0095-1137/05/$08.00⫹0 doi:10.1128/JCM.43.7.3314–3319.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Uncultivated Phylotypes and Newly Named Species Associated with
Primary and Persistent Endodontic Infections
J. F. Siqueira, Jr.* and I. N. Ro
ˆc¸as
Department of Endodontics, Esta´cio de Sa´ University, Rio de Janeiro RJ, Brazil
Received 21 January 2005/Returned for modification 12 March 2005/Accepted 16 March 2005
Endodontic infections have been traditionally studied by culture methods, but recent reports showing that over 50% of the oral microbiota is still uncultivable (B. J. Paster et al., J. Bacteriol. 183:3770–3783, 2001) raise the possibility that many endodontic pathogens remain unknown. This study intended to investigate the prevalence of several uncultivated oral phylotypes, as well as newly named species in primary or persistent endodontic infections associated with chronic periradicular diseases. Samples were taken from the root canals of 21 untreated teeth and 22 root-filled teeth, all of them with radiographic evidence of periradicular bone destruction. Genomic DNA was isolated directly from each sample, and 16S rRNA gene-based nested or heminested PCR assays were used to determine the presence of 13 species or phylotypes of bacteria. Species-specific primers had already been validated in the literature or were developed by aligning closely related 16S rRNA gene sequences. Species specificity for each primer pair was confirmed by running PCRs against a panel of several oral bacteria and by sequencing DNA from representative positive samples. All species or phylotypes were detected in at least one case of primary infections. The most prevalent species or phylotypes found in primary infections wereDialister invisus(81%),Synergistesoral clone BA121 (33%), andOlsenella uli(33%). Of the target bacteria, only these three species were detected in persistent infections. Detection of uncultivated phylotypes and newly named species in infected root canals suggests that there are previously unrecognized bacteria that may play a role in the pathogenesis of periradicular diseases.
Periradicular diseases are arguably among the most common inflammatory diseases that affect humans (10). Overwhelming evidence indicates that microorganisms infecting the root canal of the teeth are the major causative agents of these diseases (27). Data from culture and molecular studies have demon-strated that the microbiota associated with primary endodontic infections is conspicuously dominated by anaerobic bacteria and that an infected root canal can harbor from 10 to 30 bacterial species (22, 33, 34, 37). On the other hand, the mi-crobiota of persistent endodontic infections associated with treatment failure has been shown to be composed of fewer species, with dominance of facultative bacteria (32, 35). Even though over 300 bacterial species have already been isolated from or detected in infected root canals, no single species has been consistently found to be the major endodontic pathogen. Application of molecular genetic methods to the analysis of the bacterial diversity in the oral cavity has revealed a still broader spectrum of extant bacteria than previously reported by cultivation approaches (23). Overall, over 700 different spe-cies belonging to 11 divisions (or phyla) of the domainBacteria have been detected in the oral cavity of humans (17, 23, 24). About 50% of these bacteria are known only by 16S rRNA gene sequences (phylotypes) (23). This raises the interesting possibility that uncultivated and as-yet-uncharacterized species that have remained undetected in studies by traditional iden-tification methods may make up a large fraction of the living oral microbiota and may participate in the etiology of oral diseases, including periradicular diseases.
PCR amplification of conserved regions of the 16S rRNA gene, followed by cloning and sequencing of PCR products, has been widely applied to study of the bacterial diversity in different human healthy and diseased sites. However, the clon-ing approach is usually time consumclon-ing, labor intensive, and expensive, being virtually impractical for analysis of multiple samples in epidemiological investigations. Several studies have devised PCR primers based on sequences from oral clone libraries to survey larger numbers of samples from periodon-tally diseased subjects for the presence of uncultivated bacteria in an attempt to establish correlations with disease etiology (15, 19, 20, 26). Most of these studies revealed that several phylotypes may play an important role in periodontal disease. To the best of our knowledge, no study has as yet used such approaches to investigate the prevalence of uncultivated oral phylotypes in endodontic infections.
The applicability of molecular genetic methods does not rely uniquely on the detection of uncultivable bacteria but also on a more reliable identification of diverse bacterial species, par-ticularly some nutrient-demanding and therefore difficult-to-grow oral anaerobic bacteria. Several fastidious bacterial spe-cies have only been recently reported to occur in endodontic infections by molecular methods. These includeTannerella for-sythia(6, 34), severalTreponemaspecies (16, 25, 29),Prevotella tannerae(39),Filifactor alocis(28),Dialister pneumosintes(30), Haemophilus aphrophilus(34),Eubacterium infirmum(12), and Centipeda periodontii(31), all of them recognized periodontal pathogens. Members of the familyCoriobacteriaceae, particu-larly from the genera Olsenella and Atopobium, have been commonly detected in association with some oral diseases, including caries and marginal periodontitis (21, 23), but they have never been consistently found in endodontic infections,
* Corresponding author. Mailing address: Department of Endodon-tics, Esta´cio de Sa´ University, Av. Almte Ary Parreiras 311/1001 Icaraı´, Nitero´i, RJ Brazil 24230-322. Phone: 55 21 8874-1022. Fax: 55 21 2503-7289, ext. 223. E-mail: [email protected].
3314
on May 15, 2020 by guest
http://jcm.asm.org/
and difficulties in identifying these species by phenotype-based methods can be one of the reasons for that.
It has been revealed that the main putative endodontic pathogens are also the main periodontal pathogens, with few (if any) exceptions (27, 37). Several new species or phylotypes have been suggested to be involved with the pathogenesis of periodontal diseases (19, 23), but it is still uncertain whether most of these phylotypes are present in infected root canals associated with periradicular diseases. Therefore, the present investigation intended to use a devised nested or heminested PCR assay to survey samples from primary or persistent end-odontic infections for the presence of some newly named bac-terial species and uncultivated phylotypes that have been re-cently detected in association with periodontal diseases. Most of the target bacteria had never been previously found in endodontic infections. For the species already detected in end-odontic infections, prevalence had not been previously inferred because of the small number of samples examined (11, 22).
MATERIALS AND METHODS
Subjects.Root canal samples collected for previous investigations (25, 29, 32) and stored in TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8) at⫺20°C were available for reanalysis in this study. Samples were taken from patients who had been seeking root canal treatment or retreatment at the Department of End-odontics, Esta´cio de Sa´ University, Rio de Janeiro RJ, Brazil. Only teeth from adult patients (ages ranging from 18 to 80 years), all of them having radiographic evidences of periradicular disease, were included in this study. Overall, 43 root canal samples were obtained and grouped as follows, according to the clinical diagnoses: (i) 21 cases of primary endodontic infections (untreated teeth) asso-ciated with asymptomatic chronic periradicular lesions and (ii) 22 cases of per-sistent endodontic infections (root-filled teeth) associated with asymptomatic chronic periradicular lesions, which had been selected for retreatment.
All the teeth with primary endodontic infections showed carious lesions and necrotic pulps. All the teeth with persistent infections had endodontic therapy completed⬎2 years earlier. Root-filled teeth were coronally restored and no direct exposure of the filling material to the oral cavity was evident. Termini of the root canal fillings ranged from 0 to 4 mm short of the radiographic apex. All selected teeth showed no significant gingival recession and an absence of peri-odontal pockets of⬎4 mm deep.
Sampling procedures and DNA extraction.Root canal samples were taken from untreated or root-filled teeth under strict aseptic conditions as previously described (25, 29, 32). Endodontic files with the handle cut off and paper points used for sampling the canals were transferred to cryotubes containing 1 ml of 5% dimethyl sulfoxide in trypticase-soy broth (Difco, Detroit, MI) and immediately frozen at⫺20°C. Further, samples were brought to room temperature, and DNA was extracted using the protocol described previously (25, 29, 32).
The following strains were used in the present study:Actinobacillus actinomy-cetemcomitansATCC 43718,Actinomyces radicidentisCCUG 42377,Atopobium parvulumATCC 33793,Atopobium rimaeATCC 49626,Enterococcus faecalis
ATCC 29212,F. alocisATCC 35896,Fusobacterium nucleatumsubsp.nucleatum
ATCC 25586, Fusobacterium nucleatum subsp. polymorphum ATCC 10953,
Olsenella uliATCC 49627,Olsenella profusaDSM 13989,Porphyromonas end-odontalisATCC 35406,Porphyromonas gingivalisATCC 33277,Prevotella inter-mediaATCC 25611,Prevotella pallensATCC 700821,Propionibacterium propi-onicumU13a-a,Pseudoramibacter alactolyticusC11b-d,Streptococcus intermedius
ATCC 27335,T. forsythiaATCC 43037,Treponema denticolaB1 (Forsyth Dental Institute), andTreponema lecithinolyticum ATCC 700332. All bacteria were grown under appropriate culture conditions. Identification and purity of the cultures were confirmed by Gram staining, microscopic morphotyping, visualiza-tion of colony features on agar plates under magnificavisualiza-tion, growth on selective medium, and biochemical tests. Bacterial DNA was prepared as described pre-viously (25, 29, 32).
Design of specific PCR primers.16S rRNA gene species- or phylotype-specific PCR primers were used. Primer sequences were as previously described or slightly modified therefrom (4, 19), except for those specific forSynergistesoral clone BA121 (formerlyFlexistipesBA121),Synergistesoral clone E3_33 (formerly
FlexistipesE3_33),O. uli,O. profusa, andP. pallens, which were designed for this study. Briefly, 16S rRNA gene sequences of each of these bacteria were retrieved
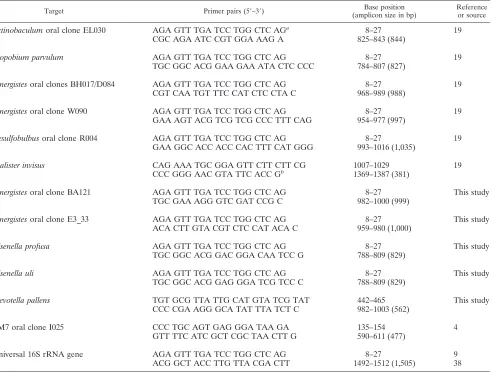
from the GenBank at the National Center for Biotechnology Information web-site and aligned with the sequences of their nearest neighbors in the phylogenetic tree using the CLUSTAL W program (36) to identify variable areas between species. Potential primers were designed from these areas and BLAST (1) was used to verify their specificity by comparing primer sequences with all available sequences in the GenBank database. Based on BLAST searches, one or two primer sequences for each of the target species/phylotypes were selected and further tested for specificity against DNA from the bank of reference strains used as controls and by sequencing of PCR products obtained from representative positive clinical samples. The PCR species- or phylotype-specific primers are shown in Table 1.
Nested or heminested PCR identification.The whole-genome DNA extracts from clinical samples were used as templates in a 16S rRNA gene-based nested or heminested PCR method devised to detect the target species/phylotypes in endodontic samples. In the first PCR, a practically full-length 16S rRNA gene fragment was amplified using a pair of universal 16S rRNA gene primers, which consisted of the forward universal primer 27f (9) and the reverse universal primer 1,492r (38). Aliquots of 5l (each) of the supernatant from clinical samples were used as targets in the first PCR. PCR amplification was performed with 25l of reaction mixture containing a 0.2M concentration of forward and reverse universal primers, 2.5l of 10⫻PCR buffer (Biotools, Madrid, Spain), 2 mM MgCl2, 1.25 U of Tth DNA polymerase (Biotools), and 25M each deoxyribo-nucleoside triphosphate (Biotools).
Afterwards, 1l of the universal reaction mixture was then used as a template for the nested or heminested specific reaction. The second PCR used to assess the occurrence of the target species/phylotypes was performed with 50l of reaction mixture containing 1M concentration of each primer, 5l of 10⫻ PCR buffer (Biotools), 2 mM MgCl2, 1.25 U of Tth DNA polymerase (Biotools), and 0.2 mM each deoxyribonucleoside triphosphate (Biotools). PCRs were per-formed in 25-well microtiter plates. Negative controls consisting of sterile ultra-pure water instead of sample were included with each batch of samples analyzed. Preparations were amplified in a DNA thermocycler (Mastercycler personal; Eppendorff, Hamburg, Germany). The PCR temperature profile for the univer-sal reaction included an initial denaturation step at 97°C for 1 min; followed by 26 cycles, each of a denaturation step at 97°C for 45 s, a primer-annealing step at 55°C for 45 s, and an extension step at 72°C for 1 min; and a final step of 72°C for 4 min. PCR cycling conditions for the second round of amplification specific forD. invisuscomprised an initial denaturation step of 95°C for 10 min; followed by 26 cycles, each of denaturation at 95°C for 30s, primer annealing at 68°C for 1 min, and an extension at 72°C for 1 min; and a final extension step of 72°C for 2 min. For TM7 oral clone I025, cycling conditions consisted of an initial dena-turation step of 95°C for 10 min; followed by 28 cycles, each of denadena-turation at 95°C for 30s, primer annealing at 60°C for 1 min, and an extension at 72°C for 1 min; and then a final extension step of 72°C for 3 min. ForOlsenellaspecies and
Synergistesoral clone BA121, the temperature profile included an initial dena-turation step at 95°C for 2 min, and a touchdown PCR was performed as follows: the denaturing temperature of each cycle was carried out at 95°C for 30 s. The annealing temperature was initially set at 68°C and was then lowered 0.5°C every other cycle until it reached 63°C. Seventeen additional cycles were carried out at 63°C. Primer annealing was performed using this scheme for 30 s, and primer extension was carried out at 72°C for 1 min. The final extension step was at 72°C for 5 min. The temperature profile for the other species/phylotypes encompassed an initial denaturation step at 95°C for 2 min, and touchdown PCR as follows: a denaturing temperature of each cycle at 95°C for 30 s amd an annealing tem-perature initially set at 64°C and then lowered 0.5°C every other cycle until it reached 61°C. Twenty-one additional cycles were carried out at 61°C. Primer annealing was performed using this scheme for 30 s, and primer extension was carried out at 72°C for 1 min. The final extension step was at 72°C for 5 min. PCR amplicons were separated by electrophoresis in a 1.5% agarose gel, which was stained with 0.5g/ml ethidium bromide and viewed under UV transillumina-tion. A 100-bp DNA ladder digest (Biotools) served as the molecular size stan-dard.
Sequencing. To confirm the specificity of the primers, randomly selected representative PCR products for each target species or phylotypes were purified with a PCR purification system (Wizard PCR Preps; Promega, Madison, WI) and then sequenced directly on the ABI 377 automated DNA sequencer using dye terminator chemistry (Amersham Biosciences, Little Chalfont, Buckingham-shire, United Kingdom). Sequence data and chromatograms were inspected and edited by using BioEdit software (http://www.mbio.ncsu.edu/BioEdit/bioedit .html) (14). Sequences were then analyzed using the BLAST algorithm (1).
on May 15, 2020 by guest
http://jcm.asm.org/
RESULTS
After the first round of amplification using universal bacte-rial primers, all samples yielded an amplicon of approximately 1,500 bp. This indicated that bacteria were present in all cases examined, demonstrating the suitability of the DNA for PCR analysis and indicating the absence of inhibitors in the reaction mixture. Negative controls using sterile ultrapure water instead of sample yielded no amplicon.
The sequences of primers specific for each species or phy-lotype are shown in Table 1. The specificity of each primer was tested against a panel of representative oral bacteria. The use of each primer set resulted in no PCR product of the expected size from nontargeted species. Primers specific forO. uli, O. profusa,A. parvulum, andP. pallensresulted in one band of the expected size when respective reference DNA was used. To confirm the specificity of each species- or phylotype-specific PCR primer, representative PCR products of the expected size obtained from clinical samples were sequenced and compared to the original sequences in GenBank database. The primer sequence forD. invisuswas originally designed to target
[image:3.585.51.543.81.454.2]Diali-steroral clone GBA27 (19) but it also annealed without any mismatches to the sequences ofD. invisus and Dialisteroral clones FY011 and BS095. However, sequences of the eight randomly selected representative PCR products from clinical samples all showed higher levels of similarity toD. invisus(99.4 to 100%). Nonetheless, even though our discussion hereafter relies on this species, the possibility exists that some of the nonsequenced PCR products could have been from other Di-alisterphylotypes. ForO. uli, the levels of similarity among the sequences were 98.7 to 100%. Two amplicons generated by the O. uliprimer yielded sequences of a newOlsenellaphylotype, EI15. For O. profusa, the levels of similarity among the se-quences were 99.1 to 100%. ForP. pallens, the levels of simi-larity among the sequences were 99.8 to 100%. ForA. parvu-lum, the level of similarity between the sequences was 99.3%. For Synergistes clones BH017/D084 (formerly Deferribacteres BH017/D084), the levels of similarity among the sequences were 98.6 to 100%. All sequences showed higher similarities to clone BH017. For Synergistes BA121, the levels of similarity among the sequences were 99.3 to 100%. For Synergistes
TABLE 1. PCR primer pairs used for detection of novel species/phylotypes in endodontic infections
Target Primer pairs (5⬘–3⬘) Base position
(amplicon size in bp)
Reference or source
Actinobaculumoral clone EL030 AGA GTT TGA TCC TGG CTC AGa 8–27 19
CGC AGA ATC CGT GGA AAG A 825–843 (844)
Atopobium parvulum AGA GTT TGA TCC TGG CTC AG 8–27 19
TGC GGC ACG GAA GAA ATA CTC CCC 784–807 (827)
Synergistesoral clones BH017/D084 AGA GTT TGA TCC TGG CTC AG 8–27 19
CGT CAA TGT TTC CAT CTC CTA C 968–989 (988)
Synergistesoral clone W090 AGA GTT TGA TCC TGG CTC AG 8–27 19
GAA AGT ACG TCG TCG CCC TTT CAG 954–977 (997)
Desulfobulbusoral clone R004 AGA GTT TGA TCC TGG CTC AG 8–27 19
GAA GGC ACC ACC CAC TTT CAT GGG 993–1016 (1,035)
Dialister invisus CAG AAA TGC GGA GTT CTT CTT CG 1007–1029 19
CCC GGG AAC GTA TTC ACC Gb 1369–1387 (381)
Synergistesoral clone BA121 AGA GTT TGA TCC TGG CTC AG 8–27 This study
TGC GAA AGG GTC GAT CCG C 982–1000 (999)
Synergistesoral clone E3_33 AGA GTT TGA TCC TGG CTC AG 8–27 This study
ACA CTT GTA CGT CTC CAT ACA C 959–980 (1,000)
Olsenella profusa AGA GTT TGA TCC TGG CTC AG 8–27 This study
TGC GGC ACG GAC GGA CAA TCC G 788–809 (829)
Olsenella uli AGA GTT TGA TCC TGG CTC AG 8–27 This study
TGC GGC ACG GAG GGA TCG TCC C 788–809 (829)
Prevotella pallens TGT GCG TTA TTG CAT GTA TCG TAT 442–465 This study
CCC CGA AGG GCA TAT TTA TCT C 982–1003 (562)
TM7 oral clone I025 CCC TGC AGT GAG GGA TAA GA 135–154 4
GTT TTC ATC GCT CGC TAA CTT G 590–611 (477)
Universal 16S rRNA gene AGA GTT TGA TCC TGG CTC AG 8–27 9
ACG GCT ACC TTG TTA CGA CTT 1492–1512 (1,505) 38
aUniversal 16S rRNA gene forward primer (base position relative toE. coli16S rRNA gene). bUniversal 16S rRNA gene reverse primer (base position relative toE. coli16S rRNA gene).
on May 15, 2020 by guest
http://jcm.asm.org/
E3_33, the level of similarity between the sequences was 99.7%. For Synergistes clone W090 (formerly Deferribacteres W090), the levels of similarity among the sequences were 98.4 to 99.2%. For Desulfobulbus R004, the levels of similarity among the sequences of the expected size were 98 to 100%. A smaller PCR product (about 175-bp long) was also found in many specimens amplified with theDesulfobulbusR004 primer and showed 100% similarity to theD. invisus sequence. For ActinobaculumEL030, the levels of similarity among the se-quences were 98 to 99.1%. For TM7 oral clone I025, the levels of similarity among the sequences were 98.7 to 100%.
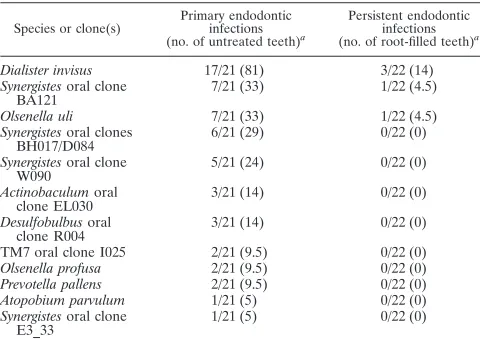
All target species/phylotypes were found in at least one sample from primary endodontic infections (untreated teeth). Only the three most prevalent species in primary infections were detected in persistent endodontic infections (root-filled teeth), but at lower frequencies. Prevalence data for the target species/phylotypes in primary or persistent endodontic infec-tions are represented in Table 2.
DISCUSSION
In this study, we investigated the occurrence of some named species and uncultivated phylotypes in endodontic infections. Named species comprised newly proposed species or species that have undergone recent taxonomic reclassification (7, 8, 18). The target species are usually difficult to cultivate in arti-ficial media and/or difficult to identify by phenotypic features (7, 8, 18), and this may have resulted in underestimation of their presence in endodontic infections. For instance,Dialister species have been consistently detected in endodontic infec-tions only after the advent of molecular genetic approaches (22, 30).Dialister pneumosinteshas been found at high preva-lence in root canals from teeth associated with different forms of periradicular diseases (30). The newly namedD. invisusis an anaerobic gram-negative coccobacillus closely related to Diali-ster pneumosintes, with 93% sequence similarity between the two taxa (8). Munson et al. (22) first isolated this species from infected root canals in a study using cultivation and 16S rRNA
gene sequencing. This was the only species detected in all five cases analyzed in their study (22). In the present investigation, D. invisuswas detected in 81% of the primarily infected root canals. This considerably high prevalence lends additional cre-dence to the assumption that this species can be an important endodontic pathogen. Although in lower prevalence values,D. invisuswas also detected in cases of failed endodontic treat-ment (14% of the teeth). This suggests that this species can also be involved with persistent endodontic infections.
Although Olsenella and Atopobium species have been re-cently isolated/detected in endodontic infections (5, 11, 22), no study had consistently investigated their prevalence in these infections. Cultural and molecular analyses of samples taken from five infected root canals revealed the occurrence ofO. uli in three cases,A. parvulumin two cases, andO. profusain one case (22). O. ulihas been found to predominate over other gram-positive rods in root canal samples taken after chemo-mechanical preparation and intracanal medication, suggesting that this species can resist intracanal disinfection measures and thereby may be involved in persistent infections (5). Herein,O. uliwas present in one-third of the samples from primary in-fections and in one root-filled tooth.O. profusaandA. parvu-lum were detected in 9.5% and 5% of the untreated teeth, respectively, but in no retreatment case. These findings re-vealed thatOlsenellaspecies, particularlyO. uli, are common members of the microbiota associated with primary endodon-tic infections.
Prevotella species have been frequently found in different types of endodontic infections (2, 13, 34). The most commonly isolated/detected species include Prevotella intermedia, Pre-votella nigrescens, andP. tannerae(2, 39), but others, including nonpigmented species, have also been found (13). In this study, we devised a pair of PCR primers to identify P. pallens, a black-pigmentedPrevotellaspecies, directly in clinical samples and detected this species in 9.5% of the primarily infected canals. This is the first report of the occurrence ofP. pallensin endodontic infections.
[image:4.585.43.283.98.270.2]In addition to newly named species, this study also focused the occurrence of as-yet-uncultivated bacteria in endodontic infections. All phylotypes targeted in this study were detected in at least one sample from primary endodontic infections; one phylotype was found in one case of persistent infection. Clones from theSynergistesdivision were found to be common mem-bers of the endodontic microbiota in untreated teeth. One-third of these samples harboredSynergistesoral clone BA121, which was first detected in one subject with refractory peri-odontitis (23). This phylotype was also detected in one root-filled tooth. Although firstly detected in endodontic samples (22),Synergistesclone E3_33 was detected in only one speci-men. Clone BH017 (or D084) from theSynergistesgroup was also detected at a relatively high prevalence value, i.e., 29%, of the primary infections. Although clones BH017 and D084 were indistinguishable by our assay, all sequences from representa-tive amplicons yielded higher similarities to the former. These two clones have also been among the most strongly associated with marginal periodontitis (19).Synergistes clone W090 was detected in 24% of the primarily infected root canals. This clone has been reported to occur in 73% of samples taken from subjects with periodontitis (19). Detection of these unculti-vatedSynergistesphylotypes at high prevalence in infected root
TABLE 2. Frequency of detection of novel species/phylotypes in primary or persistent endodontic infections as revealed by 16S
rRNA gene-based nested or heminested PCR
Species or clone(s)
Primary endodontic infections (no. of untreated teeth)a
Persistent endodontic infections (no. of root-filled teeth)a
Dialister invisus 17/21 (81) 3/22 (14)
Synergistesoral clone BA121
7/21 (33) 1/22 (4.5)
Olsenella uli 7/21 (33) 1/22 (4.5)
Synergistesoral clones
BH017/D084 6/21 (29) 0/22 (0)
Synergistesoral clone W090
5/21 (24) 0/22 (0)
Actinobaculumoral
clone EL030 3/21 (14) 0/22 (0)
Desulfobulbusoral clone R004
3/21 (14) 0/22 (0)
TM7 oral clone I025 2/21 (9.5) 0/22 (0)
Olsenella profusa 2/21 (9.5) 0/22 (0)
Prevotella pallens 2/21 (9.5) 0/22 (0)
Atopobium parvulum 1/21 (5) 0/22 (0)
Synergistesoral clone
E3_33 1/21 (5) 0/22 (0)
aNumber of positive cases for the target species/number of samples examined
(percentage).
on May 15, 2020 by guest
http://jcm.asm.org/
canals suggests that they can be previously unrecognized bac-teria that play a role in the pathogenesis of periradicular le-sions.
Other uncultivated phylotypes targeted in this study were also detected for the first time in primary endodontic infec-tions.Desulfobulbusoral clone R004 andActinobaculumoral clone EL030 were both detected in 14% of the samples. These phylotypes have been previously detected at high frequencies in subjects with marginal periodontitis (82% and 44%, respec-tively) (19). TM7 oral clone I025 was detected in 9.5% of the cases. This finding expands the list of phyla represented in endodontic infections to include TM7 bacteria. Candidate di-vision TM7 is one of the several didi-visions described for the domainBacteria, and because it has no cultivated representa-tive, members can be identified only by molecular methods. Recent studies have demonstrated a high prevalence of TM7 bacteria, particularly clone I025, in association with periodon-tal diseases (4, 19). Based on our findings, it appears that clone I025 can be found in infected root canals, but it is not so prevalent in endodontic infections as it is in periodontal infec-tions. Such a difference may be a result of different environ-mental conditions between the root canal and the gingival crevice, the latter being more favorable to the establishment of TM7 bacteria. However, differences may have been due to methodological reasons or geographical influence (3, 4).
As expected, samples from root-filled teeth yielded few pos-itive results. Because of the restricted room for bacterial es-tablishment and the low availability of nutrients within treated root canals, the number of bacterial species composing the microbiota associated with persistent endodontic infections is significantly lower than that occurring in primary endodontic infections (32, 35). Thus, only the bacterial species that are able to adapt to the bleak environmental conditions within filled root canals will succeed in causing persistent endodontic infections.E. faecalishas been shown to be the most frequently detected species in root-filled teeth, and 17 of the 22 cases examined herein had been positive for this facultative species (32). Even so, this is the first study to reveal the occurrence of D. invisus,SynergistesBA121, andO. uliin the canals of root-filled teeth; their possible association with recalcitrant perira-dicular diseases merits further elucidation.
The microbial etiology of periradicular diseases has been demonstrated to be more complex than previously anticipated by cultivation studies. In addition to detecting some cultivable species in increased prevalence, molecular methods have also expanded the list of putative endodontic pathogens by inclu-sion of some fastidious bacterial species or even uncultivated bacteria that had never been previously found in endodontic infections by cultivation procedures (27). As a consequence, a larger and ever-expanding number of species have been sus-pected to be involved with causation of periradicular diseases. Since endodontic infections develop in a previously sterile place which as such does not contain a normal microbiota, every bacterial species present in the mixed consortium has the potential to play a role in the infectious process. Although the PCR assay used in the present study does not allow quantita-tion, identification of the taxa present in endodontic samples and determination of their prevalence are still of extreme value to help unravel the bacterial diversity in infected root canals. This information will make it possible to use other techniques
such as real-time PCR, DNA microarrays, or reverse-capture DNA-DNA hybridization assays to investigate the numbers of these taxa in root canal samples.
In conclusion, the present investigation has expanded the list of endodontic bacteria by including several newly named oral species, as well as novel phylotypes recently identified by 16S rRNA gene sequence analysis. These findings point to an ur-gent need for a steady and sustained effort to obtain these bacteria in culture so that their phenotypic traits, including virulence and susceptibility to antimicrobials, can be properly assessed.
ACKNOWLEDGMENTS
This study was supported by grants from CNPq, a Brazilian Gov-ernmental Institution.
We express our gratitude to Jari Jalava, Bruce Paster, Go¨ran Sundqvist, Lu´cia Teixeira, and Arie van Winkelhoff for providing some of the bacterial strains used in this study.
REFERENCES
1.Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman.1990. Basic local alignment search tool. J. Mol. Biol.215:403–410.
2.Baumgartner, J. C., B. J. Watkins, K. S. Bae, and T. Xia.1999. Association of black-pigmented bacteria with endodontic infections. J. Endod.25:413– 415.
3.Baumgartner, J. C., J. F. Siqueira, Jr., T. Xia, and I. N. Roˆc¸as. 2004. Geographical differences in bacteria detected in endodontic infections using polymerase chain reaction. J. Endod.30:141–144.
4.Brinig, M. M., P. W. Lepp, C. C. Ouverney, G. C. Armitage, and D. A. Relman.2003. Prevalence of bacteria of division TM7 in human subgingival plaque and their association with disease. Appl. Environ. Microbiol. 69:
1687–1694.
5.Chavez de Paz, L. E., A. Molander, and G. Dahlen.2004. Gram-positive rods prevailing in teeth with apical periodontitis undergoing root canal treatment. Int. Endod. J.37:579–587.
6.Conrads, G., S. E. Gharbia, K. Gulabivala, F. Lampert, and H. N. Shah.
1997. The use of a 16S rDNA PCR for the detection of endodontopatho-genic bacteria. J. Endod.23:433–438.
7.Dewhirst, F. E., B. J. Paster, N. Tzellas, B. Coleman, J. Downes, D. A. Spratt, and W. G. Wade.2001. Characterization of novel human oral isolates and cloned 16S rDNA sequences that fall in the familyCoriobacteriaceae: de-scription of Olsenella gen. nov., reclassification of Lactobacillus uli as
Olsenella ulicomb. nov. and description ofOlsenella profusasp. nov. Int. J. Syst. Evol. Microbiol.51:1797–1804.
8.Downes, J., M. Munson, and W. G. Wade.2003.Dialister invisussp. nov., isolated from the human oral cavity. Int. J. Syst. Evol. Microbiol.53:1937– 1940.
9.Edwards, U., T. Rogall, H. Blo¨cker, M. Emde, and E. C. Bo¨tter.1989. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res.17:7843–7853.
10.Figdor, D.2002. Microbial aetiology of endodontic treatment failure and pathogenic properties of selected species. Umea University odontological dissertations no. 79. Umea University, Umea, Sweden.
11.Fouad, A. F., J. Barry, M. Caimano, M. Clawson, Q. Zhu, R. Carver, K. Hazlett, and J. D. Radolf.2002. PCR-based identification of bacteria asso-ciated with endodontic infections. J. Clin. Microbiol.40:3223–3231. 12.Fouad, A. F., K.-Y. Kum, M. L. Clawson, J. Barry, C. Abenoja, Q. Zhu, M.
Caimano, and J. D. Radolf.2003. Molecular characterization of the presence ofEubacteriumspp. andStreptococcusspp. in endodontic infections. Oral Microbiol. Immunol.18:249–255.
13.Haapasalo, M.1989.Bacteroidesspp. in dental root canal infections. Endod. Dent. Traumatol.5:1–10.
14.Hall, T. A.1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser.41:95–98.
15.Harper-Owen, R., D. Dymock, V. Booth, A. J. Weightman, and W. G. Wade.
1999. Detection of unculturable bacteria in periodontal health and disease by PCR. J. Clin. Microbiol.37:1469–1473.
16.Jung, I.-Y., B.-K. Choi, K.-Y. Kum, Y.-J. Yoo, T.-C. Yoon, S.-J. Lee, and C.-Y. Lee.2001. Identification of oral spirochetes at the species level and their association with other bacteria in endodontic infections. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod.92:329–334.
17.Kazor, C. E., P. M. Mitchell, A. M. Lee, L. N. Stokes, W. J. Loesche, F. E. Dewhirst, and B. J. Paster.2003. Diversity of bacterial populations on the
on May 15, 2020 by guest
http://jcm.asm.org/
tongue dorsa of patients with halitosis and healthy patients. J. Clin. Micro-biol.41:558–563.
18.Ko¨no¨nen, E., E. Eerola, E. V. G. Frandsen, J. Jalava, J. Ma¨tto¨, S. Salmen-linna, and H. Jousimies-Somer. 1998. Phylogenetic characterization and proposal of a new pigmented species to the genus Prevotella:Prevotella pallenssp. nov. Int. J. Syst. Bacteriol.48:47–51.
19.Kumar, P. S., A. L. Griffen, J. A. Barton, B. J. Paster, M. L. Moeschberger, and E. J. Leys.2003. New bacterial species associated with chronic peri-odontitis. J. Dent. Res.82:338–344.
20.Leys, E. J., S. R. Lyons, M. L. Moeschberger, R. W. Rumpf, and A. L. Griffen.
2002. Association ofBacteroides forsythusand a novelBacteroidesphylotype with periodontitis. J. Clin. Microbiol.40:821–825.
21.Munson, M. A., A. Banerjee, T. F. Watson, and W. G. Wade.2004. Molecular analysis of the microflora associated with dental caries. J. Clin. Microbiol.
42:3023–3029.
22.Munson, M. A., T. Pitt-Ford, B. Chong, A. Weightman, and W. G. Wade.
2002. Molecular and cultural analysis of the microflora associated with end-odontic infections. J. Dent. Res.81:761–766.
23.Paster, B. J., S. K. Boches, J. L. Galvin, R. E. Ericson, C. N. Lau, V. A. Levanos, A. Sahasrabudhe, and F. E. Dewhirst.2001. Bacterial diversity in human subgingival plaque. J. Bacteriol.183:3770–3783.
24.Paster, B. J., W. A. Falkler, Jr., C. O. Enwonwu, E. O. Idigbe, K. O. Savage, V. A. Levanos, M. A. Tamer, R. L. Ericson, C. N. Lau, and F. E. Dewhirst.
2002. Prevalent bacterial species and novel phylotypes in advanced noma lesions. J. Clin. Microbiol.40:2187–2191.
25.Roˆc¸as, I. N., J. F. Siqueira, Jr., A. F. B. Andrade, and M. Uzeda.2003. Oral treponemes in primary root canal infections as detected by nested PCR. Int. Endod. J.36:20–26.
26.Sakamoto, M., Y. Huang, M. Umeda, I. Ishikawa, and Y. Benno.2002. Detection of novel oral phylotypes associated with periodontitis. FEMS Microbiol. Lett.217:65–69.
27.Siqueira, J. F., Jr.2002. Endodontic infections: concepts, paradigms and perspectives. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod.94: 281–293.
28.Siqueira, J. F., Jr., and I. N. Roˆc¸as.2003. Detection ofFilifactor alocisin
endodontic infections associated with different forms of periradicular dis-eases. Oral Microbiol. Immunol.18:263–265.
29.Siqueira, J. F., Jr., and I. N. Roˆc¸as. 2003. PCR-based identification of
Treponema maltophilum,T. amylovorum,T. medium, andT. lecithinolyticum
in primary root canal infections. Arch. Oral Biol.48:495–502.
30.Siqueira, J. F., Jr., and I. N. Roˆc¸as.2003. Positive and negative bacterial associations involvingDialister pneumosintesin primary endodontic infec-tions. J. Endod.29:438–441.
31.Siqueira, J. F., Jr., and I. N. Roˆc¸as.2004. Nested PCR detection ofCentipeda periodontiiin primary endodontic infections. J. Endod.30:135–137. 32.Siqueira, J. F., Jr., and I. N. Roˆc¸as.2004. Polymerase chain reaction-based
analysis of microorganisms associated with failed endodontic treatment. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod.97:85–94.
33.Siqueira, J. F., Jr., I. N. Roˆc¸as, and A. S. Rosado.2004. Investigation of bacterial communities associated with asymptomatic and symptomatic end-odontic infections by DGGE fingerprinting approach. Oral Microbiol. Im-munol.19:363–370.
34.Siqueira, J. F., Jr., I. N. Roˆc¸as, R. Souto, M. Uzeda, and A. P. Colombo.2000. Checkerboard DNA-DNA hybridization analysis of endodontic infections. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod.89:744–748. 35.Sundqvist, G., D. Figdor, S. Persson, and U. Sjo¨gren.1998. Microbiologic
analysis of teeth with failed endodontic treatment and the outcome of con-servative re-treatment. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod.85:86–93.
36.Thompson, J. D., D. G. Higgins, and T. J. Gibson.1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res.22:4673–4680.
37.Tronstad, L., and P. T. Sunde.2003. The evolving new understanding of endodontic infections. Endod. Top.6:57–77.
38.Weisburg, W. G., S. M. Barns, D. A. Pelletier, and D. J. Lane.1991. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol.173:697– 703.
39.Xia, T., J. C. Baumgartner, and L. L. David.2000. Isolation and identifica-tion ofPrevotella tanneraefrom endodontic infections. Oral Microbiol. Im-munol.15:273–275.