Red
Cell
Oxygen
Affinity,
Hemoglobin
Type,
2
,3-Diphosphoglycerate,
and
pH as a Function
of Fetal
Development
Harry
Bard,
MD, and
Francois
Teasdale,
MD
From the Perinatal Service and Research Center of H#{243}pitalSainte-Justine, Department of Pediatrics, University of Montreal, Montreal, Quebec, Canada
ABSTRACT. Studies were carried out on fresh cord blood
obtained at delivery from nonstressed normal fetuses ranging from 24 to 42 weeks of gestation, to determine the relationship of 2,3-diphosphoglycerate (DPG), the intracellular red cell and extracellular pH, and the
pro-portions of adult and fetal hemoglobin in regulating the
position of fetal red cell oxygen affinity in utero. There
was a significant positive correlation between P50 and gestational age (r = .62, P < .001), the linear regression
increased from 17.8 to 22.5 mm Hg. There was also a
significant positive correlation between P50 and the
per-centage of adult type hemoglobin (HbA) (r = .67, P <
.001). In contrast gestational age had no effect of 2,3-DPG
levels, the mean and SD was 14.86 ± 2.04 mol/gm of Hb
or ipH between plasma and red cell, the mean was 0.187
± SD 0.032. However, there was a significant negative
correlation between the intraerythrocyte hydrogen ion
concentration and DPG level (r = .5, P < .025). It is
concluded therefore that the decrease in fetal oxygen
affinity as gestation progresses is related mainly to the
increase in the amount of HbA and the levels of DPG or
zpH between plasma and red cells are not a function of
gestational age. Pediatrics 64:483-487, 1979; fetal 02 af.
finity, 2,3.diphosphoglycerate, fetal hemoglobin, adult
hemoglobin.
In all mammals so far investigated the oxygen
affinity of fetal blood is higher than that of adult
blood. After birth the oxygen affinity decreases to
the adult levels. The time course of this decrease in
oxygen affinity varies with species. Although the
difference in oxygen affinity between human fetal
and adult blood has been known since 1931,’ the
mechanism responsible has only been clarified
dur-ing the last decade. It is mainly due to the intrinsic
Received for publication Nov 9, 1978; accepted Jan 26, 1979. Reprint requests to (H.B.) H#{243}pitalSainte-Justine, 3175 Chemin Sainte-Catherine, Montreal, Quebec, Canada H3T 1C5. PEDIATRICS (ISSN 0031 4005). Copyright © 1979 by the American Academy of Pediatrics.
structure of the hemoglobin molecule and the
spe-cific action of2,3-diphosphoglycerate (DPG) on the
hemoglobin molecule2’3 and the unspecific effects of
changes of the red cell pH.4’5
In the human fetus, adult hemoglobin appears
around eight weeks of gestation6 but it is after 30
weeks when there is an acceleration in the
switchover from the fetal type hemoglobin (HbF)
synthesis to the adult type (HbA).7 At five months
after birth HbA synthesis has almost completely
replaced HbF, and only a small residual synthesis
of HbF persists for the rest of life.8
Several studies9” have been reported on changes
of oxygen affinity in newborn infants. They agree
on one hand that fetal oxygen affinity decreases as
gestation progresses, but on the other hand the
interrelationship of adult and fetal hemoglobin,
2,3-DPG, and intracellular pH on red cell oxygen
affin-ity during life in utero remains unclear. Also, in a
recently completed study in the sheep fetus, it was
demonstrated that the level of DPG is not
influ-enced by gestational age’2 and the decrease in fetal
blood oxygen affinity as gestation progresses can be
attributed to the amount of adult hemoglobin
pres-ent in the fetal circulation. In other similar
stud-ies’3”4 in mammals whose red cells do not have a
switchover of hemoglobin type during the perinatal
period, there was no evidence of an increase in fetal
red blood cell DPG.
Since regulation of fetal oxygenation is of utmost
importance in perinatal medicine, a study was
planned to relate the changes in utero in human
fetal oxygen affinity as gestation progresses with
the 2,3-DPG concentrations, pH between plasma
and red blood cells, and the proportions of adult
and fetal hemoglobin. This was done by using
pla-cental cord blood obtained immediately at birth
from viable nonstressed fetuses of varying
24-
22-a’
I 20-E E
18-0 0.
16-
14-S
S
#{149}5 S
S
:
20 25 30 35 40
GESTATIONAL AGE Cweeks)
Fig 1. Blood P50 in relation to gestational age. Solid
line is calculated linear regression line; r = .62; P < .001.
484 RED CELL OXYGEN AFFINITY
MATERIALS
AND METHODS
In an attempt to reflect fetal conditions as closely
as possible, blood treated with heparin was obtained
from the placental end of the umbilical vein
im-mediately after delivery ofpreterm and term infants
of no known risk, except premature labor. The
gestational age according to maternal history was
always confirmed by physical examination of the
newborn infant.’5 The cord samples used for the
study were obtained from uncomplicated
pregnan-cies that ranged from 24 to 42 weeks of gestation,
which resulted in a newborn who was normal,
non-asphyxiated, and appropriate in weight for
gesta-tional age.’6 All blood analyses were carried out
immediately upon obtaining the samples.
Erythrocyte DPG concentration was determined
on fresh blood treated with heparin according to
the method of Keitt’7 and expressed as micromobes
per gm of Hb and micromoles per ml of RBC. The
reagents were obtained from Sigma Chemical Co,
St Louis. Also, to compare the rebationship between
the intracellular concentration of DPG and pH, the
intracellular and extracellular pH was determined
by a freeze-thaw technique.18
The P50 was determined by gas mixing
tonome-try with a IL blood gas laboratory from
Instrumen-tation Laboratory Inc, Lexington, MA (213 blood
gas analyzer tonometer, 208 gas mixing system, and
182 coximeter). The P50 was expressed in
millime-ters of mercury at a temperature of 37 C a pH 7.40
and Pco2 of 40 mm Hg. In brief, samples for fresh
blood were equilibrated in a tonometer for 30
mm-utes at 37 C with gas mixture containing 40 mm Hg
(± 0.05 mm Hg) of CO2 and varying proportions of
02 and N2. The tonometer permitted an
equiibra-tion and sampling of successive aliquots of blood at
different 02 tensions. Measurement of 02
satura-tion, pH, and Po2 provided the information for
plotting the 02 dissociation curve and P50. The Po2
was always converted to a pH 7.4 by using a Bohr
effect of 0.485’s; the pH values ranged from 7.20 to
7.44, the mean was 7.32
±
0.07. These were at leastfour experimental points for each oxygen
dissocia-tion curve. The standard deviation of the mean P50
obtained on the same blood sample was 0.84 mm
Hg, N = 10. The oxygen saturation obtained by use
of the coximeter was frequently checked by
deter-mining the oxygen contents of blood samples
equi-ibrated in the tonometer by use of the Lex-O2-Con
(Lexington Instruments Corp, Waltham, MA).
The percentage of adult and fetal hemoglobin at
birth was obtained by eluting fetal and adult
he-mogbobin from DEAE Sephadex A50 with a
de-creasing pH gradient (7.8 to 7.1) of Tris-HC1 buffer
by methods previously described.20 The absorbance
of the protein fractions was determined a 280 nm.
Thirty-seven different samples were analyzed for
this study. 2,3-DPG levels and intracellular and
extracellular pH were done on all samples. Because
of the variation in cord blood volume obtained, P50
was determined on 35 and hemoglobin
chromato-graphic separations on 31 of the samples,
respec-tively.
DPG levels were expressed in micromoles per gm
of Hb for correlation purposes and as micromobes
per ml of RBC for comparison with other studies.
When pH values were used for statistical
correla-tion they were converted to hydrogen ion
concen-tration (nEq/biter). Thirteen nonpregnant
babora-tory technicians had their red cell DPG and P50
determined as laboratory controls. The mean values
and SD obtained was 13.53
±
1.07 and 26.4 ± 0.8,respectively. These values correspond well within
the norms that exist in the literature.21’22
RESULTS
The change in P50 as gestational age advances is
shown on Fig 1. There was a significant positive
correlation between P50 and gestational age (r = .6,
P < .001). The regression line increased from 17.8
mm Hg at 24 weeks to 22.5 mm Hg at 42 weeks of
gestation. The P50s, at the mean pH in vivo (7.317
± SD 0.066), increased from 19.5 ± 1.4 mm Hg to
24.7 mm Hg ± 1.8 mm Hg.
The mean percentage of HbA increased with
gestational age (GA). Similarly as previously
re-ported23 the mean value rose from 3% at 24 weeks
to 25% at 42 weeks ofgestation (the linear regression
was percent of HbA = 1.49
x
GA+
37.68; r = .75,P < .001). When P50 and the percentage of HbA is
correlated, there is also a significant positive
cor-relation (r = .7, P < .001; P50 = 0.18 x HbA + 17.9)
as is shown on Fig 2.
The relationship of 2,3-DPG and gestational age
is demonstrated on Fig 3. There was no effect of
26-P5OmmHg
DISCUSSION
S
Fig 2. Blood P50 in relation to adult hemoglobin. Solid
line is the calculated linear regression; r = .67, P < .001.
25.0
S#{149} S
The multiple molecular forms of hemoglobin in
normal man and other animals that has developed
20 0 in the course of evolution appear to confer a
phys-iologic advantage within the period of ontogeny
during which that particular hemoglobin is present
S #{149}5 in high concentration. In people, it is the switch
15.0 from fetal to adult hemoglobin synthesis that has
.:1’________________________
evolved as the main method for the change in red0 #{149}
,
U I U cell oxygen affinity from the fetal to the airenvi-0 10 20 30 40 ronment. The oxygen dissociation curve of normal
IbHbA fetal blood represents the reaction of the
intracel-lular hemoglobin type (HbA and HbF) with oxygen
as modified by the ligands present in the
intracel-lular environment. The ligands of physiologic
im-portance are hydrogen ions and DPG.
The DPG levels obtained in this study are similar
to those reported on in cord blood at delivery by
others24 also the mean P50 of the term infants of
this report (22,1 mm Hg) is similar to the average
value (22.4 mm Hg) of 15 authors listed in a review
article by Nory. A possible explanation of the low
levels of DPG obtained in preterm infants by
oth-ers9”#{176}is that the authors did not consider the
pos-sible effects of adverse clinical conditions which
may occur in preterm deliveries that can induce
extracellular changes in pH which in turn have a
direct effect on the intraerythrocyte environment
and alter DPG synthesis.27 The acid-base instability
of the preterm infants has been observed by several
authors. The greater incidence of complicated
deliveries and asphyxia associated with preterm
_____________________________________ birth compared to term delivery could result in
more preterm infants being acidotic as compared
____________________________________ with term infants.
Fetal blood P50 appears to be mainly a function
of the amount of adult hemoglobin. Yet there is a
________________________________________ scatter of P50 vs percent of HbA which cannot be
gestational age on 2,3-DPG levels. The mean and
SD was 14.86 ± 2.04 mol/gm of Hb. The mean and
SD of the DPG levels when calculated for three
different intervals of fetal development either as
micromoles per gm of Hb or micromoles per ml of
RBC were not significantly different (Table 1).
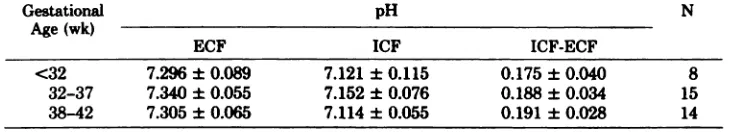
Sim-iarly the pH differences between the fetal red cells
and plasma during the different intervals of
gesta-tion remained stable (Table 2). The mean pH in
vivo was 7.317 ± 0.066 and also within the pH range
of this study the difference between intracellular
and extracellular pH was constant (pH = 0.186 ±
SD 0.032). However, there was
a
significant inversecorrelation between red cell hydrogen ion
concen-trations and DPG levels (r = .5, P < .025; DPG =
-0.31
x
[W]+
38.80).18.0-.0 16fr 110- 100- SD-S . S S S S S S S #{149}#{149}$ S S S
:
55 S 5 S 1 S #{149}S S S S F20 25 30 35 40
GESTATIONAL AGE (weeks)
Fig 3. The 2,3-DPG concentrations as a function of
gestational age. Solid line represents mean; broken line,
± 2 SD.
TABLE 1. Fetal DPG in Reb ation to Gestational Age
Gestational Age (wk) mol/gm of Hb mol/mi of RBC N <32 32-37 38-42
14.27 ± 1.31
15.35 ± 2.94
14.47 ± 1.48
4.617 ± 0.555
5.004 ± 1.005 4.704 ± 0.469
8
15
14
TABLE 2. Extracellular vs Intrac ellubar pH Differences in Relation to Gestati onal Age
Gestational
Age (wk)
pH N
ECF ICF ICF-ECF
<32 7.296 ± 0.089 7.121 ± 0.115 0.175 ± 0.040 8
32-37 7.340 ± 0.055 7.152 ± 0.076 0.188 ± 0.034 15
explained by the data obtained in this study.
How-ever, an explanation that could be possible is that
of cigarette smoking during pregnancy resulting in
differences of fetal carboxyhemoglobin
concentra-tions. This factor may be relevant in view of the
well known effect of carbon monoxide in shifting
the oxyhemogbobin equilibrium to the left. Fetal
concentrations of carboxyhemogbobin are higher
than maternal concentrations in part because of the
higher oxygen affinity of fetal bbood.#{176}
Unfortu-nately the levels of carboxyhemogbobin were not
determined in this study.
During adult life, in a variety of clinical states
where tissue oxygenation is impaired, the DPG
levels rise producing a decrease in red cell oxygen
affinity.3’ However a similar role of 2,3-DPG in
utero may be detrimental to the fetus, if the higher
oxygen affinity of fetal blood favors the placental
transfer of oxygen to the fetus. The observation by
Novy et al32 that fetal anemia does not cause a rise
in 2,3-DPG levels suggests that the ability for fetal
red blood cells to increase 2,3-DPG in utero may
be limited. The human fetus appears to respond to
the stress of threatened hypoxia by both an increase
in the synthesis of fetal hemoglobin and an increase
of its red cell mass.
To date all mammalian fetuses where fetal red
cell organic phosphates were determined at
differ-ent gestational ages showed no changes in 2,3-DPG
levels during fetal life.’24’ Now this report
dem-onstrates that in the human fetus the decrease in
oxygen affinity offetal red cells during last trimester
of gestation is mainly due to the increase in the
amount of adult type hemoglobin. And contrarily
to other studies9”#{176} and to what is often referred to
in review articles,35 fetal red blood cell 2 ,3-DPG
and hydrogen ion concentration during the last
trimester are not a function of fetal development.
In summary, this study has related the oxygen
affinity of fetal red blood cells between 24 to 42
weeks of gestation to the percentage of adult
he-moglobin, the content of 2,3-DPG, and intracellular
pH. There are no changes in 2,3-DPG or pH but
there is a change in the percentage of adult
hemo-globin and a causative relationship is proposed
be-tween the increase of adult hemoglobin and P50.
Also DPG levels are inversely related to
intraceb-lular hydrogen ion concentration which parallels
that of whole blood. This data can be useful to
investigators in evaluating pathologic as well as
therapeutic effects on the concentration of DPG in
the fetal red blood cell.
ACKNOWLEDGMENTS
This research was supported by Grant MA 5120 from
the Medical Research Council of Canada.
We are grateful for the technical assistance provided
by Mr Andrew Bator, BSc, Mrs Fernande Meban#{231}on,
BSc, and Miss Janie Prosmanne, BSc. We are also very
thankful for the cooperation of the delivery room nursing
staff.
REFERENCES
1. Haselhorst G, Stromberger K: Uber den Gasgehalt des Na-belschnurblutes vor und nach der Geburt des Kindes und Uber den Gasaustausch in der Placenta. Z Geburtshilfe Gynaekol 100:48, 1931
2. Bauer CH, Ludwig I, Ludwig M: Different effects of 2,3 diphosphoglycerate and adenosine triphosphate on the oxy-gen affinity of adult and foetal human hemoglobin. Life Sci
7:1339, 1968
3. Tyuma I, Shimizu K: Different response to organic phos-phates ofhuman fetal and adult hemoglobins. Arch Biochem
Biophys 129:404, 1969
4. Bellingham AJ, Detter JC, Lenfant C: Regulatory mecha-nisms of hemoglobin oxygen affinity in acidosis and alkalosis.
J Clin Invest 50:700, 1971
5. Astrup P, Rorth M, Thorshauge L: Dependency on acid-base status of oxyhemoglobin dissociation and 2,3 diphos-phoglycerate levels in human erythrocytes. Scand J Clin Lab Invest 26:47, 1970
6. Wood WG, Weatherall DJ: Haemoglobin synthesis during human foetal development. Nature 244:162, 1973
7. Bard H: Postnatal fetal and adult hemoglobins synthesis in
early preterm newborn infants. J Clin Invest 52:1789, 1975 8. Bard H: The postnatal decline of hemoglobin F synthesis in
normal full-term infants. J Clin Invest 55:395, 1975 9. Delivoria-Papadopoulos M, Roncevic NP, Oski FA:
Postna-tal changes in oxygen transport of term premature, and sick infants: The role ofred cell 2,3 diphosphoglycerate and adult hemoglobin- Pediatr Res 5:235, 1971
10. Orzalesi MM, Hay WW: The regulation of oxygen affinity of fetal blood. I. In vitro experiments and results in normal infants. Pediatrics 48:857, 1971
11. Versmold H, Seifert G, Riegel KP: Blood oxygen affinity in infancy: The interaction of fetal and adult hemoglobin, ox-ygen capacity, and red cell hydrogen ion and 2,3 diphospho-glycerate concentration. Respir Physiol 18:14, 1973 12. Bard H, Fouron JC, Robillard JE, et al: Red cell oxygen
affinity in fetal sheep: The role of 2,3 DPG and adult hemoglobin. JAppl Physiol: Respir Environ Exercise Phys-iol45:7, 1978
13. Bunn HF, Kitchen H: Hemoglobin function in the horse: The role of 2,3 diphosphoglycerate in modifying the oxygen affinity of maternal and fetal blood. Blood 42:471, 1973 14. Bard H, Shapiro M: Perinatal changes of2,3
diphosphoglyc-erate and oxygen affinity in mammals not having fetal type
hemoglobins. Pediatr Res 13:167, 1979
15. Dubowitz LMS, Dubowitz V, Goldberg C: Clinical assess-ment of gestational age in the newborn infant. J Pediatr 77: 1, 1970
16. Lubchenco LO, Hansman C, Boyd E: Intrauterine growth in
length and head circumference as estimated from live births
at gestational ages from 26 to 42 weeks. Pediatrics 37:403, 1966
17. Keitt AS: Reduced nicotinamide adenine dinucleotide-linked analysis of 2,3 diphosphoglyceric acid: Spectrophotometric
and fluorometric procedures. J Lab Clin Med 77:470, 1971
18. Battaglia FC, Behrman RE, Hellegers AE, et al: Intracellular
hydrogen ion concentration changes during acute respiratory acidosis and alcalosis. J Pediatr 66:737, 1965
19. Hilpert P, Fleischmann RG, Kempe D, et al: The Bohr effect related to blood and erythrocyte pH. Am J Physiol 205:337,
1963
20. Dozy AM, Kleihauer EF, Huisman HJ: Studies on the het-erogeneity of hemoglobin. XIII. Chromatography of various
human and animal hemoglobin types of DEAE-Sephadex. J Chromatogr 32:723, 1968
O-Hb dissociation analyse and mixing technique. J AppI
Physiol 30:903, 1971
22. Rand PW, Norton JM, Barker N, et al: Influence of athletic training on hemoglobin-oxygen affinity. Am J Physiol 224: 1334, 1973
23. Bard H, Makowski EL, Meschia G, et al: The relative rates ofsynthesis of hemoglobins A and F in immature red cells of newborn infants. Pediatrics 45:766, 1970
24. Weiss RR, Roginsky MS, Mann LI, et al: ErythrOCyte 2,3-diphosphoglycerate in normal and hypertensive gravid
women and their newborn infants. Am J Obstet Gynecol 124:
692, 1976
25. Novy MJ: Fetal oxygenation and its relation to maternal
and fetal blood oxygen affinity, inPreservation ofRed Blood Cells. Washington, National Academy of Sciences, 1973, pp
. 101-117
26. Desforges JF, Slawsky P: Red cell 2, 3-diphosphoglycerate and intracellular arterial pH in acidosis and alkalosis. Blood
40:740, 1972
27. Rapoport S: The regulation of glycolysis in mammalian erythrocytes. Biochemistry 4:69, 1968
28. Kildeberg P: Disturbances ofhydrogen ion balance occurring in premature infants. I. Early types of acidosis. Acta
Pae-diatr Scand 53:505, 1964
29. Malan AF, Evans A, De V Heese H: Serial acid-base deter-minations in normal premature and full-term infants during the first 72 hours of life. Arch Dis Child 4th645, 1965
30. Longo LD: The biological effects of carbon monoxide on the
pregnant woman, fetus, and newborn infant. Am J Obstet Gynecol 129:69, 1977
31. Bunn HF, Forget GB, Ranney HM: Human Hemoglobins.
Philadelphia, WB Saunders Co, 1977, pp 79-93
32. Novy MJ, Frigoletto FD, Bastardy CHL, et al: Changes in
umbilical-cord blood oxygen affinity after intrauterine
trans-fusions for erythroblastosis. N Engi JMed 285:589, 1971 33. Bard H: The effect of placental insufficiency on fetal and
adult hemoglobin synthesis. Am J Obstet Gynecol 120:67,
1974
34, Baumann R, Teischel F, Zoch R, et al: Changes in red cell,
2,3-diphosphoglycerate concentration as cause of the post-natal decrease of pig blood oxygen affinity. Respir Physiol
19:153, 1973
35. Wood WG, Clegg JB, Weatherall DJ: Development biology
of human hemoglobins, in EB Brown (ed): Progress in
Hematology. New York, Grune & Stratton, 1977, vol 10,
p43
EVALUATING
THE ROLE OF X-RAY
DIAGNOSIS
. . . the use of x-rays in medical diagnosis must be subject to the following
interrelated criteria:
C The technical and medical unit (ie, staff, equipment, etc) should be efficient’, otherwise
the results would be of no value.
. There should be demonstrable diagnostic or prognostic efficacy (called efficacy-i);
otherwise the x-ray examination would be a waste of resources and, as an unnecessary
radiation exposure, a risk to health.
C Under non-academic conditions, there should be therapeutic efficacy (efflcacy-2)-ie,
the x-ray result should influence the proposed management and treatment of the
patient. Particularly where resources for health services are limited, as in developing
countries, this level ofefficacy should be an indispensible condition ofthe examination.
. Ideally, the improvement of individual health (efficacy-3) should, in the majority of
cases, follow efficacy-2. (Unfortunately, this will not always be the case; hence the
distinction between these two criteria.)
. The ultimate objective, the improvement of public health (efficacy-4), should be the
essential criterion for the planning of health services in general, and for appropriately
integrated radiologic services in particular.
These considerations are not only generally applicable; they form the basis
for assessing the role (ie, the efficacy and efficiency) of any health management
procedure. A risk-benefit or cost-effectiveness evaluation depends upon these
criteria, which are particularly relevant to medical measures carrying a certain
risk, however small (as in radiation), and to those that are relatively expensive.
Submitted by Student