PARAMUTATION I N MAIZE DURING ENDOSPERM DEVELOPMENTI
K. S. McWHIRTER AND R. ALEXANDER BRINK
Division of Tropical Pastures, C.S.I.R.O., Mill Road, St. Lucia, Queensland, Australia, and Department of Genetics, Uniuersity of Wisconsin, Madison, Wisconsin
Received September 27, 1962
HE stages in development of maize plants at which paramutation of the R'
Tallele (colored aleurone, red seedling) occurs have not been fully defined.
BRINK (1959)
used trisomic plants of R'RStrg constitution to demonstrate that paramutation of R' is not dependent upon conjugation of R' with a paramuta- genic allele, such as R s t (stippled aleurone, green seedling), at zygotene of meiosis. Evidence has since been sought for occurrence of paramutation in vege- tative tissues and, specifically, during development of the endosperm of kernels of appropriate genotypes.There are two reasons for directing attention to the endosperm in these studies. I t is the first structure developed after fertilization in which R expression can be observed, and provides opportunity for early detection of the paramutation proc- ess. Secondly, the aleurone (the outermost layer of cells of the endosperm) is the only tissue, in the W22 inbred strain used in these investigations, in which R-
conditioned anthocyanin pigmentation is regularly manifest in a manner readily amenable to quantitative evaluation.
Paramutation of R' is not immediately evident, however, in the kernels result- ing from application of pollen from R'R' plants to R S t R s t or RmbRmb (marbled aleurone, green seedling) female plants, even though R' is paramutable and Rst
and Rmb are strongly paramutagenic alleles. The RrRstRSt and RrRmbRmb kernels resulting from these matings are as darkly pigmented as expected on the basis of additivity of the normal phenotypic effects of these alleles at the respective dosages involved (BRINK 1956;
WEYERS
1959). Only when plants are grown out from such RrRSt and RrRmb heterozygous seeds, and are then testcrossed on rgrg females, can it be wen that the pigmenting action of R' has been reduced below the standard value. The test in these latter cases, of course, is of an R' allele that has been heterozygous for a paramutagenic factor throughout a n entire sporo- phytic generation.It has now been found, however, that paramutation occurring during endo- sperm development can be detected if a weakly paramutant form of either the
R' or Rg (colored aleurone, green seedling) allele is tested. MCWHIRTER and
BRINK ( 1962) described weakly paramutagenic Rac alleles (self-colored aleurone, green seedling) which reduce the pigment-producing action of R', or Rg, only slightly below the standard value. The evidence summarized in 'this report shows
1 Paper No. 895 from the Department of Genetics, Agricultural Experiment Station, Univer-
sity of Wisconsin.
190 K. S . MCWHIRTER A N D R. A. BRINK
that such weakly paramutant R', or Rg, alleles are further reduced in pigment- producing action during the development of an endosperm i n which R', or RQ is heterozygous for a strongly paramutagenic allele.
MATERIALS A N D METHODS
The experiments were conducted with sub-lines of the uniform W22 inbred strain in which the appropriate R alleles had been incorporated by backcrossing. The R: allele, which was used in the tests for paramutation during endosperm development, is a mutant from standard RT that conditions green, rather than red, seedling (BRINK, BROWN, KERMICLE and WEYERS 1960) and colorless scutellum.
The RSc alleles, R;;, Rg",", and R;FY4, used to condition Ri prior to the test for paramutation in the endosperm are independently occurring mutants from stippled. They give self-colored aleurone, even in single dose, pigmented scutel- lum, green seedling, and are not paramutable in RsCRst heterozygotes. Ri; differs from the Ri: and R::4 alleles in paramutagenic action. Previous tests had shown that no alteration of R' could be detected following testcrosses of FIRrRi: indi- viduals on r g r g females. It was observed, however, that the pigment-producing
action of Rr was slightly, and significantly, reduced below the standard value if passed through R'R;; heterozygotes for three consecutive generations (MCWHIRTER 1961). R:;. therefore, is weakly paramutagenic, but its effect on
R" in F,R'R:i plants is subliminal. Ri: and R;'L, on the other hand, are overtly paramutagenic in F, heterozygotes with R', although only weakly so, as com- pared with the parent stippled allele (MCWHIRTER and BRINK 1962).
The test for paramutation during endosperm development involved comparison of the degree of aleurone pigmentation of kernels with endosperm genotypes of two generalized classes, Rg/r(I)/r(I) and Rg/r/r. The symbol r in the latter formula refers to the standard, nonparamutagenic rr and rg alleles, regularly giving colorless aleurone, and red and green seedling, respectively. The new symbol, r (I), in which 'I' may be read as "inducer of paramutation," represents a group of strongly paramutagenic alleles conditioning near-colorless aleurone, and either green or red seedling, derived earlier by ASHMAN (1960) from
R'RSt9 X rgrQ6 matings. Two such factors were employed in the present ex- periments, r ' ( I ) 3 and r g ( I ) 4 . The former conditions red seedling and anthers and the latter, green seedling, rarely with fine red stripes on roots and coleoptile, and green anthers sometimes mottled with red. Most kernels homozygous for these factors are colorless, but occasional seeds in each case show one or more small, widely scattered, purple spots, hence the designation "near-colorless." Evidently both alleles are slightly unstable in their pigmentation effects in plant and aleurone tissues. Tests made in the conventional way ( r g r g 0 x R"/r (I) 8
testcrosses) showed that rT (I) 3 and rg (I) 4 are paramutagenic in heterozygotes
P A R A M U T A T I O N I N E N D O S P E R M 191
instability in paramutagenic action, in addition to that observed in color ex- pression, has not been excluded.
The difference that is important in the present context between the colorless
rT and rg alleles, on the one hand, and the near-colorless r ' ( I ) 3 and rg(JJ4
factors, on the other, is that the former are nonparamutagenic, whereas the latter are strongly paramutagenic. If, therefore, paramutation in the endosperm of Rg in either standard or modified form is detectable it would be expected to result in the Rg/r (I) / r ( I ) class of kernels being consistently less pigmented than the Rg/r/r class of kernels. A significant advantage in using a paramuta- genic near-colorless allele, rather than a spotting factor like stippled, for example, in tests for paramutation in the endosperm, is that the aleurone pigmenting effect inherent in r (I) itself is negligible.
The following W22 genotypes were used to determine whether paramutation occurs in the endosperm:
Pistillate parents Staminate parents
rr (I) 3/rg R; R;
rr/rg
(I)
4 R; R;;rr/rg R; R;;
R: RE4
With these stocks each of the two generalized classes of kernels desired,
R;/r (I) /I( I ) and R;/r/r, was obtainable on the same ear, and so under the same
environmental conditions, and the classes could be distinguished by observing seedling color following germination of the kernels. The first two pistillate strains were heterozygous for a paramutagenic and a nonparamutagenic, color- less (or near-colorless) allele, distinguishable from each other, under appropriate conditions, by effect on seedling color. The third pistillate stock, r'rg, was used to test for possible influence of the seedling color markers, as such, on aleurone phenotype. The first genotype listed among the staminate parents provided a n
R: allele in standard form from a homozygote, and the second from a heterozy- gote carrying a very weakly paramutagenic Rsc mutant with a subliminal effect
in F1RgR;: heterozygotes. The R: allele derived from two remaining staminate
strains was slightly reduced in pigment-producing action by virtue of passage through heterozygotes carrying the weakly paramutagenic R;; and R",4 factors. As will be seen later, paramutation in the endosperm could be detected with certainty only when the R: factor tested was introduced into the tissue in this conditioned form.
Single pollen samples from individual plants of each of the four staminate types were spread over one ear shoot of each of the three classes of pistillate parents. Each set of ears obtained thus represented two matings which served to test for paramutation of R: during endosperm development, plus a third mating on a control r'rg female.
The ears were assigned code numbers at random prior to scoring, and the coded samples were then scored in numerical order.
192 K. S . MCWHIRTER A N D R. A. BRINK
Rg/-/- kernels were separated from the Rae/-/- kernels on the basis of aleurone
and scutellum color. The Rg kernels were distinctly mottled and the scutellum tissue in the embryo was colorless, whereas the endosperm in the Rae kernels was completely colored and the scutellums also were strongly pigmented.
The Rg/-/- kernels only were then scored in detail for aleurone pigmentation.
This was done by matching individual seeds against a set of standard kernels defining seven aleurone color classes. The latter ranged from Class 1 (colorless) to Classes 6 (dark mottled) and 7 (completely pigmented). The kernels from a given mating were thus scored in order, and were then germinated in the same sequence for determination of seedling color. Genotype of the endosperm was inferred from the color of the resulting seedling. For example, following the mating rr (I) 3/rg 0 x RiR: 8 the seeds yielding red seedlings ( R:/rr (I) 3 embryos) possess R:/rr (I) 3/r' (I) 3 endosperms, whereas those giving green seedlings ( R:/rg
embryos) have R:/rg/fl endosperms. The mean aleurone color scores were based on the first 50 kernels of each genotype, as read in order in the germinating pan. The scoring procedure is without bias, since determination of the endosperm genotype was accomplished retroactively with respect to the scoring of aleurone color.
Some matings failed to yield 50 kernels of each genotype. In these instances the mean scores were computed for 25, or 40 kernels, as available. The resulting data are entered in the tables but were not utilized in computing general mean differences or in determining the statistical significance of differences.
EXPERIMENTAL RESULTS
Matings inuolving staminate parent RER;: The data in Table 1 provide no clear
evidence one way or the o,ther, for paramutation in the endosperm of the R!
allele i n standard form, derived from the homozygote. The mean aleurone color scores for all matings are high, and might well be considered estimates of the pigment-producing action of unchanged R,".
There is a n indication, however, that R< has undergone paramutation in the TABLE 1
Rg aleurone color scores following application of pollen from R;/Rsc plants to individual female plants heterozygous for a near-colorless, paramutagenic r(I), and a colorless,
nonparamutagenic r allele
83
Male (a) Female parent r r ( I ) 3 / r g (b) Female parent r r / r g ( I ) 4 ( c ) Female parent rr/rg
plant
No. R y ( I ) r r ( I ) R y g r g Diff. Rgr'F R Y ( I ) f l ( I I Diff. Rgrrrr Rgrgrg Diff.
18 5.88 5.96 -0.08 5.86 5.84 +0.02 6.00 5.88 +0.12
21 5.90 5.98 -0.08 5.98 5.96 +0.02 5.98 5.9.1. +0.04
24 5.82 5.96 -0.14 6.00 6.00 0.00 5.98 5.9% +O.OZ
Means 5.888 5.960 -0.072 5.940 5.884 +0.056 5.968 5.940 +O.OB Significance t, = 3.47, P
<
0,M t, = 1.73, P>
0.119 5.96 5.98 -0.02 5.92 5.74 +0.18 5.92 5.96 -0.04
22 5.88 5.92 -0.04 5.94 5.88 +O.M 5.96 5.96 0.00
P A R A M U T A T I O N IN E N D O S P E R M 193
R,”/rT (I) / r r (I) kernels resulting from the r r (I) 3/rg female x R,” R,” male mating. The mean score for this class of seeds is slightly less than that for the RZ/+’/fl
seeds on each of the five testcross ears, and the differences between the group means, 0.072 units, is significant at the five percent level. Since this difference, however, corresponds to the relative displacement of only 18 kernels among the
250 of each genotype scored the evidence for paramutation is not persuasive. Similarly, the score for the R,”/rg (I) 4/rg( I ) 4 kernels is not greater on any one of the five testcross ears than that for the corresponding R,”/rT/rr seeds, following
the rg(1)4/rr? x R: R:8 matings. The difference in group means in this case,
however, is small and clearly is nonsignificant statistically. It may be concluded that, whereas the aleurone color scores for the R; testcross kernels when R: is derived from R,” R,” 6 plants are in the direction that would be expected if r’ (I) 3
and r g ( I ) 4 caused the allele to paramutate in heterozygous endosperm cells, the
data do not unquestionably establish such an effect.
The closely similar scores recorded in Table 1 for the Rfrrrr and R;rgrO kernels from the rrrgO x R; R,” matings show that the plant color markers used to identify the two classes of seeds on the testcross ears do not affect aleurone color. This relation is confirmed by the results from the corresponding tests reported in Tables 2 and 3.
Matings involving staminate parent R; R;;: The data on the paramutation of
R: in the endosperm following extraction of the allele from FIR: R”,”, plants, as shown in Table 2, parallel those for R; derived from R; R; homozygotes. This
result is consistent with the fact previously mentioned that, whereas R:: is very weakly paramutagenic over several sporophytic generations, its effect on R’ in
F, RrRi; heterozygotes is subliminal. It would not be unexpected, therefore, if
Ri
derived from Rt R”,”, and R; R: plants reacted alike in the present tests.Again, the R:/r( I) /I (I) testcross kernels from the matings on both rr ( I ) 3/rg
and 7” ( I)4/rg females are slightly, but consistently, less pigmented than the
corresponding R:/r/r control seeds. The difference, even if small, is significant at the two percent level in the former instance. Although suggestive of such an
TABLE 2
Rg aleurone color scores following application of pollen from R;/R;; plants to individual female plants heterozygous for a near-colorless, paramutagenic r(I), and a colorless,
nonparamutagenic r allele
Male (a) Female parent r7(1)3/r9 (b) Female parent r‘/rg(I)4 (c) Female parent r*/rn
plant R y R;@@ Diff.
R W Y ’ Ro@(I)rg(I) Diff.
6 6
No. RWp(I)rr(I) R:@@ Diff.
4 6.00 6.02 -0.02 6.00 5.94 +0.06 5.98 6.00 -0.02
7 6.02 6.04 -0.02 5.98 5.84 +0.14 6.00 5.94 +0.06
15 6.02 6.02 0.00 6.02 5.94 +0.08 6.00 6.00 0.00
17 5.98 6.08 -0.10 6.06 6.02 +0.04 6.02 6.06 -0.04
20 5.96 6.00 -0.W 6.00 5.96 +0.04 6.06 6.10 -0.04
Means 5.996 6.032 -0.036 6.012 5.940 +0.072 6.012 6.020 -0.008
194 K . S. MCWHIRTER A N D R . A. B R I N K
effect, these data do not prove that Rt derived from R: Ry8individuals undergoes paramutation in endosperm cells which also carry a paramutagenic r(1) allele. Matings inuoluing staminate parents R; R;:4 and R: R;;: I n contrast with the results obtained with R: R: and R3 R:; plants, the series of matings involving the staminate parents R: R:;& and RY RZ provide definitive evidence of paramutation of RQ during endosperm development. The data are reported in Table 3, Section A, for matings involving the R: RSYh male parent, and in Section B for those relating to the R: RiZ male parent. As noted earlier in the present paper, R:; and R;:b were known from previous experiments, to be weakly, although detectably, paramutagenic in F, heterozygotes with standard R', in contrast with RE whose action in this respect in corresponding F, plants is subliminal.
The data for matings of R: R:;&and R: RZ male parents on rr(1)3/rg and
r g / r g ( I ) 4 female parents show that a difference in degree of aleurone pigmenta-
tion was detected regularly in the comparison of RQ/r (I) / r (I) and Rg/r/r kernels developed on the same ear. The kernels of Rg/r(I)/r(I) endosperm genotype in each comparison were less pigmented than the kernels of RQ/r/r endosperm geno- type, irrespective of whether paramutagenicity was associated with red seedling color (as in r? (I) 3) or with green seedling color (as in rQ (I) 4).
TABLE 3
R: aleurone color scores following application of pollen f r o m R; R;;4 and R.g/R<;; plants (R;;, and Rfic are weakly paramutagenic) to indiuiduals heterozygous for a near-colorless,
paramutagenic r(l), and a colorless, nomparamutagenic r allele 99
A . Male parent=RgS Rsci34
R L l e ( a ) Female parent r T ( I ) ? / r q ( b ) Female parent r r / r g ( 1 ) 4 ( c ) Female parent 1 1 / 1 9
plmrt - -
No. R g r r ( I ) r l ( I i Rgrgrg Diff. R.'Jrrr'' R g r g ( I ) r g ( I ) Diff. R.rrl-rV Rgrgrg Diff.
6 5 5 5 5 6
- ~~ ~~~
2 4.90 5.54 -0.64 5.60 4.92 t 0 . 6 8 5.60 5.42 $0.18
5 5.26 5.66 -0.40 5.14 4.78 +0.36 5.46* 5.27* +O.l8
10 5.66 5.76 -0.10 5.70 5.48 +0.22 5.70 5.64 +0.06
12 5.10 5.38 -0.28 5.43* 5.10* $0.33 5.28 5.24 +0.04
15 5.48 5.82 -0.34 5.64 5.32 +0.32 5.58 5.46 4-0.12
17 3.92 4.24 -0.32 4.241 4.08t t 0 . 1 6 4.26 4.24 t 0 . 0 2
20 Ear not obtained 4.42 3.86 +0.56 4.08 4.02 +0.06
24 5.10 5.32 -0.22 5.62 5.30 +0.32 5.34 5.30 +0.04
25 5.40 5.66 -0.26 5.80 5.44 t 0 . 3 6 5.65* 5.50* +0.15
Means 5.004 5.358 -0.354 5.348 4.878 t 0 . 4 7 5.058 5.028 t 0 . 0 3
Significance t, = 5.97, P < 0.001
19 4.22 4.84 -0.62 4.86 3.92 +0.94 4.62 4.90 -0.28
t7 = 5.41, P < 0.001 t, = 0.2.0, P
>
0.8B. Male parent=RG RsC99
2 4.54 5.20 -0.66 4.38 4.18 t O . 2 . 0 4.92 5.12 -0.20
3 4.56 4.78 -0.22 4.34 3.98 +0.36 4.58 4.80 -0.22.
4 4.52 4.86 -0.34 4.16 3.52 +0.64 4.38 4.44 -0.06
8 4.52 4.90 -0.38 4.44 3.98 t 0 . H 4.86 4.64 +O.Z2
Means 4.535 4.935 -0.40 4.330 3.915 t 0 . 4 1 5 4.685 4.750 -0.065
t, = 4.51, P
<
0.05 t, = 0.64, P>
0.05Significance t, = 4.30, P
<
0.05* I
P A R A M U T A T I O N I N E N D O S P E R M 195
The mean differences between R g / r ( I ) / r ( I ) and Rg/r/r kernels were both significant at the 0.001 level for matings involving the
Ri
R::, male parent, and at the 0.05 level for matings involving the R: R:: male parent (where fewer male plants were tested). The differences in score between endosperm genotypes in each of the comparisons were not large, but in view of the regularity of the results it is not surprising that the mean differences were highly significant.In contrast to the results from the matings on r'(1) 3/r9 and r r / r g ( 1 ) 4 females, the control matings of the same male plants on r'/rg females showed no consistent difference in aleurone color between Rgrrrr and Rgrgrg kernels. The difference between class scores varied appreciably from ear to ear in this test, but were random in direction, and the mean differences were not statistically significant. An essential feature of these experiments is that the Ri gametes derived from
R; R;:, and R{ R;; male plants were slightlv reduced in pigment-producing potential relative to the level for standard R:. The reduction is reflected in the fact that the mean aleurone color scores for the R i / r r / r r and Ri/rg/rg classes of kernels were consistently lower in matings involving the R; R",4 and R,"
R:;
male parents (Table 3 ) than in those involving the Ri R: male parent (Table 1 ) . Paramutation of the R: factor during the development of R ; / r ( I ) / r ( I ) endo- sperm kernels was detected, therefore, with R: alleles that previously had undergone weak paramutation in R: Ry;, or R," R,"," heterozygotes.The data in Table 3, Section A, also show that there were differences between individual R: R;Z4 parent plants i n the level of paramutation induced in the R;
factor. The differences between male parent plants were highly consistent over all three classes of pistillate parents used in the matings. This variation in level of paramutation of RZ is of no statistical consequence for the present experiment because it is partitioned out by the "t" test which was applied to the difference between paired mean scores. It is noteworthy, however, that the magnitude of the difference between endosperm genotypes was not related to the level of para- mutation occurring in the R; R;;, male parent plant.
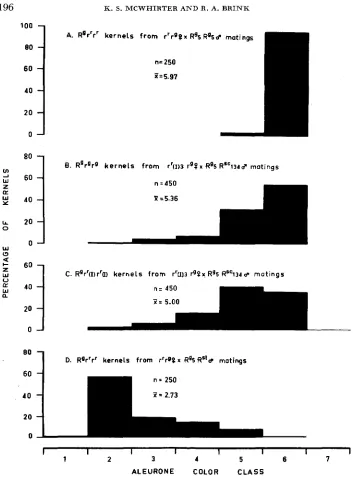
The principal features of the experimental results obtained with the R: allele, as just recounted, are illustrated in Figure 1. The slight conditioning effect on
Ri
196 K. S. MCWHIRTER A N D R. A. BRINK
100
40
20
0
80
20
0
60
20
0
A. Rgrrr' k e r n e l s f r o m r ' r g t x Rg5 R g 5 d matings
1
n= 250z-5.97
B. Rgrgrg k e r n e l s f r o m r'lI)3 r g ? x Rg5 R'c134d matings
C. Rgrrmrra) k e r n e l s f r o m r'im rg$xR% RSCi34d matings
D. Rgr'r' kernels from rrrg% x RgS RStd matings
1
n = 250
Y = 2.13
20
0
I I I I I I I 1
1 2 3 4 5 6 7
ALEURONE COLOR C L A S S
PARAMUTATION IN ENDOSPERM 197
tation of R: directly by
R",",,
(polygon B ),
and of conditioned R: during endosperm development (polygon C),
involved only moderately small reductions in pig- ment-producing potential relative to that which R: is capable of undergoing(polygon
D)
-
Paramutation during endosperm development of the
Rr
allele: Data from a nunexpected source indicate that the R' factor also undergoes paramutation during endosperm development.
The data were obtained by comparison of the aleurone pigmentation of R r / r r
(I) 3/r' ( I ) 3 and R r / r r / r r kernels from the matings rr (I) 3/r' ( I ) 3 0 X Rr/Rt: male
and r'r+ 0 X R'/R;i male, respectively. The two classes of pistillate parents were
closely related, but were not sibs. The R'/RE male plants used were F3 R'
R::
heterozygotes derived by self-pollination of a single R'/RZ F, plant. Separation of R r / r r ( I) 3 / r r ( 1 ) 3 from R",/r' ( I ) 3 / r r ( I) 3 kernels, and R r / r r / r r from Ri:/r'/r+ kernels was made on the basis of aleurone pigmentation only. Table 4 con-
tains the mean scores obtained for 50 R r / r r ( I ) 3 / r r ( 1 ) 3 and 50 Rr/rr/rr kernels from each of the respective pairs of matings involving a single R' R,":
F3
male plant.A consistent difference was found between the mean values for the two classes
of kernels, and in each comparison the R'/rr ( I ) 3 / r r ( I ) 3 endosperms were the less pigmented. Figure 2, which is a photograph of a pair of ears involving a single R' RE F, male parent, illustrates the extent of the observed difference in pigmentation.
The possibility exists of some bias in these data resulting from incomplete separation of the R'/r/r from R i ; / r / r kernels, although this seems unlikely in view of the fact that no significant deviations from the expected 1: 1 ratio were observed. The magnitude of the differences in score between Rr/r+ ( I ) 3 / r r (I) 3
and R r / r r / r r kernels suggest that the R' factor was reduced in pigment-producing
action during the development of the R r / r r
(I)
3 / r r ( I ) 3 kernels. Thus it appearsTABLE 4
Rr aleurone color scores following application of pollen from Rr R;: F3
plants to r r (I) 3/rr (I) 3 and rrrr female plants
Male plant No.
2 3
4
7 13 16
18
28 30
Mean Significance
Female parent=rP/r' Ftmale parent=rr(I)3/r*(I) 3
RprrrV R 3 ' ( I ) 3 r 7 ( I ) 3
kernels kernels Difference
6.00 5.66 +0.34
6.00 5.78 +0.22
5% 5.56 +0.40
5.94 5.48 +0.46
6.04 5.70 +0.34
6.00 5.80 + O . N
5.80 5.38 +0.42
6.02 5.72 +0.30
6.02 5.66 +0.36
5.975 5.637 +0.338
198 K . S. MCWHIRTER A N D R. A. BRINK
FIGURE 2.-I:ars rrsultirlg from niiitings of a single R'/R;: F:, malr plinnt on an r'/r' female (left) ancl nn r ' ( I ) 3/r'(I)3 female ( r i g h t ) . Each ear contains approximately one.half R : ; / r / r kernels. which t i r e completely rolorrcl. The R r / r r / r r kernels on the ear at left are very darkly mottled, whereas the Rr/rr(I)/rr(I) kernels on the rar at right an- distinrtlv mottled. the difTerenre in aleurone pigmentation k i n g due to paramutation of the R r nllelr orrurririg during endosperm development.
probable that the R r factor, in addition to the R: mutant from R', is responsive to paramutation during endosperm development.
These results are in seeming contrast with those presented earlier for matings involving the RY Ry, F, male parent. The difference is referable. however, to the number of generations during which RY and R;; were propagated in heterozygotes before the test was made. The results presented in Table 2 showed that RPJ from
R:/R:: F, male plants was not detectably altered in pigment-producing potential
relative to standard R:. nor was R: from this source altered during the develop- ment of R9/rr( I ) 3 / r r ( I ) 3 endosperms. T h e data in Table 4, on the other hand. show that following three generations of heterozygosity of R' with RiS. R' was altered in a manner that made the allele detectably responsive to the para- mutagenic effect of r'( 1)3 during Rr/rr( I ) 3/rr( I ) 3 endosperm development.
Erjidence excluding participation of the specific modifier ( M I ' ) of stippled in
paramutation during endosperm development: ASHMAN ( 1960) showed that the
PARAMUTATION I N ENDOSPERM 199 over units distal to the R locus. M S t has the effect of markedly increasing the number of pigmented spots in the aleurone, in, for example, the comparison of
R s t M S t / r r M s t / r r M S t with R S t m s t / r r m s t / r r m s t kernels. The near-colorless, para- mutagenic alleles ( r r ( I ) 3 and r Q ( I ) 4 ) used in the present experiments were linked with M S t , whereas the colorless, nonparamutagenic alleles ( rr and r g )
were linked with the null element, symbolized by mst (ASHMAN 1960). The stippled modifier, M s t , has unusual properties, including transposability (ASH-
MAN 1960). Earlier experiments had disclosed no relation between
M s t and
paramutation at the R locus but it seemed desirable to test directly for a possible effect during endosperm development. The result was again negative, as the following evidence shows.The relevant data were derived from an experiment which utilized, in principle, the same procedures as outlined for the demonstration of paramutation during endosperm development. Matings of individual R f / R s male plants on
rrMSt/rgmst, rrmst/rgMst and rrmst/rgmst female parents were made, the r r and
rg alleles in all cases being nonparamutagenic. Comparison of the aleurone phenotypes of R i / r r M s t / r r M s t with R!/rgmst/rgmst, R!/rrmst/rrmst with R f /
rgMSt/rgMSt, and R5g/rrmst/rrmst with Rz/rgmst/rgms kernels, respectively, from
these matings were then made. The data, entered in Table 5, show that there were no significant differences in any of the comparisons. The difference in M S t constitution between the r ( 1 ) and r stocks used in the experiments on paramutation in the endosperm, therefore, is irrelevant.
DISCUSSION
The evidence presented here together with that from earlier, less direct, tests
(BRINK 1959, 1960) demonstrates that paramutation at the R locus in maize occurs in vegetative cells. The data also provide the basis for certain inferences concerning the nature of the paramutation process. The most important addi- tional conclusion is that the paragenetic changes affecting R action are cumula- tive and there appears to be a positive relationship between degree of paramuta- tion and duration of vegetative development.
TABLE 5
R; aleurorze color scores following application of pollen from Rf/RSt
plants to r r M S t / r g , r ' . / r 0 4 S t and r P / r g female plants+
(a) Female parent r r M v t / r g
RggT'T' R;rp.rU
Family No. of male
no. plants tested kernels kernels Diff.
M742 5 3.90 4.13 -0.23
Mean 10 3.48 3.70 -0.22 Significance t, = 1.75, P
>
0.1-
M743 5 3.05 3.26 -0.21
(b) Female parent r r / r g M a t
Rgrrrr Rp.rqrp. R;rrrp R F
kernels hernels Diff. kernels kernels Dlff (c) Female parent rr/rU
_____
-
-
____-
3.49 3.50 -0.01 3.59 3.77 -0.18 2.79 2.82 -0.03 2.73 2.71 +0.02 3.14 3.16 -0.16 3.16 3.24 -0.08
t, = 0.43, P
>
0.5 & = 0.86, P>
0.3200 K. S . MCWHIRTER A N D R. A. BRINK
The pistillate plants used in the matings to determine whether paramutation occurs in the endosperm were heterozygous for a colorless, or near-colorless, strongly paramutagenic factor (rr (I) 3 or rQ (I) 4) and a colorless, nonparamuta- genic factor (rr or r g ) . The criterion of paramutation was a reduction in the
pigment-producing action of a n R y allele, introduced through the pollen, in
R+ (I) r (I) but not in Ryr r kernels randomly distributed, although separately recognizable, on the same ears. The Rg allele employed (R:) was introduced at fertilization into the nucleus of the central cell of the female gametophyte (from which the endosperm then develops) in three states. The first of these repre- sented
Ri
in its standard condition, as obtained from an R: R: stock culture. I n the second class of matings R; was extracted from the heterozygotes with Rji, a very weakly paramutagenic mutant from stippled whose paramutagenic action in F,RYRii plants previously had been shown to be subliminal in testcrosses onr g r y females. Thirdly, R: was derived from R; RiZ or R:
Rtih
plants, in both ofwhich its pigment-producing potential is slightly, but measurably, reduced in a single generation by virtue of the weak paramutagenic effect of the similarly acting Ri," and R;;h mutants from stippled. The conditioning of R$ as a result of passage through R: R;," or R: R;;, plants, proved to be important for the detection of paramutation in endosperms carrying an r (I) factor.
The entire series of test matings makes possible eight independent comparisons of the immediate effect on R: pigmenting action of heterozygosity in endosperm cells for the paramutagenic r(1) and the control, r factors. In each of the eight cases the mean aleurone color score for the R:/r( I) / I * (I) kernels was found to be lower than that of the corresponding R : / r / r seeds. This is the result expected, of course, if R: undergoes paramutation in the endosperm.
When the data are examined according to R< source, however, only in the cases in which the allele was introduced into the endosperm in conditioned form (from R:
RZ
and R: R;;4 heterozygotes) was the difference in mean score betweenRt/r( I) / r (I) and R f / r / r kernels readily detectable and statistically significant. Standard R;, from R: R; plants, and R; extracted from R; R;; F, individuals gave no greater reduction in color score when made heterozygous with the para- mutagenic r(1) allele than would result frequently from random sampling. This result could mean either that standard R:, in contrast with conditioned R f , is not paramutable in the endosperm or that the latter allele merely lies closer to the threshold at which paramutation is expressed as an overt change in aleurone phenotype. The direction of the changes creates a certain presumption in favor of the latter conclusion, but it is evident from the small magnitude of the differ- ence observed in these experiments that matings involving standard R; are un- likely, even in a large scale test, to provide convincing evidence for paramutation during endosperm development. In summary, therefore, the data on conditioned
Rf provide proof of paramutation in the endosperm, and those on standard
R;, while inconclusive, point to the same result,
P A R A M U T A T I O N I N E N D O S P E R M 20 1
previous observations that the phenomenon is progressive. MCWHIRTER and
BRINK (1962) found that R' extracted from heterozygotes with very weakly
paramutagenic alleles was further reduced in pigment-producing potential if it was then passed through a heterozygote with stippled, a strongly paramutagenic allele. Presumably a common underlying mechanism is involved in these se- quential alterations of R action.
Unpublished data obtained by ASHMAN (1959) show that the two near- colorless alleles originally derived from R' Rat 0 x r g r g 8 matings, and here termed
r ' ( I ) 3 and rQ(1)4, are about as strongly paramutagenic as stippled when tested
in the conventional way. That is to say, when pollen from R' Rat, Rr/r"(I) 3, and R'/rg(I)4 plants is applied to r g r g females the resulting R'rgrg kernels, in all
three cases, are markedly reduced in pigmentation as compared with the R'rgrg seeds from r g r g ? X R'R'6 control matings. I n contrast with these results, the immediate effect of introducing R; (which is equivalent to standard R' in aleurone pigmenting effect) into endosperms carrying either r ' ( I ) 3 o r rg(1)4 is to lower the aleurone color score by an amount that is so small as to be statistically nonsignificant. Even when conditioned R;, from R: RiZ or R," R::,, plants, is similarly tested, the effect on aleurone color, while statistically significant, and readily detectable, is still relatively small. Obviously, the change in pigment- producing potential that R; undergoes on being introduced directly into endo- sperm cells containing a paramutagenic factor is much less than that occurring when its counterpart, R', is passed through sporophytes heterozygous for one of the same paramutagenic alleles. This is despite the fact that the endosperm, in these tests, contains two effective paramutagenic alleles and an R; factor that is already weakly paramutant. The quantitative shift in the distribution of
R:/r'(I)/r(I) kernels relative to that of R : / r / r kernels (polygons C and B,
respectively, in Figure 1) suggests further that paramutation does not occur as a discrete change at some distinct stage in endosperm development. Rather, the results imply that the change in pigment-producing potential recognized as paramutation is the cumulative result of small changes which occur sequentially throughout somatic cell development. The expectation follows that degree of paramutation should progressively increase as the number of cell generations increases during development of the sporophyte.
202 K. S. MCWHIRTER A N D R. A. BRINK
maize plant, with its single, apical, staminate inflorescence, is not well suited for determining, by conventional testcrosses, whether paramutation is progressive as development of the individual proceeds. It has been shown, however, that the very weakly paramutagenic factor, RiS, referred to earlier in this paper, is not detectably paramutagenic in F, R'R;; plants, but that the R' allele derived from
F, R' R;; individuals, in a self-pollinated lineage, is clearly paramutant ( MC-
WHIRTER 1961). This fact may be construed as evidence for the progressive occurrence of paramutation through three sexual generations. The present evi- dence, from a comparison of endosperm and sporophyte, is not decisive on this point but it is in harmony with expectation if there is a positive relation between number of somatic cell divisions and degree of paramutation.
SUMMARY
Following the demonstration by BRINK (1959) that paramutation of the R'
allele (colored aleurone, red seedling) was not dependent upon conjugation of R'
with a paramutagenic allele at zygotene meiosis, evidence has been sought for occurrence of paramutation in vegetative tissues.
Paramutation during endosperm development of an RQ allele conditioned in a particular way was demonstrated by comparison of the aleurone pigmentation of kernels of endosperm genotypes Rg/r (I) / I (I) and Rg/r/r. The comparison
was made in matings in which the pistillate parents were heterozygous for a near-colorless, strongly paramutagenic factor ( r r (I) 3 or I Q ( I)4) and a colorless,
nonparamutagenic factor (r' or I"). Paramutation, if it occurred during endo-
sperm development, would be expected to result in reduction of the pigment- producing action of the Rg allele, introduced through the pollen, in the Rg/r (I)
/
I (I) kernels relative to the RQ/r/r kernels developed on the same ear.
The RQ allele, used in the tests for paramutation during endosperm develop- ment, was introduced into the primary endosperm nucleus at fertilization in three states:
( 1 ) Standard RQ from an RgRg stock culture.
( 2 ) Rg from Rg/R;t F, heterozygotes. where 22,"; is a very weakly paramuta- genic mutant from RSt (stippled aleurone, green seedling). The paramutagenic action of Ri; on RQ in Rg R:: F, plants had previously been shown to be subliminal i n testcrosses on r I females.
( 3 ) Conditioned RQ from RQ RE and RQ R& plants. R;; and R"I",, are inde- pendently derived, weakly paramutagenic mutants from RSt, and in both of these heterozygotes RQ was slightly, but detectably, reduced below the standard level of pigment-producing action. This reduction is the essential feature of the conditioning process.
The source of As, in terms of the conditioned state, proved to be important for the detection of paramutation in endosperms carrying an r ( I ) factor. Only in matings in which RQ was introduced into the endosperm from RgR;; and
P A R A M U T A T I O N I N ENDOSPERM 203 and statistically significant. With standard Rg, from RgRg plants, and Rg from
Rg R,"; F, plants, the differences between Rg/r (I) / r ( I) and Rg/r/r kernels were always in the expected direction, but were of small magnitude and were statistically nonsignificant.
Paramutation of Rg during endosperm development was clearly shown by the secondary alteration in pigmenting potential of weakly paramutant (condi- tioned) Rg factors derived from Rg R:: and Rg R",4 heterozygotes. It is suggested that paramutation involves cumulative genetic changes affecting the pigment- producing action of Rg occurring sequentially during somatic development. The data are consistent with the expectation of a positive relation between degree of paramutation of Rg and the number of somatic cell generations in which RQ is heterozygous for a paramutagenic factor.
ACKNOWLEDGMENTS
This study was aided by grants from the Research Committee of the Graduate School, University of Wisconsin, of funds supplied by the Wisconsin Alumni Research Foundation, and by grants from the National Science Foundation. The work was carried out during tenure by K. S. MCWHIRTER, of a C.S.I.R.O. Over- seas Studentship held jointly with a Fellowship from the Department of Genetics, University of Wisconsin.
LITERATURE CITED
ASHMAN, R. B., 1960 BRINK, R. A., 1956
Stippled aleurone in maize. Genetics 45: 19-34.
A genetic change associated with the R locus in maize which is directed and
Paramutation a t the R locus in maize plants trisomic f o r chromosome 10. Proc. Natl. Sci.
Paramutation and chromosome organization. Quart. Rev. Biol. 35: 120-137. potentially reversible. Genetics 4 1 : 872-889.
U.S. 45: 819-827. 1959
1960
BRINK, R. A., D. F. BROWN, J. KERMICLE, and W. H. WEYERS, 1960 paramutant R phenotype in maize. Genetics 45: 1297-1312. MCWHIRTER, K. S., 1961
allele i n maize. Ph.D. thesis. University of Wisconsin Library. Madison, Wisconsin. MCWHIRTER, K. S., and R. A. BRINK, 1962
R locus i n maize. Genetics 47: 1053-1074.
WEYERS, W. H., 1959
Locus dependence of the
Paramutation studies with self-colored-mutants from the stippled
Continuous variation in level of paramutation at the