
Hethylococcu» a m a m i
m t t r i — » He thy l o e « t i «
[image:36.363.17.340.6.408.2]by W hittenbury e t a l . 1970b).
I t i s o f paramount im portance t h a t th e problem r e la t i n g to c o rr e c t
c l a s s i f i c a t i o n o f methanotrophs i s th o r o u g h ly r e so lv e d , not ju s t because
i t would g iv e academ ic s a t i s f a c t i o n , b u t because o f the im p lic a t io n s f o r
in d u s t r i e s . For exam ple, in both Europe and Japan, m ic ro b ia l p aten ts are
iss u e d acc ordin g t o genus and/or s p e c ie s th ese can cover a l l the sp e c ie s
which b elo n g to t h a t genus. Even i f new s p e c ie s are found, these
s p e c ie s a r e r e s t r i c t e d by the p aten t which has been a lre a d y is s u e d . But
the p aten t has no r e s t r i c t i o n on any o t h e r genus. The confu sion o f
c l a s s i f i c a t i o n makes i t easy to pro po se a new genus. In t h is case a
p aten t which has be e n is s u e d , has n o t r e s t r i c t i o n f o r the newly proposed
genus. I n the U n it e d S t a t e s , p a te n ts a r e is s u e d acc ordin g to sp e c ie s
o n ly , so the above problem does not a r i s e .
1.4 P h ysiolo g y and Biochem istry o f O b li g a t e M eth a n e-oxidisin g
B a c t e r ia
1 .4 .1 B a s ic Growth Requirements
M e th a n e -o x id isin g b a c t e r i a a r e s t r i c t l y a e r o b ic , due to t h e ir need
f o r gaseous oxygen i n the i n i t i a l o x id a t i o n o f methane (H ig g in s and
Quayle, 1970). They a r e cap able o f u t i l i s i n g e i t h e r methane o r methanol
a s a s o le source o f carbon and en e rg y . The growth o f m eth an e-oxidisin g
b a c t e r i a i s i n h i b i t e d by the a d d it io n o f o rd in a ry h etero tro ph ic
m e ta b o lite s in norm al c o n ce n tration s (E c c le s t o n and K e lly , 1972, 1973)«
No growth fa c t o r s a r e re q u ir e d f o r th e grow th o f these organisms as they
sou rc e, c a lc iu m , Magnesium, p o t a s s iu e , s u lp h a t e , phosphate and trace
elements (D a lt o n and W hittenbury, 1976). Some amino acids stim u late the
growth o f m eth an e-o xid isin g b a c t e r i a (M u r r e ll , 1981).
1 .4 .2 Carbon Metabolism
M e th a n e -o x id isin g b a c t e r i a a r e c a p a b le o f o x id is in g methane to
carbon d io x id e com pletely. Methane i s f i r s t o f a l l o x id is e d to methanol
by way o f the a c t io n o f a methane monooxygenase (MHO). Methanol i s then
fu r t h e r o x id is e d to formaldehyde by a methanol dehydrogenase.
Formaldehyde can be a ssim ila t e d in to the c e l l to fo ra c e l l m a te ria ls o r
i t can be fu r t h e r o x id ise d by way o f a d i s s l a l l a t o r y rou te v i a formate
and f i n a l l y t o carbon d io x id e , to p ro v id e th e c e l l with energy f o r i t s
a s s i a i la t o r y pathway. Both a s s i a i la t o r y and d i s s i a i la t o r y pathways work
sim u ltan eo u sly i n the c e l l .
1 .4 .3 Cl-compound A s sim ila tio n Pathways
Three d e v ic e s f o r the a s s im ila t io n o f Cj-compounds a r e recognized
to d a t e : - the r lb u i o s e monophosphate pathway f o r formaldehyde
a s s im i la t io n , th e s e rin e pathway and the r l b u l o s e diphosphate pathway
(C a lv in c y c le ) f o r carbon d io x id e a s s im i la t io n . Some organisms may use
more than one mechanise f o r C ^ -a s s im i la t io n e i t h e r when su b je c t e d to
d i f f e r e n t grow th c o n d itio n s; o r indeed when two mechanisms a r e used
sim u lta n e o u sly . However th ere i s no c o n v in c in g evidence a s y et o f two
o r more com plete C .- a s s i m ila t i o n pathways o p e ra t in g sim u ltaneously in
any one m ic ro-organism . N e v e rt h e le s s , I t I s c l e a r that c e r t a in
C j - u t i l i z e r s , w h i le usin g one pathway as th e m ajor source o f fix e d
cartoon a r e c a p a b le o f a s s im i la t in g sm all amounts o f C1-compound by a
R ib u lo s e monophosphate pathway (R M P ), a pathway o f formaldehyde
a s s im ila t io n was i n i t i a l l y proposed by Kemp and Quayle (1967) and
fu r t h e r e la b o r a t e d by Lawrence and Q u a yle (1970a) and Kemp (1 9 7 4 ). The
o v e r a ll e f f e c t o f the c y c le i s to s y n t h e s iz e a C^-compound from th ree
m olecules o f formaldehyde. The c y c le i s conven iently d iv id e d in t o three
s ta g e s ; f i x a t i o n , cleavag e and rearrangem ent.
S tage I . F ix a tio n : by th e a c t io n o f hexulose phosphate synthase,
th ree m o le c u le s o f formaldehyde a r e condensed with th re e molecules o f
r ib u lo se -5 -p h o s p h a t e to y i e l d th ree m o le cu les o f fr u e tose-6 -p h osph ate.
S tage 2 , Cleavage: one m olecule o f h exu lose-6-phosph ate i s
isom erised t o fru c tose -6 -ph o sp h a te and t h i s i s then s p l i t in to two C^-
compounds. T h i s bein g ach ieved e i t h e r by the enzymes o f the g l y c o ly t i c
sequence o r b y the Entn er-D oudoroff pathway enzymes.
Stage 3 . Rearrangement: t h is s t a g e in v o lv e s the re g e n e ra tio n o f the
th ree m ole c u le s o f r ib u lo se -5 -p h o s p h a t e from the two m olecules o f
fr u c t o se -6 -p h o sp h a te and one m olecule o f glycera ld eh yd e-3 -p h osph a te
produced in s t a g e s 1 and 2.
1 .4 .5 S e r in e Pathway
The ca rb o n a s s im ila t io n pathway i n the Type I I m eth an e-oxidisin g
b a c t e r i a w ere f i r s t stu died by Q uayle and co-workers (Lawrence e t a l . .
1970«. 1 9 7 0 b). The o v e r a ll r e s u lt o f th e pathway i s t o in co rp orate two
m olecules o f formaldehyde and one m o le c u le o f carbon d io x id e in t o a
C^-compound. 3~phosphoglycerate. The c y c le i s i n i t i a t e d by two
m olecules o f s e r i n e which a r e f o rmed from two m olecules o f g ly c in e plu s
in to 3 -ph osp h oglyc era te f o r a s s i m i la t i o n in to c e l l carbon o r converted
in to phosphoenol pyruvate (P E P ). PEP c arbo xylase c a t a ly s e s the
c a r b o x y la t io n o f PEP w ith carbon d io x id e to fo r a o x a lo a c e t a t e , which
su b seq u e n tly forms m alyl-CoA. The a a ly l- C o A i s then c leav ed in t o two
C2 u n it s t o a c t as fu r t h e r a c c e p t o rs o f C1 u n its to m aintain the
c y c le o f r e a c t i o n s .
1 .4 .6 R ib u lo s e Diphosphate Pathway o f Carbon D iox id e A s sim ila tio n
M ethylococcus cap su la tu s ( B a t h ) , a Type I methanotroph, appeared to
use on ly th e r i b u lo s e monophosphate pathway f o r a s s im i la t io n (S tr^ a
e t a l . . 197*0 • However, i t h as now been shown to possess
hydroxypyru vate redu ctase, a key enzyme in the s e r i n e pathway, although
a t low l e v e l (Reed, 1976). Furtherm ore the presence o f the key enzymes
o f the C a lv i n c y c le , r i b u lo s e dip h osp h ate c a rb o x y la se and
p h o sp h o ribu lo k in ase were dem onstrated i n c e l l e x t r a c t s o f Hethylococcus
c a p su la tu s (B ath ) (T a y lo r , 1977)* The r a t e o f C02 f i x a t i o n by whole
c e l l was lo w , and c o n trib u te d o n ly a bo u t 2.5% (w t/w t) o f the t o t a l
c e l l c a rb o n . F ix a tio n was o b se rv e d o n ly w ith the presence o f methane,
in d i c a t i n g an energy requirem ent f o r in c o rp o r a tio n (T a y lo r e t a l . ,
1980).
R ib u lo s e diphosphate c a r b o x y la s e from t h is organism r e q u ir e s a
d iv a le n t c a t io n f o r a c t i v i t y . I t h as an a lk a lin e pH optimum; i s
in h ib i t e d b y 6-phosphogluconate and p o ssesses an oxygenase a c t i v i t y
(T a y lo r e t a l . . 1980). T h is s eco n d ary a c t i v i t y o f th e enzyme gen erates
phosphoglucon ate, which may b e s u b se q u e n tly c le a r e d by a s p e c i f i c
phosphogluconate phosphatase. T h is was observed i n e x t r a c t s o f
M ethylococcus capsu latu s (B a t h ) (T a y lo r e t a l . , 19 81 ). The m etabolic