1535-9778/08/$08.00⫹0 doi:10.1128/EC.00446-07
Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Key Function for the CCAAT-Binding Factor Php4 To Regulate Gene
Expression in Response to Iron Deficiency in Fission Yeast
䌤
†
Alexandre Mercier,
1Stephen Watt,
2Ju
¨rg Ba
¨hler,
2and Simon Labbe
´
1*
De´partement de Biochimie, Faculte´ de Me´decine, Universite´ de Sherbrooke, Sherbrooke, Quebec J1H 5N4, Canada,1and
Fission Yeast Functional Genomics Group, Wellcome Trust Sanger Institute, Hinxton, Cambridge, CB10 1HH, United Kingdom2
Received 8 December 2007/Accepted 18 January 2008
The fission yeastSchizosaccharomyces pomberesponds to the deprivation of iron by inducing the expression
of thephp4ⴙgene, which encodes a negative regulatory subunit of the heteromeric CCAAT-binding factor. Once
formed, the Php2/3/4/5 transcription complex is required to inactivate a subset of genes encoding iron-using
proteins. Here, we used a pan-S. pombemicroarray to study the transcriptional response to iron starvation and
identified 86 genes that exhibitphp4ⴙ-dependent changes on a genome-wide scale. One of these genes encodes
the iron-responsive transcriptional repressor Fep1, whose mRNA levels were decreased after treatment with
the permeant iron chelator 2,2ⴕ-dipyridyl. In addition, several genes encoding the components of
iron-depen-dent biochemical pathways, including the tricarboxylic acid cycle, mitochondrial respiration, amino acid biosynthesis, and oxidative stress defense, were downregulated in response to iron deficiency. Furthermore, Php4 repressed transcription when brought to a promoter using a yeast DNA-binding domain, and iron deprivation was required for this repression. On the other hand, Php4 was constitutively active when gluta-thione levels were depleted within the cell. Based on these and previous results, we propose that iron-dependent inactivation of Php4 is regulated at two distinct levels: first, at the transcriptional level by the iron-responsive GATA factor Fep1 and second, at the posttranscriptional level by a mechanism yet to be identified, which inhibits Php4-mediated repressive function when iron is abundant.
Iron is an essential trace element for most organisms (22). Due to its ability to easily gain and lose electrons, iron serves as a catalytic cofactor for a wide variety of enzymes involved in fundamental metabolic processes, including the tricarboxylic acid (TCA) cycle, respiration, DNA synthesis and repair, and the biosynthesis of metabolites (13). Although abundant in nature, iron is generally found in forms that are biologically unavailable. To counter the difficulties of acquiring sufficient iron for cellular function, organisms have developed diverse strategies to scavenge iron from the environment or reallocate it within the cell, while avoiding the buildup of concentrations that could lead to cellular toxicity.
InSchizosaccharomyces pombe, when the intracellular iron
level is raised, the protein Fep1 represses transcription of iron uptake genes (26, 47, 48). Based on DNA microarray analysis, it has been suggested that Fep1 is also required for the iron-regulated expression of genes encoding proteins involved in iron-sulfur cluster formation, compartmentalization, and utili-zation of inorganic iron (53). Fep1 is a member of the GATA factor family, which is comprised of transcription factors that contain either one or two Cys2/Cys2-type zinc finger motifs that recognize and bind to GATA-containing DNA sequences (56). Fep1 orthologs are found in many yeast genomes, including
Ustilago maydis (Urbs1) (2, 3), Neurospora crassa (Sre) (18,
66),Aspergillus nidulans(SreA) (17, 43, 44),Candida albicans
(Sfu1) (28, 49), andCryptococcus neoformans(Cir1) (21), but not inSaccharomyces cerevisiae(47). Instead, inS. cerevisiae, the genes required for high-affinity iron uptake are activated by the iron-responsive transcription factors Aft1 and Aft2, for which there are no orthologs in the majority of other fungi, except those that belong to the sensu stricto group of the genus
Saccharomyces(9, 10, 54, 64).
When facing iron starvation, organisms possess strategies to upregulate genes encoding the components of the iron trans-port machinery. They also induce an alternative pathway to inactivate expression of genes encoding nonessential iron-binding proteins, allowing a redistribution of intracellular iron stores. InEscherichia coli, a small noncoding regulatory RNA, denoted RyhB, is induced under conditions of iron starvation (32). It hybridizes with its target mRNAs encoding iron-con-taining enzymes and mediates their degradation through an RNase E-dependent process (31). In contrast, under iron-plete conditions, the repressor Fur binds to the promoter re-gion of RyhB and shuts down its transcription (32). A similar mechanism has been reported for the action of two small RNAs, termed PrrF1 and PrrF2, in the control of iron-regu-latable genes inPseudomonas aeruginosa(62).
InS. cerevisiae, the RNA-binding protein Cth2 is synthesized
when the availability of iron is limited (52). The induction of
CTH2expression requires functionalAFT1/AFT2genes. Cth2 harbors two CX8CX5CX3H-type zinc finger motifs that bind to
AU-rich RNA elements in the 3⬘untranslated regions (UTR) of a large number of transcripts (52). Many of the transcripts encode iron-containing cellular components, including en-zymes involved in the TCA cycle, heme and Fe-S cluster bio-synthesis, and the electron transport chain. In response to iron starvation, Cth2 specifically binds its mRNA targets and
trig-* Corresponding author. Mailing address: De´partement de Biochimie, Faculte´ de Me´decine, Universite´ de Sherbrooke, 3001, 12e Ave. Nord, Sherbrooke, Quebec J1H 5N4, Canada. Phone: (819) 820-6868, ext. 15460. Fax: (819) 564-5340. E-mail: [email protected].
† Supplemental material for this article may be found at http://ec .asm.org/.
䌤Published ahead of print on 25 January 2008.
493
on September 8, 2020 by guest
http://ec.asm.org/
gers their degradation. In contrast, in the presence of iron,
CTH2expression is strongly downregulated, allowing replen-ishment of its target mRNAs. In S. cerevisiae, when iron is scarce, a second mechanism operates at the transcriptional level (58). It implicates theGLT1 gene that encodes a Fe-S cluster-containing enzyme, named glutamate synthase (Glt1). Iron limitation-dependent downregulation ofGLT1expression is independent of Aft1 (58). The repression ofGLT1 in re-sponse to iron deprivation occurs via acis-acting palindrome sequence, CGGN15CCG, found within its promoter region. Mutation of both half-sites of thiscis-acting element results in a loss of iron-dependent regulation ofGLT1expression. Cur-rently, it is unclear whichtrans-acting factor regulates the iron-dependent transcription ofGLT1(58).
In a previous study, we identified three S. pombe genes (pcl1⫹, sdh4⫹, and isa1⫹) encoding putative iron-containing proteins that are downregulated in response to iron deficiency (39). This regulation occurs at the transcriptional level and requires the conserved core sequence, 5⬘-CCAAT-3⬘. Consis-tent with the base pair composition of the regulatory sequence,
the S. pombe CCAAT-binding factor, which consists of four
subunits, Php2, Php3, Php4, and Php5, plays a key role in controlling the levels ofpcl1⫹,sdh4⫹, andisa1⫹expression in response to iron deprivation (39). When heterologously ex-pressed inS. cerevisiae, theS. pombe php2⫹,php3⫹, andphp5⫹
genes functionally complement theS. cerevisiae hap2⌬,hap3⌬, andhap5⌬mutants, respectively (37, 46, 63). InS. cerevisiae, Hap2, Hap3, and Hap5 are constitutively expressed and re-quired for the formation of a DNA-binding complex (36, 38). Hap4 is responsible for the transcriptional activation capability of the multimeric complex and is not required for the DNA-binding activity (15).HAP4gene expression is repressed when cells are grown in the presence of glucose, while it is induced in media containing a nonfermentable carbon source. As a consequence, the positive transcriptional activity of the CCAAT-binding factor is regulated mainly by the biosynthesis of the activation subunit Hap4. Until recently, a Hap4 homolog had not been identified inS. pombe. However, a recent com-putational search using the completeS. pombegenome infor-mation revealed one open reading frame (SPBC16E9.01c or
php4⫹) that was predicted to encode a putative Hap4 homolog (61). AlthoughS. pombe Php4 harbors a limited overall se-quence homology withS. cerevisiae Hap4, it contains a con-served domain that is present in Hap4. In S. cerevisiae, this domain is known to be required for the association of Hap4 with Hap2/Hap3/Hap5 (36). By monitoring steady-state levels ofpcl1⫹,sdh4⫹, andisa1⫹transcripts, we determined that the inactivation of php4⫹ results in sustained levels of pcl1⫹,
sdh4⫹, andisa1⫹gene expression without any change in re-sponse to iron starvation, suggesting that Php4 might act as a negative regulatory subunit of the CCAAT-binding factor. Consistent with a role for Php4 as a regulatory protein under low-iron-supply conditions, its expression is turned off under conditions of iron repletion by the iron-dependent GATA-type transcriptional repressor Fep1 (39). Consequently, php4⫹ is expressed only during conditions of iron deprivation.
Genomic profiling in C. albicans has revealed that the
HAP43 gene, which encodes a putativeC. albicans Php4
or-tholog, is upregulated under iron-deficient conditions (28). In contrast, when environmental iron is abundant, analogous to
Php4 fromS. pombe, the expression ofHAP43is repressed in an Sfu1-dependent manner (28). InA. nidulans, it has been elegantly shown that in the presence of iron, the GATA-type transcription factor SreA downregulates the expression of the
HAPX-encoded CCAAT-binding subunit of the Hap complex (19). Conversely, under iron-limiting conditions,HAPXis in-duced, and like S. pombe Php4, the HapX subunit interacts with the CCAAT-binding core complex, which is constituted of HapB, HapC, and HapE (19). Once assembled, the hetero-meric transcription factor represses genes that encode iron-using proteins, as well as theSREAgene (19). Furthermore, using a bimolecular fluorescence complementation assay, it has been shown that the interaction of HapX with the HapB/C/E heterotrimer occurs only during iron deficiency, suggesting the existence of an additional control mechanism of the HapX activity besides its transcriptional control by SreA (19).
To gain further insight into the role of Php4 in preventing futile expression of genes encoding iron-rich enzymes when iron is limiting, we examined global gene expression by DNA microarray analysis. First, we sought to identify genes whose transcription is expressed at lower levels during iron deficiency. Second, we identified genes whose expression is upregulated in aphp4⌬disruption strain when iron is scarce. These two com-bined data sets identified 86 genes that are regulated in a Php4-dependent manner, including the gene encoding the iron-dependent GATA repressor Fep1. Using a one-hybrid approach, we showed that a constitutively expressed Gal4-Php4 fusion protein is able to repress transcription in response to iron deficiency when brought to a DNA promoter. In con-trast, this Php4-mediated repression was annihilated when iron is replete, revealing an iron-sensing ability for Php4. On the other hand, we observed that Php4 is constitutively active when GSH levels are depleted in agcs1⌬strain. Taken together, the results reveal that Php4 is a central transcriptional repressor in fission yeast that is required during iron deficiency for the downregulation of a large set of genes encoding iron-using proteins.
MATERIALS AND METHODS
Yeast strains and growth conditions.TheS. pombestrains used in this study were the wild-type FY435 (h⫹his7-366 leu1-32 ura4-⌬18 ade6-M210) (8) and two isogenic mutant strains, php4⌬ (h⫹ his7-366 leu1-32 ura4-⌬18 ade6-M210 php4⌬::Kanr) andfep1⌬(h⫹his7-366 leu1-32 ura4-⌬18 ade6-M210 fep1⌬::ura4⫹)
(39, 47). All three strains were cultured in yeast extract medium (YE) containing 0.5% yeast extract and 3% glucose that was further supplemented with 225 mg/liter of adenine, histidine, leucine, uracil, and lysine (1). To maintain strains bearing plasmids, synthetic Edinburgh minimal medium with necessary auxotro-phic requirements was used (1). Liquid cultures were seeded to anA600of 0.5 and grown to exponential phase (A600of⬃1.0) and then treated with 250M 2,2⬘-dipyridyl (Dip) or 100M FeCl3for 90 min.
RNA isolation, sample labeling, and microarray hybridization.After the iron deprivation (Dip) or iron repletion (FeCl3) of yeast strains, 5 optical density units of cells (⬃1⫻108cells/ml) was harvested by centrifugation and snap-frozen in liquid nitrogen. Total RNA was extracted by the hot phenol method as described previously (11). RNAs were quantified spectrophotometrically, and then 20g of RNA was labeled by directly incorporating Cy3- and Cy5-dCTP using Superscript reverse transcriptase (Invitrogen, Carlsbad, CA) as described previously (29). The resulting cDNA was hybridized onto glass DNA microarrays containing probes for 99.3% of all known and predictedS. pombegenes (http://www.sanger .ac.uk/PostGenomics/S_pombe/). Microarrays were scanned using a GenePix 4000B laser scanner (Axon Instruments, Foster City, CA). Data were subse-quently analyzed with GenePix Pro software. Unreliable signals were filtered out, and data were normalized using a customized Per1 script (29). The script applies cutoff criteria to discard data from weak signals. Genes that did not give
on September 8, 2020 by guest
http://ec.asm.org/
ducible results between the biological repeats were eliminated. Furthermore, genes with 50% of their data points missing were also discarded. Data acquisi-tion, processing, and normalization were further analyzed using GeneSpring GX software (Agilent Technologies, Cheshire, United Kingdom). Normalized signals were exported from GeneSpring into Microsoft Excel.
Experimental design.We adopted an experimental design composed of two nodes: wild-type iron deprived (⫺Fe)7wild-type iron replete (⫹Fe) and wild-type (⫺Fe)7php4⌬(⫺Fe). The corresponding time courses were performed as three independent biological repeats. Two of them were used in the microarray protocol for which the Cy dyes were swapped. The third independent biological repeat was used for quantification of mRNAs using the RNase protection pro-tocol. Following microarray data acquisition and within-array normalization, gene values from the wild type (⫹Fe) andphp4⌬(⫺Fe) were divided by the corresponding value of wild type (⫺Fe), set as the reference sample. The ex-pression ratios of biological repeat experiments were averaged. Genes were classified asphp4⫹-dependent if their expression changed 1.5-fold more than the average of two repeats during iron repletion versus iron deprivation and if they were upregulated 1.5-fold more than the average of two repeats in thephp4⌬ strain (AMY15) during iron starvation compared with the wild-type strain under the same conditions. Gene annotations were taken from the GeneDB at the Sanger Institute (http://www.genedb.org/genedb/pombe/index.jsp). Fine-map-ping studies performed in different laboratories have revealed that the highly conserved five core nucleotides CCAAT are almost invariably conserved to modulate the binding with high affinity of the yeast CCAAT-binding complex (30). Because of the almost absolute requirement for each of the CCAAT nucleotides, this was the pentanucleotide that we used. Additional flanking nucleotides on the 5⬘side and on the 3⬘side of the CCAAT consensus sequence might be required for preferential binding, but we have not considered such (preferred) sequences when we searched for putative CCAAT elements in the promoter regions of genes encoding iron-using proteins (see Table 2).
RNase protection analysis.Total RNA was extracted using the hot phenol method as described above for the microarray experiments. RNAs were quan-tified spectrophotometrically. RNA from each sample was used in the RNase protection protocol as described previously (6), with the following modifications. A 15-g sample of RNA was hybridized with the indicated riboprobes (Table 1) for 15 h at 55°C in a 30-l reaction mixture containing 1⫻hybridization buffer (40 mM PIPES [piperazine-N,N⬘-bis(2-ethanesulfonic acid); pH 6.4], 1 mM EDTA, 400 mM NaCl, 50% formamide). Subsequent to hybridization, 250l of RNase digestion buffer (50 mM sodium acetate [pH 4.4], 400 mM NaCl, 10 mM EDTA) containing 15 units of RNase T1 (Calbiochem, San Diego, CA) per ml was added, and the reaction mixtures were kept at 30°C for 90 min. After incubation, each RNA sample was precipitated with a solution containing 265l of 4.0 M guanidine thiocyanate, 530l of isopropanol, and 10l of 10 mg/ml tRNA. The precipitation was allowed to proceed for 30 min at⫺20°C. RNA pellets were washed twice with 70% ethanol and resuspended in 10l of form-amide loading buffer. Five microliters of each sample was loaded per lane on a 6% polyacrylamide-8 M urea gel and subjected to electrophoresis. The gels were
dried and exposed to a PhosphorImage screen, and the radioactive bands were quantified by using PhosphorImager SP and ImageQuant software (Molecular Dynamics, Sunnyvale, CA). All riboprobes used in this study are listed in Table 1. DNA templates for antisense riboprobes were cloned into the BamHI and EcoRI sites of pBluescript SK(⫹) (Stratagene, La Jolla, CA). The resultant constructs were linearized with BamHI for subsequent labeling with [␣-32P]UTP and the T7 RNA polymerase.
One-hybrid system.The reporter plasmid pUR4xGal4-lacZwas created in four steps. First, we introduced an SphI-BamHI PCR-amplified DNA fragment containing 367 bp of the 5⬘-noncoding region and the first 10 codons of the zym1⫹gene (27) into the SphI-BamHI-cut Yep357R vector (41). Second, the resulting plasmid, designated Yep357Rzym1⫹-367lacZ, was subsequently di-gested with SpeI (the SpeI restriction site was previously introduced in one primer used for the PCR amplification and placed immediately after the SphI site) and Bsu36I. This latter restriction site is located within thelacZgene. Subsequently, the purified SpeI-Bsu36Izym1⫹-367lacZDNA fragment was ex-changed with a corresponding restriction DNA fragment into the plasmid pSP1fio1⫹-1155lacZ(47), generating the plasmid pSP1zym1⫹-367lacZ. Third, two high-performance liquid chromatography-purified complementary oligonu-cleotides were annealed pairwise to form a partial double-stranded DNA. This product was made completely double-stranded by incubation with the Klenow fragment of DNA polymerase I in the presence of deoxyribonucleotides. The resulting double-stranded DNA molecule containing four consensus Gal4-bind-ing sites (5⬘-CGGN11CCG-3⬘) (59) was then amplified by PCR. Because the primers contained NotI and SpeI restriction sites, the purified PCR-amplified fragment was digested with these enzymes and inserted immediately upstream of the minimalzym1⫹promoter gene fused tolacZin pSP1zym1⫹-367lacZ. Fourth, the resulting construct, designated pSP14xGal4-lacZ, was digested with NotI, filled-in with Klenow, and digested with HindIII. The purified DNA fragment was cloned via NotI (Klenow) and HindIII sites into the SmaI and HindIII sites of pUR18 (5), generating pUR4xGal4-lacZ.
To construct the pJK-194promphp4⫹plasmid, a 194-bp SacII-XbaI PCR-amplified DNA segment of thephp4⫹promoter was inserted into the same sites of pJK148 (23). Subsequently, the two iron-responsive GATA sequences (posi-tions⫺188 to⫺183 and⫺165 to⫺160) found in thephp4⫹promoter (39) were mutated, thereby rendering its expression unregulated by iron or iron starvation. Precisely, the oligonucleotide 5⬘-GCAACACCGCGGTAGATCGCCGTCAAT TTAATCGTATCTCTCTGTCCGATTAAAACAAACCACCAACG-3⬘ was used in combination with another oligonucleotide (5⬘-CTAGTCTAGAGAGCA AAAAAAAAGTTGGGGGGATAGG-3⬘) to generate by PCR aphp4⫹ pro-moter DNA fragment containing multiple point mutations in GATA elements (underlined letters represent nucleotide substitutions). The modifiedphp4⫹ pro-moter was used to replace the wild-type propro-moter fragment in plasmid pJK-194promphp4⫹to generate pJK-194*promphp4⫹. The open reading frame en-coding the Gal4 DNA-binding domain (DBD) (positions 1 to 147) was PCR amplified from the pAS1 plasmid (14) and then inserted into the BamHI and SalI restriction sites of pJK-194*promphp4⫹to create pJK-194*promphp4⫹-GAL4. Thephp4⫹allele was amplified by PCR using primers designed to generate SalI and Asp718 sites at the upstream and downstream termini of the open reading frame. Once generated, the DNA fragment was inserted in frame to the C terminus of the Gal4 DBD. The resulting plasmid, pJK-194*promphp4⫹- GAL4-php4⫹, and two control plasmids, designated pJK-194*promphp4⫹-GAL4and pJK-194*promphp4⫹-GAL4-GFP, were subsequently linearized with NruI for integration in aphp4⌬disruption strain. Purified integrants were transformed with pUR4xGal4-lacZ. Liquid cultures of purified cotransformants were seeded to anA600of 0.5 and then grown to exponential phase (A600of 1.0). At this log phase, cultures were treated with 250M Dip or 100M FeCl3, or left un-treated. Isolated RNA samples from cells that were treated or untreated were analyzed by RNase protection assays. Whole-cell extracts were also prepared from aliquots of cultures used for steady-state mRNA analyses. Extract prepa-ration and protein detection were conducted as described previously (39). West-ern blot analysis was performed with monoclonal anti-Gal4 antibody RK5C1 (Santa Cruz Biotechnology, Santa Cruz, CA), monoclonal anti-PCNA antibody PC10 (Sigma, St. Louis, MO), and horseradish peroxidase-conjugated secondary antibodies (Amersham Biosciences, Arlington Heights, IL) and developed with enhanced chemiluminescent detection reagents.
Fluorescence microscopy.S. pombestrain FY435 was transformed with pSP1-GFP, pSP1-SV40-GFP, or pJK-194*promphp4⫹-GAL4-GFP. The first two mids were constructed by a strategy described previously (7). To generate plas-mid pJK-194*promphp4⫹-GAL4-GFP, thegfpgene was isolated by PCR using primers corresponding to the start and stop codons of the open reading frame from the pSF-GP1 plasmid (25). To clone the PCR product, thegfpgene was obtained using primers that contained SalI and Asp718 restriction sites. The
TABLE 1. Target genes monitored by RNase protection assays
Gene identifier Gene name Riboprobe length (bp)
Reference or source
SPBC32H8.12c act1⫹ 151 39
SPAC26F1.14c aif1⫹ 190 This study
SPCC1235.02 bio2⫹ 175 This study
SPCC757.07c ctt1⫹ 170 This study
SPCC191.07 cyc1⫹ 170 This study
SPBC29A3.18 cyt1⫹ 182 This study
SPAC13A11.02c erg11⫹ 191 This study
SPAC24B11.13 hem3⫹ 182 This study
SPCC645.03c isa1⫹ 188 39
SPAC9E9.03 leu2⫹ 181 This study
SPBC16E9.01C php4⫹ 182 39
SPAC140.01 sdh2⫹ 186 This study
SPCC330.12c sdh3⫹ 170 This study
SPBP23A10.16 sdh4⫹ 178 39
SPAC10F6.01C 192 This study
SPAC20G8.04C 192 This study
SPAC24C9.06C 194 This study
SPAPB1E7.07 196 This study
on September 8, 2020 by guest
http://ec.asm.org/
purified fragment was digested with these enzymes and subsequently inserted into the pJK-194*promphp4⫹-GAL4plasmid. Cells were grown to logarithmic phase and viewed by direct fluorescence microscopy as described previously (50). An Eclipse E800 epifluorescent microscope (Nikon, Melville, NY) equipped with an Orca ER digital cooled camera (Hamamatsu, Bridgewater, NJ) was used to obtain the fluorescence and differential interference contrast images of the cells. Microscopy analysis of the samples was realized using a magnification of⫻1,000 with the following filters: 465 to 495 nm (green fluorescent protein [GFP]) and 340 to 380 nm (DAPI [4⬘,6⬘-diamidino-2-phenylindole]). The cell fields shown in this study (see Fig. S3B in the supplemental material) are representative of three independent experiments.
RESULTS
Effects of iron status and Php4 on theS. pombe
transcrip-tome.Our previous work revealed that threeS. pombemRNAs
(pcl1⫹,sdh4⫹, and isa1⫹) encoding proteins that function in iron-dependent pathways are downregulated in response to iron deprivation (39). We found that this repression ofpcl1⫹,
sdh4⫹, and isa1⫹is mediated by the CCAAT-binding factor Php4. To determine whether additional genes are differentially regulated as a function of iron availability, we applied a mi-croarray approach based on the following conditions. First, microarrays were hybridized with probes derived from RNA isolated from iron-replete versus iron-depleted wild-type cells. Therefore, genes with higher expression levels in the presence of iron (or lower expression levels in the presence of the iron chelator Dip) were detected. Second, microarrays were also hybridized with probes generated from RNA purified from iron-depleted php4⌬ mutant cells versus iron-depleted wild-type cells. Therefore, genes with higher expression levels in
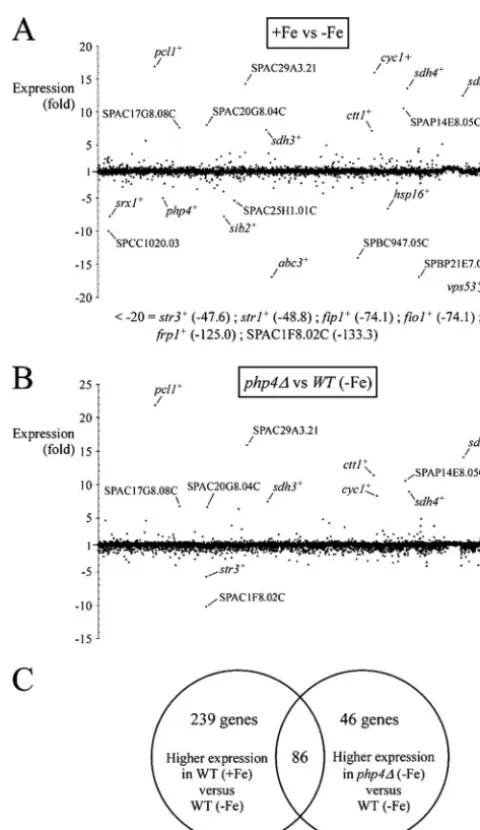
php4mutants (in iron-poor medium) were identified. The first set of array results revealed that 325 genes were expressed at higher levels in iron-replete cells (averaging⬎1.5-fold), includ-ingpcl1⫹,cyc1⫹, and sdh4⫹, as well as novel putative genes (Fig. 1; see Table S1 in the supplemental material). On the other hand, 251 genes were highly expressed in iron-depleted cells (see Table S2 in the supplemental material). As one would expect, some of these genes, such asfrp1⫹,fip1⫹,fio1⫹,
str1⫹,str2⫹, andstr3⫹, were involved in the uptake of iron (Fig. 1; see Table S2 in the supplemental material) (26). Further-more, we also noted significant changes in the transcriptional profiles of other genes, including SPAC1F8.02c (uncharacter-ized ORF), SPAC3A12.15 (a putative subunit of the vesicular GARP complex), SPBC359.05 (a putative vacuolar ABC trans-porter), and SPBC947.05c (a putative ferrireductase) (see Ta-ble S2 in the supplemental material). The second set of array results showed that 132 genes were expressed at higher levels in iron-limitedphp4⌬mutant cells (averaging⬎1.5-fold) (Fig. 1; see Table S3 in the supplemental material). These potential Php4 targets included partially or fully characterized genes, pcl1⫹,
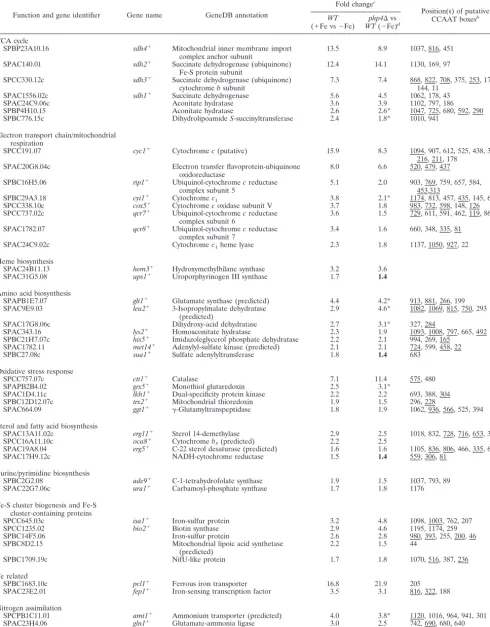
sdh4⫹, andisa1⫹(39), as well as novel uncharacterized genes (see Table S3 in the supplemental material). Overall, 86 genes were expressed at higher levels in both data sets, including genes en-coding the TCA cycle proteins Sdh1 to Sdh4, Aco1, and Kgd2; the electron transport chain enzymes Cyc1, Rip1, Cyt1, Cox5, and Qcr7/8; the amino acid biosynthesis enzymes glutamate synthase, Lys2, and His5; and the Fe-S cluster biogenesis protein Isa1 (Ta-ble 2). Accordingly, almost all of these genes have one or more copies of the CCAAT consensus sequence within their promoters (Table 2). Taken together, these results identified many tran-scripts that exhibit iron-dependent expression patterns. Several
genes exhibited decreased abundance in iron-starved wild-type cells and are potentially under the control of the CCAAT-binding factor Php4.
FIG. 1. Genome-wide picture of theS. pombe response to iron availability and the global effects ofphp4⌬deletion relative to wild-type cells during iron deficiency. (A) Differentially expressed genes in iron-replete wild-type cells (⫹Fe) (100M FeCl3) versus iron-limited wild-type cells (⫺Fe) (250M Dip). In total, 4,944 genes (xaxis) were analyzed by DNA microarrays. All differentially expressed genes are shown, but only 20 of them are labeled on the graph. Lists of genes differentially expressed as a function of iron availability are in Tables S1 and S2 in the supplemental material. Six genes (str3⫹,str1⫹,fip1⫹,
fio1⫹,frp1⫹, and SPAC1F8.02c) were downregulated more than 20-fold in wild-type cells grown in the presence of iron. (B) Differentially expressed genes in wild-type (WT) versusphp4⌬mutant cells grown in iron-limiting medium. As depicted in panel A, in total, 4,944 genes (x
axis) were analyzed by DNA microarrays. All differentially expressed genes are depicted, but only 12 of them are labeled on the graph. A list of regulated genes is in Table S3 in the supplemental material. (C) Venn diagram representing the mRNA expression profile of wild-type cells grown under iron-replete conditions compared to cells grown under conditions of iron deprivation and the mRNA expression profile of wild-type cells grown under low iron conditions compared tophp4⌬
mutant cells grown under the same conditions.
on September 8, 2020 by guest
http://ec.asm.org/
TABLE 2. Genes derepressed by both Fe repletion and aphp4⌬deletion
Function and gene identifier Gene name GeneDB annotation
Fold changec
Position(s) of putative CCAAT boxesb
WT (⫹Fe vs⫺Fe)
php4⌬vs WT(⫺Fe)d
TCA cycle
SPBP23A10.16 sdh4⫹ Mitochondrial inner membrane import
complex anchor subunit
13.5 8.9 1037, 816, 451
SPAC140.01 sdh2⫹ Succinate dehydrogenase (ubiquinone)
Fe-S protein subunit
12.4 14.1 1130, 169, 97
SPCC330.12c sdh3⫹ Succinate dehydrogenase (ubiquinone)
cytochromebsubunit
7.3 7.4 868, 822, 708, 375, 253, 175, 144, 11
SPAC1556.02c sdh1⫹ Succinate dehydrogenase 5.6 4.5 1062, 178, 43
SPAC24C9.06c Aconitate hydratase 3.6 3.9 1102, 797, 186
SPBP4H10.15 Aconitate hydratase 2.6 2.6* 1047, 725, 680, 592, 290
SPBC776.15c DihydrolipoamideS-succinyltransferase 2.4 1.8* 1010, 941
Electron transport chain/mitochondrial respiration
SPCC191.07 cyc1⫹ Cytochromec(putative) 15.9 8.3 1094, 907, 612, 525, 438, 380,
216, 211, 178
SPAC20G8.04c Electron transfer flavoprotein-ubiquinone
oxidoreductase
8.0 6.6 520, 479, 437
SPBC16H5.06 rip1⫹ Ubiquinol-cytochromecreductase
complex subunit 5
5.1 2.0 903, 769, 759, 657, 584, 453,313
SPBC29A3.18 cyt1⫹ Cytochromec1 3.8 2.1* 1174, 813, 457, 435, 145, 65
SPCC338.10c cox5⫹ Cytochromecoxidase subunit V 3.7 1.8 983, 732, 598, 148, 126
SPCC737.02c qcr7⫹ Ubiquinol-cytochromecreductase
complex subunit 6
3.6 1.5 729, 611, 591, 462, 119, 86
SPAC1782.07 qcr8⫹ Ubiquinol-cytochromecreductase
complex subunit 7
3.4 1.6 660, 348, 335, 81
SPAC24C9.02c Cytochromec1heme lyase 2.3 1.8 1137, 1050, 927, 22
Heme biosynthesis
SPAC24B11.13 hem3⫹ Hydroxymethylbilane synthase 3.2 3.6
SPAC31G5.08 ups1⫹ Uroporphyrinogen III synthase 1.7 1.4
Amino acid biosynthesis
SPAPB1E7.07 glt1⫹ Glutamate synthase (predicted) 4.4 4.2* 913, 881, 266, 199
SPAC9E9.03 leu2⫹ 3-Isopropylmalate dehydratase
(predicted)
2.9 4.6* 1082, 1069, 815, 750, 293
SPAC17G8.06c Dihydroxy-acid dehydratase 2.7 3.1* 327, 284
SPAC343.16 lys2⫹ Homoaconitate hydratase 2.3 1.9 1093, 1008, 797, 665, 492
SPBC21H7.07c his5⫹ Imidazoleglycerol phosphate dehydratase 2.2 2.1 994, 269, 165
SPAC1782.11 met14⫹ Adenylyl-sulfate kinase (predicted) 2.1 2.1 724, 599, 458, 22
SPBC27.08c sua1⫹ Sulfate adenylyltransferase 1.8 1.4 683
Oxidative stress response
SPCC757.07c ctt1⫹ Catalase 7.1 11.4 575, 480
SPAPB2B4.02 grx5⫹ Monothiol glutaredoxin 2.5 3.1*
SPAC1D4.11c lkh1⫹ Dual-specificity protein kinase 2.2 2.2 693, 388, 304
SPBC12D12.07c trx2⫹ Mitochondrial thioredoxin 1.9 1.5 296, 228
SPAC664.09 ggt1⫹ ␥-Glutamyltranspeptidase 1.8 1.9 1062, 936, 566, 525, 394
Sterol and fatty acid biosynthesis
SPAC13A11.02c erg11⫹ Sterol 14-demethylase 2.9 2.5 1018, 832, 728, 716, 653, 375
SPCC16A11.10c oca8⫹ Cytochromeb5(predicted) 2.2 2.5
SPAC19A8.04 erg5⫹ C-22 sterol desaturase (predicted) 1.6 1.6 1105, 836, 806, 466, 335, 64
SPAC17H9.12c NADH-cytochrome reductase 1.5 1.4 559, 306, 81
Purine/pyrimidine biosynthesis
SPBC2G2.08 ade9⫹ C-1-tetrahydrofolate synthase 1.9 1.5 1037, 793, 89
SPAC22G7.06c ura1⫹ Carbamoyl-phosphate synthase 1.7 1.8 1176
Fe-S cluster biogenesis and Fe-S cluster-containing proteins
SPCC645.03c isa1⫹ Iron-sulfur protein 3.2 4.8 1098, 1003, 762, 207
SPCC1235.02 bio2⫹ Biotin synthase 2.9 4.6 1195, 1174, 259
SPBC14F5.06 Iron-sulfur protein 2.6 2.8 980, 393, 255, 200, 46
SPBC8D2.15 Mitochondrial lipoic acid synthetase
(predicted)
2.2 1.5 44
SPBC1709.19c NifU-like protein 1.7 1.8 1070, 516, 387, 236
Fe related
SPBC1683.10c pcl1⫹ Ferrous iron transporter 16.8 21.9 205
SPAC23E2.01 fep1⫹ Iron-sensing transcription factor 3.5 3.1 816, 322, 188
Nitrogen assimilation
SPCPB1C11.01 amt1⫹ Ammonium transporter (predicted) 4.0 3.8* 1120, 1016, 964, 941, 301
SPAC23H4.06 gln1⫹ Glutamate-ammonia ligase 3.0 2.5 742, 690, 680, 640
SPAC10F6.01c Sulfite reductase beta subunit (predicted) 2.7 2.5 677
Continued on following page
on September 8, 2020 by guest
http://ec.asm.org/
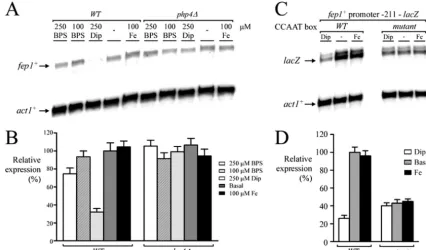
Repression of fep1ⴙ transcript upon addition of the iron
chelator Dip.DNA microarrays indicated that the steady-state
levels of fep1⫹ mRNA in iron-treated wild-type cells were increased 3.5-fold above the levels observed in cells treated
with Dip (Table 2). Likewise, in thephp4⌬mutant cells,fep1⫹
mRNA levels increased (3.1-fold) after 90 min of Dip treat-ment (Table 2). This result was surprising because our previous studies, using the iron chelator bathophenanthrolene
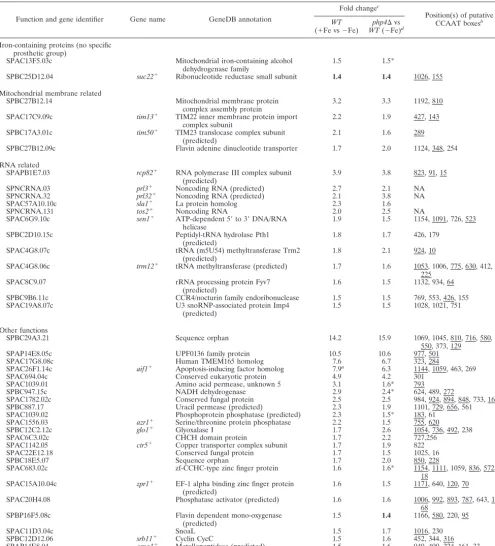
disul-TABLE 2—Continued
Function and gene identifier Gene name GeneDB annotation
Fold changec
Position(s) of putative CCAAT boxesb
WT (⫹Fe vs⫺Fe)
php4⌬vs WT(⫺Fe)d
Iron-containing proteins (no specific prosthetic group)
SPAC13F5.03c Mitochondrial iron-containing alcohol
dehydrogenase family
1.5 1.5*
SPBC25D12.04 suc22⫹ Ribonucleotide reductase small subunit 1.4 1.4 1026, 155
Mitochondrial membrane related
SPBC27B12.14 Mitochondrial membrane protein
complex assembly protein
3.2 3.3 1192, 810
SPAC17C9.09c tim13⫹ TIM22 inner membrane protein import complex subunit
2.2 1.9 427, 143
SPBC17A3.01c tim50⫹ TIM23 translocase complex subunit (predicted)
2.1 1.6 289
SPBC27B12.09c Flavin adenine dinucleotide transporter 1.7 2.0 1124, 348, 254
RNA related
SPAPB1E7.03 rcp82⫹ RNA polymerase III complex subunit
(predicted)
3.9 3.8 823, 91, 15
SPNCRNA.03 prl3⫹ Noncoding RNA (predicted) 2.7 2.1 NA
SPNCRNA.32 prl32⫹ Noncoding RNA (predicted) 2.1 3.8 NA
SPAC57A10.10c sla1⫹ La protein homolog 2.3 1.6
SPNCRNA.131 tos2⫹ Noncoding RNA 2.0 2.5 NA
SPAC6G9.10c sen1⫹ ATP-dependent 5⬘to 3⬘DNA/RNA
helicase
1.9 1.5 1154, 1091, 726, 523
SPBC2D10.15c Peptidyl-tRNA hydrolase Pth1
(predicted)
1.8 1.7 426, 179
SPAC4G8.07c tRNA (m5U54) methyltransferase Trm2
(predicted)
1.8 2.1 924, 10
SPAC4G8.06c trm12⫹ tRNA methyltransferase (predicted) 1.7 1.6 1053, 1006, 775, 630, 412,
225
SPAC8C9.07 rRNA processing protein Fyv7
(predicted)
1.6 1.5 1132, 934, 64
SPBC9B6.11c CCR4/nocturin family endoribonuclease 1.5 1.5 769, 553, 426, 155
SPAC19A8.07c U3 snoRNP-associated protein Imp4
(predicted)
1.5 1.5 1028, 1021, 751
Other functions
SPBC29A3.21 Sequence orphan 14.2 15.9 1069, 1045, 810, 716, 580,
550, 373, 129
SPAP14E8.05c UPF0136 family protein 10.5 10.6 977, 501
SPAC17G8.08c Human TMEM165 homolog 7.6 6.7 323, 284
SPAC26F1.14c aif1⫹ Apoptosis-inducing factor homolog 7.9a
6.3 1144, 1059, 463, 269
SPAC694.04c Conserved eukaryotic protein 4.9 4.2 301
SPAC1039.01 Amino acid permease, unknown 5 3.1 1.6* 793
SPBC947.15c NADH dehydrogenase 2.9 2.4* 624, 489, 272
SPAC1782.02c Conserved fungal protein 2.5 2.5 984, 924, 894, 848, 733, 160
SPBC887.17 Uracil permease (predicted) 2.3 1.9 1101, 729, 656, 561
SPAC1039.02 Phosphoprotein phosphatase (predicted) 2.3 1.5* 183, 61
SPAC1556.03 azr1⫹ Serine/threonine protein phosphatase 2.2 1.5 755, 620
SPBC12C2.12c glo1⫹ Glyoxalase I 1.7 2.6 1054, 736, 492, 238
SPAC6C3.02c CHCH domain protein 1.7 2.2 727,256
SPAC1142.05 ctr5⫹ Copper transporter complex subunit 1.7 1.9 822
SPAC22E12.18 Conserved fungal protein 1.7 1.5 1025, 16
SPBC18E5.07 Sequence orphan 1.7 2.0 850, 228
SPAC683.02c zf-CCHC-type zinc finger protein 1.6 1.6* 1154, 1111, 1059, 836, 572,
18 SPAC15A10.04c zpr1⫹ EF-1 alpha binding zinc finger protein
(predicted)
1.6 1.5 1171, 640, 120, 70
SPAC20H4.08 Phosphatase activator (predicted) 1.6 1.6 1006, 992, 893, 787, 643, 123,
68
SPBP16F5.08c Flavin dependent mono-oxygenase
(predicted)
1.5 1.4 1166, 580, 220, 95
SPAC11D3.04c SnoaL 1.5 1.7 1016, 230
SPBC12D12.06 srb11⫹ Cyclin CycC 1.5 1.6 452, 344, 316
SPAP14E8.04 oma1⫹ Metallopeptidase (predicted) 1.5 1.6 940, 400, 224, 161, 33
a
Determined by RNase protection assays.
b
NA, not applicable. Underlining, CCAAT boxes in reverse orientation (position relative to the 5⬘C).
c⫹
Fe, iron replete;⫺Fe, iron depleted. Boldface, value below the threshold but associated with enzymes that need or potentially need iron for their activity.
d
*, value originates from only one biological repeat (second value not available or aberrant).
on September 8, 2020 by guest
http://ec.asm.org/
fonic acid (BPS) (100M), found no apparent effect onfep1⫹
transcript levels by iron availability (47). To clarify the situa-tion, we reanalyzedfep1⫹gene expression. The wild-type strain was grown in the presence of iron and two distinct iron chela-tors at different concentrations, and the steady-state levels of
fep1⫹mRNA were assayed by RNase protection experiments. As we found previously (47), the nonpermeant iron chelator BPS had no significant effect onfep1⫹transcript levels when cells were grown in medium containing 100M BPS (Fig. 2A and B). Increasing the concentration of BPS to 250M only slightly affected the fep1⫹ transcript levels. However, upon treatment with 250M Dip (a permeant iron chelator), the steady-state levels of fep1⫹ mRNA were significantly de-creased 3.3-fold under basal levels (Fig. 2A and B). This re-duction in fep1⫹ mRNA levels parallels those observed by DNA microarrays (3.5-fold) (Table 2). In the presence of ex-ogenous iron (100M),fep1⫹mRNA was expressed with an efficacy similar to that observed under basal conditions (Fig. 2A and B). Using an isogenic strain harboring an insertionally inactivatedphp4allele, we determined whether Php4 plays an essential role infep1⫹downregulation in the presence of 250
M Dip. As shown in Fig. 2A, cells harboring an inactivated
php4⫹ gene (php4⌬) failed to repress fep1⫹expression after treatment with 250 M Dip. We therefore searched for a CCAAT consensus sequence upstream offep1⫹. Interestingly, we found three putative CCAAT-type regulatory sequences (positions⫺184 to⫺188,⫺322 to⫺326, and⫺816 to⫺820) that might serve to controlfep1⫹expression in response to iron
deficiency. To test whether some of these elements could con-trol fep1⫹ expression as a function of iron availability, we inserted multiple point mutations that foster changes known to abolish the binding of the CCAAT-binding factor to CCAAT boxes. We used a DNA segment containing the fep1⫹locus starting at⫺211 from the translational initiator codon up to
⫹30 that was fused tolacZ. Mutation of the base pairs within the ⫺188CCAAT⫺184element (AACCG instead of CCAAT)
fostered a lack of iron responsiveness in thefep1⫹gene (Fig. 2C and D). In contrast, a wild-type copy of thefep1⫹promoter gene showed an iron limitation-dependent downregulation of its expression (Fig. 2C and D). Taken together, these results reveal that, upon treatment with a permeant iron chelator, the CCAAT-binding factor Php4 and thefep1⫹promoter region located between positions ⫺184 and ⫺188 are required for iron starvation-mediated repression offep1⫹expression.
Exposure of cells to low concentrations of iron triggers downregulation of genes encoding enzymes of the TCA cycle.
The TCA cycle is a sequence of reactions in mitochondria that oxidizes the acetyl moiety of acetyl-CoA and reduces coen-zymes that are reoxidized through the electron transport chain, linked to the formation of ATP. In addition to acting as a “metabolic furnace,” some intermediates of the TCA cycle (e.g., citrate,␣-ketoglutarate, succinyl-CoA, and oxaloacetate) serve as precursors for anabolic processes, including the bio-synthesis of fatty acids, amino acids, heme, and glucose (35). The eight enzymes of the classic TCA cycle are encoded by 15 genes inS. pombe. Four of the enzymes, termed citrate
syn-FIG. 2. Dip-dependent downregulation offep1⫹transcript levels requires Php4 and a pentanucleotide sequence, CCAAT. (A) Logarithmic phase cultures of isogenic strains FY435 (wild type [WT]) and AMY15 (php4⌬) were untreated (-) or incubated in the presence of BPS (100 and 250M), Dip (250M), or FeCl3(Fe) (100M). Fifteen-milliliter samples were taken after 90 min of treatment. RNA was prepared from each sample and analyzed by RNase protection assays. mRNA steady-state levels offep1⫹andact1⫹(indicated with arrows) were analyzed with respect to thephp4⫹ allele status. (B) Quantification of three independent RNase protection assays, including the experiment shown in panel A. (C) Wild-type and⫺188CCAAT⫺184mutantfep1⫹promoter fragments were assayed by RNase protection assay. Total RNA was isolated from
untreated cultures (-) or cultures treated with Dip (250M) or FeCl3(100M). ThelacZandact1⫹ mRNA levels are shown with arrows. (D) Quantification of three independent RNase protection assays, including the experiment shown in panel C.
on September 8, 2020 by guest
http://ec.asm.org/
thase, aconitase, fumarase, and malate dehydrogenase, are encoded by single genes: SPAC6C3.04, SPAC24C9.06c, SPCC18.18c, and SPCC306.08c, respectively. The other four enzymes are constituted of subunits encoded by distinct genes: SPAC11G7.03 and SPBC902.05c for the NAD⫹-dependent isocitrate dehydrogenase; SPBC3H7.03, SPBC776.15c, and SPAC1002.09c for the ␣-ketoglutarate dehydrogenase; SPAC16E8.17c and SPCC1620.08 for the succinyl-CoA ligase; and SPAC1556.02c, SPAC140.01, SPCC330.12c, and SPBP23A10.16 for the succinate dehydrogenase (Sdh1 to Sdh4). Our gene expression profiling data suggest that at least six loci encoding components of enzymes of the TCA cycle were downregulated in response to iron deprivation (Table 2). To assess whether our microarray results were successful in identifying iron-regulated genes, we carried out RNase protec-tion analysis (using an independent biological repeat) to ex-amine the relative expression of four genes encoding compo-nents that function in the TCA cycle. In the presence of the iron chelator Dip, sdh2⫹, sdh3⫹, sdh4⫹, and aco1⫹ mRNA levels were strongly repressed, by 12.6-, 7.9-, 11.9-, and 14.7-fold, respectively, compared with their basal levels of expres-sion observed in untreated cells (Fig. 3). Under iron-replete conditions, transcript levels from sdh2⫹, sdh3⫹, sdh4⫹, and
aco1⫹remained approximately equal to those observed in un-treated cells (Fig. 3). To further investigate the relation be-tween the Php4 protein and the expression ofsdh2⫹, sdh3⫹,
sdh4⫹, and aco1⫹, we deleted php4⫹ (php4⌬). php4⌬ cells showed sustained levels ofsdh2⫹, sdh3⫹, sdh4⫹, andaco1⫹
expression without any strong change in response to iron de-privation (Fig. 3). Consistently, sdh2⫹, sdh3⫹, sdh4⫹, and
aco1⫹also contain CCAAT sequences within their promoters (Table 2). Taken together, these data indicate that the repres-sion of S. pombe sdh1, sdh2, sdh3, and sdh4;aco1; and the SPBC776.15c-encoded e2 component of␣-ketoglutarate
dehy-drogenase occurs through the activity of the CCAAT-binding factor Php4 that represses transcription from these loci in response to iron deficiency.
Genes encoding components involved in the mitochondrial electron transport chain are repressed under conditions of
iron deprivation.The microarray data further suggested that
additional components of iron-dependent metabolic pathways were differentially expressed as a function of iron availability (Table 2). In mammals and obligate aerobic yeasts, such as
Yarrowia lipolytica,Neurospora crassa, andDebaryomyces
han-senii, components of the respiratory chain are contained in
four large iron-rich protein complexes embedded in the inner mitochondrial membrane. Of these complexes, the last three are conserved in budding and fission yeasts, whereas complex I is absent from these organisms (24, 45). Instead, these yeasts have specialized NADH-Q oxidoreductases that are distinct yet able to oxidize mitochondrial NADH to NAD (57). When comparing the mRNA expression profile of wild-typeS. pombe
cells grown under iron-replete conditions to cells cultured un-der low iron conditions, we observed that key components of the respiratory chain are responsive to changing iron concen-trations. Transcript levels of SPAC20G8.04c, cyt1⫹, rip1⫹,
qcr7⫹, andqcr8⫹that potentially encode constituents of com-plex III decreased in low iron conditions (Table 2). Likewise, the expression of cytochromec-related constituents (cyc1⫹and
cox5) that play a role in the flow of electrons through com-plexes III and IV are also modulated by high/low iron concen-trations (Table 2). To independently verify the array data, RNase protection assays were performed on a subgroup of genes encoding putative or partially characterized components of the respiratory chain. As shown in Fig. 4, SPAC20G8.04c,
cyt1⫹, and cyc1⫹ transcript levels were downregulated 6.6-, 6.0-, and 10.6-fold, respectively, under conditions of iron star-vation in a wild-type strain. In thephp4⌬mutant strain, the
FIG. 3. RNase protection analysis of four transcripts encoding components of the TCA cycle. (A) Total RNA was prepared from the wild-type (WT) strain and itsphp4⌬mutant derivative grown to logarithmic phase. When indicated, cultures were untreated (-) or treated with either Dip (D) (250M) or FeCl3(Fe) (100M) for 90 min. Fifteen-microgram RNA samples were probed, and RNA loads were normalized to the level of transcripts of theact1⫹gene. The results shown are representative of three independent experiments. (B) Histograms show the normalized expression levels of the indicated mRNAs. Values are the averages of triplicate measurements⫾standard deviations.
on September 8, 2020 by guest
http://ec.asm.org/
SPAC20G8.04c gene was unregulated in response to iron de-privation, exhibiting a steady-state level of mRNA similar to that observed under basal or iron-replete conditions (Fig. 4). Surprisingly, however, we found that in the presence of Dip (250M),cyt1⫹andcyc1⫹transcript levels inphp4⌬cells were still slightly downregulated 2.1- and 1.6-fold, respectively, un-der iron-limiting conditions (Fig. 4). A gene regulated solely by Php4, like SPAC20G8.04c, would not be expected to show decreased expression inphp4⌬cells in response to iron-limit-ing conditions. However, genes such ascyt1⫹andcyc1⫹may require Php4 for full repression (compare wild-type versus
php4⌬cells under iron-limiting conditions) (Fig. 4), but other factors must be involved as well.
Iron deprivation negatively affects expression of genes en-coding proteins involved in biotin-, iron sulfur cluster-, and
amino acid-biosynthetic pathways.The transcriptional profiles
of genes encoding several iron sulfur cluster enzymes impli-cated in biotin, iron sulfur center, and amino acid biogenesis were influenced by the cellular iron status (Table 2). Among them, the biotin synthase (Bio2), the iron sulfur assembly pro-tein 1 (Isa1), a putative glutamate synthase (SPAPB1E7.07), and the 3-isopropylmalate dehydratase (Leu2) were repressed 2.9-, 3.2-, 4.4-, and 2.9-fold, respectively, under conditions of iron deficiency (Table 2). To determine whether our microar-ray approach was successful in identifying transcripts that are low in iron-poor medium, we measured bio2⫹, isa1⫹, SPAPB1E7.07, and leu2⫹ steady-state mRNA levels in un-treated or un-treated cells with either iron or the iron chelator Dip. As shown in Fig. S1 in the supplemental material, under conditions of low iron,bio2⫹,isa1⫹, SPAPB1E7.07, andleu2⫹
expression was decreased 3.1-, 3.5-, 14.7-, and 3.9-fold, respec-tively, compared with the basal levels observed in the wild-type strain. As one would expect, using either DNA microarray or RNase protection analysis,bio2⫹,isa1⫹, andleu2⫹transcripts
exhibit similar iron starvation-dependent changes (compare Table 2 to Fig. S1 in the supplemental material), except for SPAPB1E7.07, for which we obtained a more pronounced repression of its steady-state mRNA level by RNase protection assay (14.7-fold) compared to that observed by DNA microar-ray analysis (4.4-fold). To investigate the role of Php4 in the regulation ofbio2⫹,isa1⫹, SPAPB1E7.07, andleu2⫹mRNAs under iron deprivation, we measured the mRNA downregula-tion of these transcripts in the absence of Php4 (see Fig. S1 in the supplemental material,WTversusphp4⌬mutant). In the
php4⌬ mutant strain, the levels of these four mRNAs were virtually unregulated by iron or iron starvation. Furthermore, we observed that the corresponding genes harbor one or more potential CCAAT sequences in their promoters (Table 2). Collectively, these data show that Php4 functions in the down-regulation of these mRNAs in response to iron deprivation.
Limiting iron reduces the expression of several genes
en-coding iron-containing proteins.In addition to the
above-men-tioned genes, microarray experiments indicated that, under conditions of iron starvation, other genes encoding iron-con-taining proteins involved in diverse cellular functions, includ-ing ergosterol biosynthesis (erg11⫹), apoptosis induction (aif1⫹), oxidative stress defense (ctt1⫹), heme synthesis (hem3⫹), and sulfate assimilation (SPAC10F6.01c), were also downregulated (Table 2). To verify these data, we analyzed these transcript levels by RNase protection analyses using un-treated, iron-starved, or iron-replete cells (see Fig. S2 in the supplemental material). Using wild-type cells, our data showed downregulation of erg11⫹ (3.7-fold), aif1⫹ (7.9-fold), ctt1⫹
(3.9-fold), hem3⫹ (3.8-fold), and SPAC10F6.01c (3.9-fold) transcripts in response to iron deprivation (Fig. S2 in the sup-plemental material). In contrast, under both standard (un-treated) and iron-replete conditions, transcript levels for these genes were readily detectable, being more abundant compared
FIG. 4. Effect ofphp4⌬mutation on the expression of three genes encoding proteins involved in the mitochondrial electron transport chain. (A) Wild-type (WT) orphp4⌬cells were incubated in the absence (-) or presence of Dip (D) (250M) or FeCl3(Fe) (100M) for 90 min. After total RNA extraction, the SPAC20G8.04c,cyt1⫹,cyc1⫹, andact1⫹steady-state mRNA levels were analyzed by RNase protection assays. Results shown are representative of three independent experiments. (B) Quantification of SPAC20G8.04c,cyt1⫹, andcyc1⫹levels after the treatments shown in panel A. The values are the means of triplicate determinations⫾standard deviations.
on September 8, 2020 by guest
http://ec.asm.org/
with levels of mRNAs observed under iron-limiting conditions in the presence of Dip (Fig. S2 in the supplemental material). To test whether Php4 was required for iron starvation-depen-dent repression, we analyzed php4⌬ cells, which showed sustained levels of erg11⫹, aif1⫹, ctt1⫹, hem3⫹, and SPAC10F6.01c gene expression without substantial changes in response to iron starvation (Fig. S2 in the supplemental mate-rial). Furthermore, the promoter regions of these genes, except for hem3⫹, contain putative consensus CCAAT sequences, suggesting a direct role for Php4 in downregulating their ex-pression in response to iron-limiting conditions.
Gal4-Php4 fusion protein is a transcriptional repressor.
Given our previous results (39) and based on the data scribed so far, we propose that, under conditions of iron de-privation, Php4 associates with the Php2/Php3/Php5 heterotri-mer and acts as a transcriptional repressor. According to this model, Php4 would provide the repression function, while the CCAAT-binding Php2/Php3/Php5 heterotrimer would serve to recruit Php4 to the promoter of target genes. One critical point of this model is the ability of Php4 to act as a repressor when brought to the DNA at a promoter. To test this hypothesis, we fused the DBD of an S. cerevisiae protein, Gal4, to the N terminus of the intact Php4 protein. To test whether insertion of the Gal4 DBD interfered with Php4 function, the tagged
(GAL4-php4⫹) and untagged (php4⫹) coding sequences were
separately integrated into aphp4⌬strain, and integrants were analyzed for their ability to repress isa1⫹ mRNA levels in response to iron-limiting conditions. As shown in Fig. 5, when
Gal4-Php4 was expressed in aphp4⌬ mutant strain, it func-tionally restored iron limitation-dependent downregulation of
isa1⫹expression in a manner similar to that of the wild-type Php4 protein. In the absence of Php4,isa1⫹mRNA levels were found to be constitutive and unresponsive to cellular iron sta-tus (Fig. 5).
To investigate the mechanism whereby Php4 functions to regulate gene expression, four consensusGAL4binding sites were introduced upstream of a minimal promoter-lacZ re-porter gene to provide binding sites for the Gal4-Php4 fusion protein.php4⌬mutant cells were cotransformed with both the reporter gene construct and integrative expression plasmids, making intact the Gal4 DBD, Gal4-GFP, or the Gal4-Php4 protein. The results shown in Fig. 6A reveal that Gal4-Php4 represses the reporter gene in the presence of the iron chelator Dip (4.2-fold). In contrast, in the absence of the iron chelator (untreated cells) or in the presence of iron (100M FeCl3), the Gal4-Php4 fusion protein shows no repression (Fig. 6A and B). Likewise, the Gal4 DBD alone and Gal4-GFP failed to mediate downregulation ofisa1⫹expression under both iron-limiting and iron-replete conditions (Fig. 6A and B). To verify that the Gal4 DBD alone, Gal4-GFP, or the Gal4-Php4 pro-tein was present inphp4⌬cells, total protein extracts from cells transformed with integrative plasmids expressing the indicated Gal4 derivative molecules were analyzed by immunoblotting. As shown in Fig. S3A in the supplemental material, the loss of repression in response to iron was not due to the lack of protein expression, because all the fusion proteins tested in the one-hybrid assay were synthesized as confirmed by immuno-blot analyses. For simplicity, a single control lane of the Gal4 DBD alone (predicted molecular mass of 16.8 kDa) is shown (see Fig. S3A in the supplemental material). As an additional control, we also verified that the nuclear localization signal of Gal4 was recognized by an S. pombe importin  receptor, thereby allowing its transport in the nucleus (see Fig. S3B in the supplemental material).
While the one-hybrid approach revealed that Php2/Php3/ Php5 subunits do not have to be the DBD for repressional activity of Php4 when iron is limiting, one interpretation is that Php4 may still interact with Php5 during iron deficiency to mediate transcriptional repression when brought to the DNA by the Gal4 DBD. To investigate this possibility, we performed the one-hybrid experiment with aphp4⌬php5⌬double mutant strain. As shown in Fig. 6C and D, in the presence of the iron chelator Dip, we still detected a clear repression of the re-porter gene (3.9-fold). Conversely, in the absence of Dip (un-treated cells) or in the presence of iron, the Gal4-Php4 fusion protein fostered no repression. Taken together, the results show that Php4 alone can act as a transcriptional repressor when brought to the DNA by the Gal4 DBD.
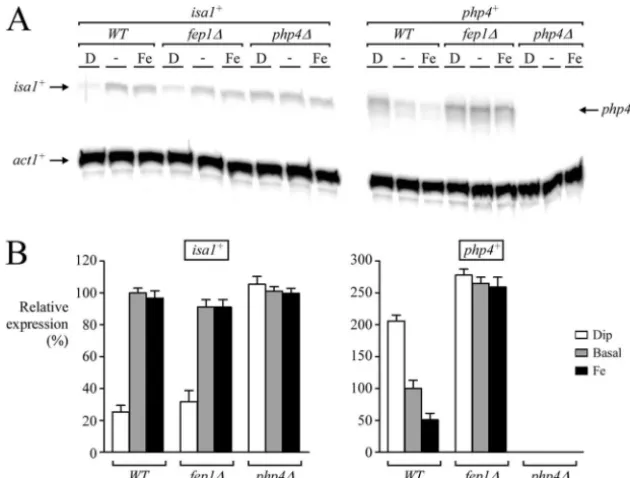
Php4-mediated regulation occurs infep1⌬strain cells.We
have previously demonstrated that Fep1 interacts with the
php4⫹promoter GATA elements in an iron-dependent man-ner, repressingphp4⫹gene expression in response to iron (39). On the other hand, under low-iron-supply conditions, Fep1 cannot bind to DNA and fails to repressphp4⫹gene expres-sion (39). In the previous section, we described the use of an integrative plasmid expressing the Gal4-Php4 fusion protein under the control of thephp4⫹promoter in which the Fep1 binding sites,⫺188AGATAT⫺183and⫺165TGATAA⫺160, have FIG. 5. The Gal4-Php4 fusion protein is functional. (A) The tagged
Php4 protein was expressed into aphp4⌬mutant strain and tested for its ability to downregulate theisa1⫹gene expression in response to iron-limiting conditions.php4⌬cells harboring the integrative plasmids pJK148 (vector alone), pJK148php4⫹, and pJK148GAL4-php4⫹were incubated in the absence (-) or presence of Dip (D) (250M) or FeCl3 (Fe) (100M) for 90 min at 30°C. Total RNA was extracted and then used in the RNase protection protocol to determineisa1⫹andact1⫹
mRNA levels. Wild-type strain FY435 (WT) was used as a control. (B) Quantification of three independent RNase protection assays, in-cluding the experiment shown in panel A.
on September 8, 2020 by guest
http://ec.asm.org/
been mutated, therefore exhibiting no iron-dependent changes in its expression (see Fig. S3A in the supplemental material). Surprisingly, when GAL4-php4⫹expression is not under the control of Fep1, iron could still ensure Gal4-Php4 inactivation. As shown in Fig. 6A and B, no repression of the reporter gene was observed in the presence of iron. In contrast, a strong repression did occur under conditions of iron starvation. This suggests that in the absence of Fep1-mediated regulation an additional mechanism exists to ensure an iron-dependent in-activation of Gal4-Php4. Given this observation, we used a
fep1⌬mutant strain and examined the transcriptional profile of
isa1⫹, a Php4-dependent target gene. To ascertain if in the absence of Fep1,isa1⫹mRNA levels are still expressed under basal and iron-replete conditions, we carried out RNase pro-tection assays with wild-type andfep1⌬cells. As shown in Fig. 7, isa1⫹ transcript levels were clearly detectable from both wild-type andfep1⌬cells in the presence of iron. In contrast, in wild-type andfep1⌬cells treated with Dip,isa1⫹steady-state mRNA levels were decreased 4.0- and 2.9-fold, respectively, compared with the basal levels observed in untreated cells. Accordingly, isa1⫹ mRNA was constitutively expressed and unresponsive to iron deprivation in aphp4⌬strain. As previ-ously shown (39),php4⫹mRNA levels are unresponsive to iron for repression infep1⌬cells and show constitutive expression. As a control, thephp4⫹transcript was absent in php4⌬cells (Fig. 7). Collectively, these results suggest that in the absence
of Fep1, iron can still trigger the inactivation of the Php4 protein through an additional mechanism that has yet to be established.
Php4 is unable to sense iron during GSH deprivation.It is
known that theS. cerevisiaetranscription factor Aft1 is acti-vated by iron deficiency and inactiacti-vated by iron repletion (65). Although the iron-dependent mechanism by which Aft1 is in-activated remains unclear, it has been shown that the presence of glutathione (GSH) is important for iron-mediated inactiva-tion of Aft1 funcinactiva-tion (55). In fact, in GSH-deficient cells, theS.
cerevisiaeiron regulon genes are constitutively upregulated and
are less responsive to iron for repression (55). Like Aft1,S.
pombe Php4 is active during iron deficiency, except that it
represses (instead of activates) transcription. Because an S.
pombe gcs1⌬ disruption strain exhibits GSH auxotrophy (4),
we tested the effect of this strain with respect to the ability of Php4 to mediate repression ofisa1⫹expression. As shown in Fig. 8,gcs1⌬cells exhibited lowisa1⫹mRNA levels that were unaffected by the presence of iron or Dip. In contrast, in wild-type cells,isa1⫹transcript levels were repressed only in the presence of Dip (2.6-fold). This is consistent with the fact that the repression of isa1⫹ by Php4 in response to iron is unlikely, because php4⫹ transcription is downregulated by Fep1 when sufficient iron is present in wild-type cells (39). Thus, the fact thatisa1⫹transcripts were strongly repressed in
gcs1⌬ cells (even in the presence of iron) could suggest that
FIG. 6. The Gal4-Php4 fusion protein represses transcription under conditions of iron deficiency. (A)S. pombecells bearing aphp4⌬deletion were cotransformed with pJK148GAL4DBD (Gal4 DBD alone) plus pUR4xGAL4-lacZ, pJK148GAL4-GFP plus pUR4xGAL4-lacZ, or pJK148GAL4-Php4plus pUR4xGAL4-lacZ.php4⌬cells containing the indicated plasmids were grown to anA600of⬃1.0. At this optical density, Dip (D) (250M) or FeCl3(Fe) (0 and 100M) was added, and cultures were incubated for 90 min. Total RNA was prepared from culture aliquots.lacZandact1⫹mRNA levels (arrows) were detected using RNase protection assays. The data illustrated are representative of three independent experiments. As a negative control, the wild-type strain was transformed with pUR4xGAL4-lacZalone. (B) The histogram represents the average repression levels mediated by the Gal4-Php4 fusion protein. (C) php4⌬ php5⌬ double mutant cells were cotransformed with pJK148GAL4DBD(Gal4 DBD alone) plus pUR4xGAL4-lacZor pJK148GAL4-Php4plus pUR4xGAL4-lacZ.php4⌬php5⌬cells harboring the indicated plasmids were treated as described in the legend for panel A. Total RNA was prepared and then analyzed by RNase protection assays.
lacZandact1⫹mRNA levels are indicated with arrows. (D) Quantification oflacZtranscript levels after treatments. The values are the averages
of triplicate determinations⫾standard deviations.
on September 8, 2020 by guest
http://ec.asm.org/
php4⫹expression failed to respond to iron. To investigate this point, we monitoredphp4⫹mRNA levels ingcs1⌬cells. We found thatphp4⫹expression was no more repressed by iron in the absence of GSH (Fig. 8). Because the expression ofphp4⫹
is normally negatively regulated by iron and this regulated expression requires a functional Fep1, we analyzed the mRNA levels offio1⫹, a well-known Fep1 target (47). We found that, likephp4⫹mRNA,fio1⫹mRNA was constitutively expressed in the absence of GSH, confirming the inability of Fep1 to repress its transcript steady-state levels in response to iron. In contrast, under iron-replete conditions,fio1⫹expression was found to be reduced 15.5-fold compared with the basal level of thefio1⫹transcript detected in untreated wild-type cells (Fig. 8). Because fep1⫹ is itself a Php4 target gene and Php4 is constitutively active in gcs1⌬ cells, the inactivation of Fep1 could be explained by a sustained Php4-mediated repression of
fep1⫹ transcript levels during GSH deficiency. This can be ascertained by monitoringfep1⫹mRNA steady-state levels in the absence of GSH. As shown in Fig. 8, when GSH was depleted,fep1⫹mRNA was repressed irrespective of iron sta-tus. Its steady-state transcript levels were much lower than that observed in untreated wild-type cells (gcs1⫹). Taken together, these results reveal that the disruption of GSH biosynthesis leads to a constitutive activation of Php4, rendering genes under its control continually downregulated regardless of the cellular iron status.
DISCUSSION
Work in a number of laboratories has demonstrated the existence of novel cellular mechanisms that redistribute the
limited iron available during iron deficiency. In bacteria such
asE. coli,Pseudomonas aeruginosa, andVibrio cholerae, as well
asS. cerevisiae, these mechanisms operate mainly at the
post-transcriptional level (33, 40, 52). In the above-mentioned bac-teria, the mechanism implicates small antisense RNAs that trigger the degradation of mRNAs (33). In budding yeast, the RNA-binding protein Cth2, which is a member of the tristet-raprolin family of proteins, targets specific mRNA molecules for degradation (52). InS. pombe, transcriptional regulation of gene expression plays a key role in the iron-sparing response (39). We have demonstrated that, during iron deficiency, the CCAAT-binding factor subunit Php4 is responsible for the transcriptional repression of three genes related to iron utili-zation (39). In this study, we ascertain through a genome-wide microarray analysis that Php4 is a key component capable of coordinating the downregulation of multiple genes in response to iron starvation. Among these genes, several encode proteins involved in iron-dependent metabolic pathways, such as the TCA cycle, mitochondrial respiration, oxidative stress defense, and heme biosynthesis. We also found two genes encoding proteins with iron-related functions: pcl1⫹and fep1⫹. While the first one was expected because it was previously identified as a Php4 target gene (39), the iron limitation-dependent downregulation offep1⫹gene expression was unexpected. Our previous studies, using the nonpermeant iron chelator BPS, found that the steady-state levels offep1⫹mRNA are consti-tutive and unresponsive to cellular iron status (47). In contrast, in this study, when iron was limited by the addition of the permeant iron chelator Dip, gene expression profiles indicated
fep1⫹ gene expression decreased 3.5-fold. If the differential
FIG. 7. Inactivation of Php4 can be uncoupled from iron-mediated Fep1-dependent extinction ofphp4⫹transcript levels. (A) Wild-type (WT),
fep1⌬, andphp4⌬cells were grown to logarithmic phase. Cultures were divided for their respective treatments (untreated [-], 250M Dip [D], or 100M FeCl3[Fe]) and grown for 90 min. After total RNA extraction, theisa1⫹andphp4⫹steady-state mRNA levels were analyzed by the RNase protection protocol with actin (act1⫹) as an internal control. Results shown are representative of three independent experiments. (B) Quantifi-cation ofisa1⫹andphp4⫹mRNA levels after treatments shown in panel A. The values are the averages of triplicate determinations⫾standard deviations.
on September 8, 2020 by guest
http://ec.asm.org/
effects of Dip and BPS chelators are due to the permeability of Dip and the lack of BPS permeability, then one would expect that the effects observed with Dip might be reproduced with BPS when cells are grown for longer times so that the intra-cellular iron reserves are depleted after some cell divisions. Future experiments should confirm this interpretation.
A recent study has examined differences in the gene expres-sion levels ofS. pombein response to iron availability (53). In the presence of the iron chelator ferrozine, which is not mem-brane permeable (42), 24 genes were repressed ⬎1.5-fold, while here we found 86 genes to be downregulated in a Php4-and Dip-dependent manner. This difference may be due to the use of different iron chelators. To clarify this point, we assayed the iron-dependent regulation ofpcl1⫹gene expression using both ferrozine and Dip. While we found no significant alter-ation in the transcription ofpcl1⫹mRNA in response to fer-rozine (Alexandre Mercier and Simon Labbe´, unpublished data),pcl1⫹transcript levels were downregulated 7.0-fold after treatment with Dip. Differences between the two studies may also be explained by the fact that cells were not treated at the same optical density with iron chelators. Therefore, experi-mental setups, including the nature of the iron chelator, time courses, and the duration of treatment, may account for dif-ferences, thereby complicating any comparison of expression profiles between the two studies.
To identify genes that exhibit iron- and php4⫹-dependent changes on a genome-wide scale, we carried out two types of DNA microarray experiments. First, we compared the mRNA
expression profile of wild-type cells grown under iron-replete conditions to cells grown under conditions of iron deprivation. We found that 251 genes were upregulated in response to iron deficiency. Among these genes, many of them code for known components of the reductive and nonreductive iron transport systems (26). We also found several uncharacterized genes (see Table S2 in the supplemental material). Interestingly, the
SPBC359.05-encoded permease is predicted to be an ABC
transporter that is thought to be localized to the vacuolar membrane (20). Because there is no known vacuolar oxidase-permease iron-transporting complex in S. pombe, this gene provides an attractive candidate for future study. We also iden-tified 325 genes that were expressed at higher levels in high-iron medium (see Table S1 in the supplemental material). In the second experiment, we compared the mRNA expression profile of wild-type cells grown under low iron conditions to that ofphp4⌬mutant cells grown under these same conditions. Messenger RNAs corresponding to 132 genes were signifi-cantly upregulated in the absence ofphp4⫹. Of the 132 genes, 33% are predicted to encode proteins involved in iron-depen-dent biochemical pathways. Based on the prediction that Php4 target genes would be expressed at higher levels in iron-replete wild-type cells than in iron-deficient cells and that they would be expressed at higher levels in iron-limitedphp4⌬mutant cells than in iron-limited wild-type cells, the overlap of the two sets of arrays includes 86 genes (Table 2). Among these 86 genes, 55% encode proteins that have been assigned a probable func-tion in iron-related processes based on homology with a known
FIG. 8. Php4 constitutively represses its target genes in cells depleted of GSH. (A) The isogenic strainsgcs1⫹(WT) andgcs1⌬were cultivated in yeast extract plus supplements and then washed twice. Cells were subsequently inoculated in selective Edinburgh minimal medium lacking GSH for 16 h, at which step the cultures were divided and then incubated in the absence (-) or presence of Dip (D) (250M) or FeCl3(Fe) (100M) for 90 min. Total RNA from the wild type (gcs1⫹) and thegcs1⌬disruption strain was analyzed by RNase protection assays.isa1⫹,php4⫹,fio1⫹,
fep1⫹, andact1⫹mRNA levels are indicated by arrows. Each culture was assayed three times. (B) Graphic representations of quantifications are shown. Values are the averages of triplicate determinations⫾standard deviations.