Jolanta Sarowska
1, Gabriela Bugla-Płoskońska
2, Bożena Futoma-Kołoch
2,
Zuzanna Drulis-Kawa
3The Sensitivity Level of
Salmonella
Enterica
ESBL
+Transconjugants to Normal Human Serum Correlated
with OMP Band Patterns Obtained by SDS-PAGE
Poziom wrażliwości transkoniugantów
Salmonella
enterica
ESBL
+na działanie normalnej surowicy ludzkiej w korelacji z profilami OMP
uzyskanymi metodą SDS-PAGE
1 Department of Biology and Medical Parasitology, Wroclaw Medical University, Wrocław, Poland 2 Institute of Genetics and Microbiology, University of Wroclaw, Wrocław, Poland
3 Department of Pathogen Biology and immunology, University of Wroclaw, Wrocław, Poland
Abstract
Objectives. The aim of the study was to assess the susceptibility of Salmonella strains to the bactericidal activity of normal human serum (NHS) and to analyze the outer membrane protein (OMP) profiles.
Material and Methods. The clinical strain Klebsiella pneumoniae 20 and six Salmonella enterica strains – S. Enteritidis 518, S. Typhimurium 244 and S. Hadar 55 and their ESBL+ transconjugants S. Enteritidis 518/20,
S. Typhimurium 244/20 and S. Hadar 55/20, obtained with K. pneumoniae 20 as the donor of ESBL plasmid were used. The bactericidal activity of NHS was determined as described by Jankowski et al. Isolation of the OMPs was performed according to the procedure described by Murphy and Bartos. Discontinuous sodium dodecyl sulphate (SDS) gel electrophoresis was carried out as described by Laemmli.
Results. This study examined the susceptibility of Salmonella recipient strains and their ESBL+ transconjugants
belonging to three serovars – Enteritidis, Typhimurium and Hadar – to NHS. It was shown that the recipient strains (S. Enteritidis 518, S. Typhimurium 244, and S. Hadar 55) and the ESBL conjugative plasmid donor strain
(K. pneumoniae 20) were resistant to the bactericidal action of NHS, whereas three Salmonella transconjugants
identified as ESBL producers (S. Enteritidis 518/20, S. Typhimurium 244/20 and S. Hadar 55/20) demonstrated high sensitivity to serum. SDS-PAGE gel analysis showed that the parental strains and transconjugants exhibited different outer membrane protein patterns. The observed changes included the presence or absence of particular proteins.
Conclusions. It seems that the acquisition of new plasmids may have unfavorable consequences for pathogenic bacteria and increase their susceptibility to serum activity (Adv Clin Exp Med 2010, 19, 6, 669–677).
Key words:Salmonella, NHS, OMP.
Streszczenie
Cel pracy. Określenie podatności szczepów Salmonella na bakteriobójcze działanie normalnej surowicy ludzkiej (NHS) i analiza profili białek błony zewnętrznej (OMP) badanych szczepów.
Materiał i metody. W badaniach wykorzystano kliniczny szczep Klebsiella pneumoniae 20 oraz sześć izolatów
Salmonella enterica: S. Enteritidis 518, S. Typhimurium 244, S. Hadar 55, a także ich ESBL+ transkoniuganty:
S. Enteritidis 518/20, S. Typhimurium 244/20 i S. Hadar 55/20, otrzymane z użyciemu szczepu K. pneumoniae 20, jako dawcy plazmidu typu ESBL. Bakteriobójczą aktywność NHS oceniano metodą opisaną przez Jankowskiego et al. Izolację białek OMP przeprowadzono zgodnie z procedurą opisaną przez Murphy’ego i Bartosa. Elektroforezę białek na żelu poliakrylamidowym w obecności SDS wykonano w układzie Laemmli’ego.
Wyniki. W badaniach określono wrażliwość na działanie NHS wyjściowych szczepów biorców Salmonella oraz ich ESBL+ transkoniugantów należących do trzech serowarów: Enteritidis, Typhimurium i Hadar. Wykazano, że
zarówno szczepy biorców (S. Enteritidis 518, S. Typhimurium 244, S. Hadar 55), jak i szczep dawcy plazmidu typu ESBL (K. pneumoniae 20) były oporne na bakteriobójcze działanie NHS, a trzy uzyskane transkoniuganty ESBL+
Adv Clin Exp Med 2010, 19, 6, 669–677 ISSN 1230-025X
ORIGINAL PAPERS
The Salmonella enterica Enteritidis serotype has been found to predominate among the non-typhoidal Salmonella species responsible for infec-tions in humans, and is one of the most frequent etiological factors in severe gastroenteritis in chil-dren both in Poland and in the majority of Euro-pean Union countries. S. Typhimurium is the next most commonly isolated type, followed by S. Hadar, S. Virchow, and S. Infantis, which are less common [1, 2]. An increase in the participation of multire-sistant Salmonella species in human infections has recently been observed all over the world [3]. The resistance of Salmonella species isolated in Poland to antibacterial agents has also increased [4].
Apart from salmonellosis of typical location, extraintestinal infections caused by Salmonella rods are becoming increasingly common [5]. These infections are generally characterized by a severe course, and the majority of the risk-group patients (children below the age of 4 and people over 60) require adequate antimicrobial therapy as well as hospitalization [2, 6]. Extended-spectrum cephalosporins are the basic treatment for serious infections due to nontyphoidal Salmonella spp. and have been used as the drugs of choice in treat-ing extraintestinal salmonellosis. The resistance of Salmonella to beta-lactam antibiotics is always acquired. As the information for this type of re-sistance is mostly included in the plasmid’s genes, it can be easily transferred between different spe-cies of Enterobacteriaceae that coexist in the hu-man intestine [7]. Conjugation is regarded as one of the main pathways for horizontal gene transfer among bacteria, and it requires donor and recipi-ent strains. Data published by many authors con-firm the effectiveness of the conjugational transfer of plasmid-located genes coding for ESBLs [8, 9].
This study examined the susceptibility of Sal-monella ESBL+ transconjugants belonging to three
serovars – Enteritidis, Typhimurium, and Hadar – to normal human serum (NHS). Unexpectedly, these strains became more susceptible to NHS. To establish a possible relationship between sensitiv-ity to serum and the OMP profiles of Salmonella strains, the susceptibility to the bactericidal effect of NHS and OMP band patterns were studied.
Material and Methods
The clinical strain Klebsiella pneumoniae 20 and six Salmonella enterica strains – S. Enteriti-dis 518, S. Typhimurium 244 and S. Hadar 55 and their ESBL+ transconjugants S. Enteritidis 518/20,
S. Typhimurium 244/20 and S. Hadar 55/20, ob-tained with K. pneumoniae 20 as the donor of ESBL plasmid were used. The transmission of ESBL plasmids and the antibiotic susceptibility of the strains were described in a previous study [10]. The parental Salmonella strains were isolated from fecal samples from children with gastroenteritis treated at the Lower Silesian Pediatric Center in Wrocław, Poland [2].
Normal human serum (NHS) was obtained by pooling sera from five healthy adult volunteers. The NHS samples were collected and kept frozen in 0.5 ml portions at –70°C. A suitable volume of the serum was thawed immediately before each ex-periment and was used only once.
Determination of C3 and C4
Complement Protein Levels
and IgG and IgM Antibody
Levels in NHS
The levels of C3, C4, IgG, and IgM were de-termined by the single radial immunodiffusion method using specific antibodies. In this meth-od, antigen-antibody precipitation was observed. Agarose plates with monospecific C3, anti-C4, anti-IgG and anti-IgM polyclonal antibodies (MEGA, Gliwice, Poland) were used. The investi-gation was carried out according to the producer’s instructions.
Bactericidal Activity of Serum
The bactericidal activity of NHS was deter-mined as described by Jankowski et al. [11]. Brief-ly, the strains were grown overnight and then bac-terial cells of the early exponential growth phase were transferred to fresh nutrient broth (Biomed, Warsaw) and incubated at 37°C for 0.5 hour. Af-ter incubation, the bacAf-terial cells were centrifuged (4000 rpm for 20 minutes). The bacteria fromSalmonella (S. Enteritidis 518/20, S. Typhimurium 244/20, S. Hadar 55/20) okazały się bardzo wrażliwe na działanie
surowicy. Rozdział SDS-PAGE wykazał różnice na poziomie OMP między szczepami wyjściowymi Salmonella, a ich ESBL+ transkoniugantami. Obserwowane zmiany polegały na obecności lub braku poszczególnych białek.
Wniosek. Prawdopodobnie pozyskanie nowego plazmidu przez komórki bakteryjne wpłynęło na niekorzystny dla tych patogenów wzrost podatności na aktywność surowicy (Adv Clin Exp Med 2010, 19, 6, 669–677).
a young log-phase culture were added to 50% NHS (diluted with NaCl) in a final concentration of 106
CFU/ml. The samples were incubated in a water bath at 37°C. After 0, 60 and 180 minutes of in-cubation, the samples were collected, diluted and cultured on nutrient agar plates. After 18 hours of incubation at 37°C, the colony-forming units (CFU/ml) were calculated. The bacterial number at time 0 was taken as 100% cell survival. Strains with survival rates > 100% in serum after 180 min-utes (3 hours) of incubation were considered resis-tant (NHSR) and those with survival rates < 100%
were considered susceptible to the bactericidal ac-tion of the serum (NHSS).
The control of experiments was NHS decomple-mented by heating the sample at 56°C for 30 min.
Isolation of OMPs
In the present study the zwitterionic deter-gent Zwitterdeter-gent Z 3–14® (Calbiochem, Behring)
was used for the isolation of bacterial OMPs from the Salmonella strains. The isolation of OMPs was determined as described previously [12–14]. Zwitterionic detergents lack conductivity and electrophoretic mobility and are also suited for breaking protein-protein interactions. The addi-tion of zwitterionic detergents can also improve protein solubilization [15]. Isolation of the OMPs was performed according to the procedure de-scribed by Murphy and Bartos [16] with some mi-nor modifications. The OMPs were isolated from bacteria grown in 100 ml of Brain Heart Infusion broth (Difco) at 37°C for 18 h. After incubation, the bacterial cells were centrifuged (4000 rpm for 20 min at 4°C) and the pellet was suspended in 2.5 ml of buffer B (1 M sodium acetate, 0.001 M β–mercaptoethanol, pH 4.0). Then 22.5 ml of solution containing 5% Zwittergent Z 3–14 and 0.5 M CaCl2 was added. This solution was stirred
for 1 h at room temperature. To precipitate the nucleic acids 6.25 ml of 96% cold ethanol was added very slowly. The mixture was centrifuged at 12 300 rpm for 10 min at 4°C. The remaining proteins were precipitated by addition of 93.5 ml of 96% cold ethanol and centrifuged at 12 300 rpm for 20 min at 4°C. The pellet was suspended in 5 ml of buffer Z (0.05% Zwittergent, 0.05 M Tris, and 0.01 M EDTA, pH 8.0) and stirred for 1 h at room temperature. The solution was kept at 4°C overnight and centrifuged at 8 700 rpm for 10 min at 4°C. OMPs were present in the buffer Z soluble fraction after centrifugation. After the preparation of the OMPs we checked the enzymatic activity of succinic dehydrogenase, a marker for the cytoplas-mic membranes, in the soluble fraction of buffer Z using the method of Rockwood et al. [17]. The
Bradford protein assay was used to determine the concentration of proteins in the samples [18].
The samples of Salmonella OMPs were tested for the presence of biological membranes. When the preparations were assayed for the enzymatic activity of succinic dehydrogenase, the Zwitter-gent-extracted OMPs from the tested Salmonella strains contained no detectable succinic dehydro-genase activity, which confirms that they were free of membrane contaminations.
Zwitterionic detergents have found wide-spread use in electrophoresis [19]. The Zwitter-gent Z 3–14® deterZwitter-gent has been found suitable for Escherichia coli OMP separation by two-di-mensional electrophoresis (2-DE) using the pH 3–10 immobilized pH gradient IPG strips, and for Salmonella OMP separation by 2-DE in a capillary tube system [20].
Polyacrylamide
Gel Electrophoresis
Discontinuous sodium dodecyl sulphate (SDS) gel electrophoresis was carried out on slabs with 12.5% acrylamide, as described by Laemmli [21]. The samples were applied to the slabs after heating at 100°C for 4 minutes. The SDS-PAGE Molecular Weight Standards (broad range) 161-0317 (Bio-Rad) were used for molecular weight calibration. They contain nine proteins from 6.5–200 kDa (myosin-200, ß-galactosidase-116.25, phosphory-lase b-97.4, serum albumin-66.2, ovalbumin-45, carbonic anhydrase-31, trypsin inhibitor-21.5, ly-sozyme-14.4, and aprotinin-6.5 kDa). OMPs pre-pared in the sample buffer were loaded into each of the wells of the gels in similar concentrations (8–10 µg/well). Electrophoresis was carried out at
100 V for 25 minutes and then at 200 V for 45 min-utes. When the electrophoresis was completed, the gel was placed in a solution containing 25% meth-anol, 10% acetic acid and 0.05% Coomassie bril-liant blue (SIGMA) for 1 hour. Then the gel was decolorized in 10% acetic acid for 3 to 5 hours.
Molecular Analysis of OMPs
Quantity One v. 4 (Bio-Rad) was used for the molecular analysis of the OMPs. Similarity densitograms for some protein inclusions were constructed with the UPGMA (Unweighted Pair Group Method, Arithmetic Mean) algorithm.Results and Discussion
pneumoniae strains to clinical Salmonella isolates belonging to three serovars – Enteritidis, Ty-phimurium and Hadar – which leads to Salmonella resistance to the beta-lactam antibiotics most often used in treatment. It was concluded that “the effi-ciency of ESBL plasmid transferability depended on the Salmonella serovar. The Salmonella enterica se-rovar Enteritidis strains were the best ESBL plasmid recipients among the clinical Salmonella serovars” [10]. Conjugation efficiency was found from 10–4 to
10–8 per donor cell. Conjugation experiments and
plasmid analysis revealed the presence of the same plasmids in the Klebsiella donor strain and in the Salmonella ESBL+ transconjugants [10].
ESBL-posi-tive transconjugants, like the recipients, were found to be fully sensitive to amikacin, ciprofloxacin and trimethoprim-sulfametoksazole. All Salmonella transconjugants were characterized by resistance to ampicillin and resistance or reduced sensitivity to ceftazidime and cefotaksime, and they were posi-tive in ESBL tests (DDST and E-test ESBL).
It should be noted that some R plasmids could simultaneously encode resistance and virulence factors. It is therefore important to test features other than the drug resistance of transconjugants receiving a plasmid determining ESBL production. The focus of the present study was assessment of the strains’ susceptibility to the bactericidal activity of normal human serum (NHS) and analysis of the outer membrane proteins (OMP) profiles. Kustos et al. noted that outer membrane proteins are nec-essary factors of bacterial cells and play important role in several essential functions of the micro-organisms: serum sensitivity, drug susceptibility and pathogenicity [22].Some OMPs undoubtedly amplify the sensitivity of bacteria such as Neisseria meningitidis [23], Aeromonas hydrophila [24], and K. pneumoniae [25] to the bactericidal activity of serum. Other OMPs enhance bacterial resistance to the cytolytic activity of complement, for exam-ple DsrA (Haemophilus ducreyi) [26] and UspA2 (Moraxella catarrhalis) [14, 27]. For Salmonella strains, Bugla et al. [28] also suggested the partici-pation of OMPs in modulating the susceptibility of Salmonella rods to serum.
The complement cascade is part of both the adaptive and innate immune systems and consists of at least 35 proteins, mostly in pre-activated en-zymatic forms. It is set off within seconds when a foreign pathogen enters. As described in a previ-ous study [36], activation of the complement cas-cade is achieved through three major pathways: the classical (CP), the alternative (AP), and the lectin (LP) [29, 30]. The CP is activated mainly by com-plexes of antigen and antibody (IgG1, IgG2, IgG3, or IgM) [31]. The AP is important in innate host defense against bacterial infection. Amplification of
the AP can be stimulated by invading microorgan-isms in the absence of specific antibodies [31, 32]. The LP begins when mannose-binding lectin (MBL) binds to microbial surface carbohydrates. MBL cooperates with the proenzymes MASP1, MASP2, MASP3 and Map19, which are associated with MBL in the presence of calcium ions (Ca2+).
Ficolins of types H, L, and M as well as MBL initi-ate the LP pathway [33]The CP, AP, and LP pro-ceed to the formation of the C5b678-9 membrane attack complex (MAC), capable of insertion into biological membranes and causing cell lysis and death. It has been accepted that the complement system is important in protecting against micro-bial infection, but also contributes to the patho-physiology of a number of non-infectious diseases [29]. The resistance of Gram-negative bacteria to serum’s lytic activity may be one of the virulence factors essential in the development of sepsis.
Research on OMPs extracted from the tested Salmonella strains is very important because of their accessibility to the host defense system. The results of sensitivity to the bactericidal action of NHS and the SDS-PAGE analysis of the OMPs of Salmonella transconjugant reveal that the paren-tal strains and transconjugants exhibited different outer-membrane banding patterns.
Susceptibility to NHS was assessed in three serovars of Salmonella: Enteritidis, Typhimurium and Hadar, before and after conjugation with the K. pneumoniae 20 strain used as the donor of ESBL conjugative plasmid. The level of C3 in NHS was determined to be 90 mg/dl (standard for NHS: 55–120 mg/dl) and the level of the C4 component was 48 mg/dl (standard for NHS: 20–50 mg/dl). The level of IgM was 109 mg/dl (standard for NHS: 60–280 mg/dl); the level of IgG was 1418 mg/dl (standard for NHS: 750–1500 mg/dl).The sensi-tivities of the tested strains to 50% NHS are given in Table 1. It was shown that the recipient strains (S. Enteritidis 518, S. Typhimurium 244 and S. Ha-dar 55) and the ESBL conjugative plasmid donor strain (K. pneumoniae 20) were NHSR. After
treat-ing the bacterial cells with 50% NHS, the percent survival of bacterial cells after 3 hours of incuba-tion was 241.9 and 1585.4 for K. pneumoniae 20 and S. Hadar 55, respectively. Two Salmonella transconjugants (S. Enteritidis 518/20 and S. Ty-phimurium 244/20) identified as ESBL producers demonstrated high sensitivity to serum. NHS dem-onstrated bacteriostatic activity against S. Hadar 55/20. The percent survival of these bacterial cells was from > 0.001 to 75.00 after 3 hours of incuba-tion. The bacteria proliferated very intensively in the inactivated at 56°C for 30 min NHS.
with ceftazidime was investigated at the same time. The recipients as well as the transconjugants in both mediums presented a similar level of vi-ability, estimated as the numbers of colonies on agar plates (data not published).
The sensitivity of Gram-negative bacteria to the bactericidal activity of NHS depends on the structure and organization of the bacterial outer membrane [28, 34]. It is known that the structure of O-specific side chains of the lipopolysaccharide (LPS) and the OMPs play important roles in the resistance of bacterial cells to the lytic activity of complement [28, 29]. In general, rough strains (R) of Gram-negative bacteria, in which the LPS lacks an O-antigenic polysaccharide chain, are sensitive to the bactericidal action of serum, while smooth (S) strains are resistant [29].In the case of Salmo-nella strains, resistance to serum may be enhanced by the Rck (17–19 kDa) and PagC (17–19 kDa) outer membrane proteins. It has been confirmed that the rck gene was present in virulent S. Enter-itidis and S. Typhimurium serovars [35].
The results of the current studyindicate that in intact populations (before conjugation) of the Sal-monella strains (S. Typhimurium 244, S. Enteriti-dis 518) with the confirmed presence of virulence plasmids, cells that were resistant to 50% NHS dominated [10]. However, S. Hadar 55, which lacked virulence plasmids, also demonstrated low susceptibility to the lytic activity of NHS. Unex-pectedly, Salmonella transconjugants carrying ad-ditional ESBL+ plasmids showed high sensitivity to
NHS. It seems that the acquisition of new plasmids may have unfavorable consequences for
pathogen-ic bacteria and increase their susceptibility to se-rum activity. It was established by, among others, Jankowski and Cisowska [34] that the E. coli K12 W1485 strain that obtained resistance plasmids (R577 and R785) for tetracycline, chlorampheni-col, vibramycin, sulphonamide and streptomycin became sensitive to NHS. The data presented in this paper also confirm this phenomenon. It is very interesting that some R plasmids can increase the susceptibility of bacteria to the action of sera. A probable explanation of this could be the re-modeling of the envelope of the bacterial cells – e.g., the OMP composition after the conjugation process. Chaffer et al. [36] showed that E. coli O2 isolates that possess 35kDa OMA are highly resis-tant to the bactericidal effect of serum. They also demonstrated that the presence of a strong 35 kDa OMP band contributes to complement sensitivi-ties and suggested that these membrane’s changes resulted in a possibility of complement attack [36]. The data presented in this paper indicate a similar dependence (Figure 1, Table 2).
Lin et al. noted that many OMPs are involved in bacterial virulence, particularly serum resistance [37]. Cirillo et al. pointed out that Rck gene (17– –19 kDa OMP) located on The S. Typhimurium virulence plasmid is associated with high-level se-rum resistance [35]. This OMP inhibits comple-ment lysis by preventing the polymerization of the C9 protein of the complement cascade. In the present study, all the parental and transconjugant strains showed the presence of the 17–19 kDa OMP (probably Rck) regardless of differences in serum susceptibility. It seems that it is not the
Table 1. Bactericidal activity of 50% NHS against K. pneumoniae 20 (donor), S. Enteritidis 518, S.Typhimurium 244,
S. Hadar 55 (recipients), and S. Enteritidis 518/20, S. Typhimurium 244/20, S. Hadar 55/20 (transconjugants)
Tabela 1. Bakteriobójcza aktywność 50% NHS wobec K. pneumoniae 20 (dawca), S. Enteritidis 518, S. Typhimurium 244,
S. Hadar 55 (biorcy) i S. Enteritidis 518/20, S. Typhimurium 244/20, S. Hadar 55/20 (transkoniuganty)
Strains
(Szczepy) T0
a
(CFU/ml)b T(CFU/ml)60 T(CFU/ml)180 %
b
K. pneumoniae 20 6.2 × 105 4.7 × 105 1.5 × 106 241.93
S. Enteritidis 518 1.2 × 106 1.4 × 106 9.0 × 106 750.00
S. Enteritidis 518/20 1.3 × 106 2.0 ×´ 101 1.0 × 101 < 0.01
S. Typhimurium 244 6.0 × 105 2.0 × 106 3.5 × 106 583.33
S. Typhimurium 244/20 5.0 × 105 1.0 × 102 1.7 × 101 < 0.01
S. Hadar 55 4.1 × 105 1.2 × 106 6.5 × 106 1585.37
S. Hadar 55/20 3.2 × 105 1.5 × 105 2.4 × 105 75.00
a 100% survival of bacterial cells in serum. a 100% przeżywalność bakterii w surowicy.
% b percent of survival bacterial cells in serum after 3 h of incubation.
presence of Rck protein in Salmonella ESBL+
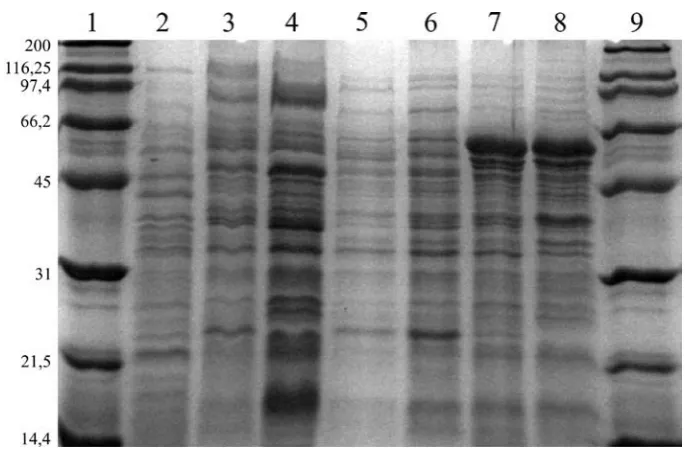
tran-sonjugants that determines resistance to serum. The SDS-PAGE gel analysis (Table 2) showed that the parental strains and transconjugants ex-hibited different outer membrane protein patterns. The changes included the presence or absence of particular proteins. New OMPs (absent before conjugation) with molecular masses of 25 kDa and 44 kDa were observed in the S. Enteritidis 518/20 isolate. A similar situation was documented for the S. Typhimurium 244/20 strain (43 kDa). There was only one case of a protein with a similar molecular weight (43 kDa) in the K. pneumoniae 20 donor strain and the transconjugant S. Typhimurium 244/20. The acquisition of ESBL plasmids also resulted in the loss of some OMPs in the trans-conjugants compared with the parental strains. In the case of the S. Enteritidis 518/20 isolate, no bands for the 15, 22, 53, 58 and 78 kDa proteins were noted. For the remaining transconjugants, no bands were observed for the 16, 38, 44, 55 and 82 kDa proteins (S. Typhimurium) and 27, 65 and 66 kDa (S. Hadar 55/20).
As noted above, the mechanisms of various OMPs‘ role in serum resistance have still not been precisely explained. Lin et al. pointed out, OMPs of Gram-negative bacteria promote bacterial resis-tance to complement action [37], but – as noted above – there are some OMPs which certainly in-crease bacterial sensitivity to the bactericidal
activ-ity of serum. Galdiero et al. [38] also showed that S. Typhimurium porins induce an inflammatory effect by activating complement, acting on both the classical and the alternative pathways. The 44 kDa protein was proposed by Negm and Pistole as the OmpD-like protein that recognizes S. Ty-phimurium by macrophages in the first stages of infection; minimal concentrations of this protein could block the attachment of S. Typhimurium to macrophages. [39] However, the OMPs of Salmo-nella provide specialized functions that enable the organisms to survive in the host environment, and a specific OMP can be associated with the pheno-typic expression of virulence [35]. Gram-negative bacteria are almost always resistant to the bacteri-cidal action of serum, but there are some Gram- -negative strains that are sensitive to the lytic ac-tion of complement [28, 40].
The acquisition of R plasmid (ESBL) seems to be favorable for pathogenic Salmonella strains be-cause it enables them to survive in the presence of antibiotic. However, simultaneous expression of R plasmid genes can lead to dramatic changes in the outer membrane protein profile, which result in an increase in serum susceptibility. In conclu-sion, the results of these investigations suggest that ESBL-positive Salmonella transconjugants obtained by conjugation would be eliminated by an efficiently functioning immune system during systemic infections or bacteremia.
Fig. 1. The SDS-PAGE patterns of outer membrane proteins (OMPs). Lanes 1 and 9: SDS-PAGE Molecular Weight Standards (Broad Range) 161-0317 (Bio-Rad); Lane 2: OMPs isolated from K. pneumoniae 20; Lane 3: S. Enteritidis
518; Lane 4: S. Enteritidis 518/20; Lane 5: S. Typhimurium 244; Lane 6: S. Typhimurium 244/20; Lane 7: S. Hadar 55; Lane 8: S. Hadar 55/20
Ryc. 1. Profile SDS-PAGE białek błony zewnętrznej (OMP). Ścieżki 1 i 9: SDS-PAGE – standardy mas
cząsteczkowych (Broad Range 161-0317 – Bio-Rad), ścieżka 2: OMPs wyizolowane z komórek K. pneumoniae 20; ścieżka 3: S. Enteritidis 518; ścieżka 4: S. Enteritidis 518/20; ścieżka 5: S. Typhimurium 244;
Table 2. OMP profiles of: aD – donor, aR1 – recipient and aTR1 – transconjugant, bD – donor, bR2 – recipient and bTR2 – transconjugant and cD – donor, cR3 – recipient and cTR3 – transconjugant Tabela 2. Profile OMP: aD – dawca, aR1 – biorca, aTR1 – transkoniugant, bD – dawca, bR2 – biorca, bTR2 – transkoniugant, cD – dawca, cR3 – biorca, cTR3 – transkoniugant Strains (Szczepy) OMPs (kDa) 84 82 78 67 66 65 58 55 53 51 44 43 41 38 35 33 32 29 28 27 25 23 22 19 17 16 15 14 a, b, cD + – – + – – + – – – – + – – – + – + – + – + + + – + – + aR1 – – + – – – + – + – – – – – + – + + – + – + + + + – + – aTR1 – – – – – – – – – – + – – – + – + + – + + + – + + – – – bR2 – + – – – + – + – – + – – + + – + + – + + + + + + + + – bTR2 – – – – – + – – – – – + – – + – + + – + + + + + + – + – cR3 – – – – + + – – – + + – + – + – + – + + – + – + + – + – cTR3 – – – – – – – – – + + – + – + – + – + – – + – + + – + – D – donor ESBL + – K. pneumoniae 20 – NHS
R –
(total concentration of OMPs – 491.117 µg/ml) R1 – recipient ESBL – – S . Enteritidis 518 – NHS
R –
(total concentration of OMPs – 393.694 µg/ml) TR1 – transconjugant ESBL + – S . Enteritidis 518/20 – NHS
S –
(total concentration of OMPs – 1.027.664 µg/ml) R2 – recipient ESBL
– S. Typhimurium
244
–
NHS
R –
(total concentration of OMPs – 355.799 µg/ml) TR2 – transconjugant ESBL +S . Typhimurium 244/20 – NHS S– (total concentration of OMPs – 589.295 µg/ml) R3 – recipient ESBL
– S. Hadar
55
–
NHS
R –
(total concentration of OMPs – 610.524 µg/ml) TR3 – transconjugant ESBL +S . Hadar 55/20 – NHS S– (total concentration of OMPs – 699.792 µg/ml) D – dawca ESBL + – K. pneumoniae 20 – NHS
R –
(całkowite stężenie OMP – 491,117 µg/ml) R1 – biorca ESBL – – S . Enteritidis 518 – NHS
R –
(całkowite stężenie OMP – 393,694 µg/ml) TR1 – transkoniugant ESBL + – S . Enteritidis 518/20 – NHS
S –
(całkowite stężenie OMP – 1027,664 µg/ml) R2 – biorca ESBL
– S. Typhimurium
244
–
NHS
R –
(całkowite stężenie OMP – 355,799 µg/ml) TR2 – transkoniugant ESBL +S . Typhimurium 244/20 – NHS S– (całkowite stężenie OMP – 589,295 µg/ml) R3 – biorca ESBL
– S. Hadar
55
–
NHS
R –
Acknowledgments
We would like to thank Magdalena Kasendra, MSc, for her contribution to SDS-PAGE and figure preparation.
References
[1] Threlfall E: Salmonella. [In:] Microbiology and Microbial Infections. Ed. Boriello S, ASM Press, Washington 2005, 10th ed, 54, 1398–1427.
[2] Sarowska J, Drulis-Kawa Z, Korzekwa K, Lewczyk E, Jankowski S, Doroszkiewicz W: The occurrence of acute diarrhoeas induced by rotaviruses and Salmonella strains in children hospitalised in the Lower Silesian J. Korczak Paediatrics Centre in Wroclaw. Adv Clin Exp Med 2005, 14, 759–763.
[3] Threlfall E: Antimicrobial drug resistance in Salmonella: problems and perspectives in food- and water-borne infections. FEMS Microbiol Rev 2002, 26, 141–148.
[4] Lazinska B, Rokosz A, Sawicka-Grzelak A, Luczak M: Strains of genus Salmonella isolated from extraintestinal infections. Med Dosw Mikrobiol 2005, 57, 287–294.
[5] Diez-Dorado R, Tagarro-Garcia A, Baquero F: Non-typhoid Salmonella bacteriemia. An Ped (Barc) 2004, 60, 344–348.
[6] Saps M, Pensabene L, Di Martino L, Staiano A, Wechsler J, Zheng X, Di Lorenzo C: Post-infectious functional gastrointestinal disorders in children. J Pediatr 2008, 152, 812–816.
[7] Miriagou V, Tassios P, Legakis N, Tzouvelekis L: Expanded-spectrum cephalosporin resistance in non-typhoid
Salmonella. Int J Antimicrob Agents 2004, 23, 547–555.
[8] Franiczek R, Dolna I, Krzyzanowska B, Szufnarowski K, Kowalska-Krochmal B: Conjugative transfer of multi-resistance plasmids from ESBL-positive Escherichia coli and Klebsiella spp. clinical isolates to Escherichia coli strain K12 C600. Adv Clin Exp Med 2007, 16, 239–247.
[9] Rasheed JK, Anderson GJ, Yigit H, Queenan AM, Doménech-Sánchez A, Swenson JM, Biddle JW, Ferraro MJ, Jacoby GA, Tenover FC: Characterization of the Extended-Spectrum β-Lactamase reference Strain, Klebsiella
pneumoniae K6 (ATCC 700 603), which produces the novel enzyme SHV-18. Antimicrob Agents Chemother
2000, 44, 2382–2388.
[10] Sarowska J, Drulis-Kawa Z, Guz K, Jankowski S, Wojnicz D: Conjugative transfer of plasmid encoding extend-er-spectrum beta-lactamase to recipient Salmonella strains. Adv Clin Exp Med 2009, 18, 63–70.
[11] Jankowski S, Rowiński S, Cisowska A, Gamian A: The sensitivity of Hafnia alvei strains to the bactericidal effect of serum. FEMS Immunol Med Microbiol 1996, 13, 59–64.
[12] Futoma-Kołoch B, Bugla-Płoskońska G, Doroszkiewicz W, Kaca W: Survival of Proteus mirabilis O3 (S1959), O9 and O18 strains in Normal Human Serum (NHS) correlates with the diversity of their outer membrane pro-teins (OMPs). Pol J Microbiol 2006, 55, 153–156.
[13] Bugla-Płoskońska G, Futoma-Kołoch B, Skwara A, Doroszkiewicz W: Use of zwitterionic type of detergent in isolation of Escherichia coli O56 outer membrane proteins improves their two-dimensional electrophoresis (2-DE). Pol J Microbiol 2009, 58, 205–209.
[14] Bugla-Płoskońska G, Korzeniowska-Kowal A, Guz-Regner K: Reptiles as a source of Salmonella O48 – clinically important bacteria for children: the relationship between resistance to normal cord serum and outer membrane protein patterns. Microb Ecol 2010.
[15] Shaw MM, Riederer BM: Sample preparation for two-dimensional gel electrophoresis. Current Advancements in the Methodology. GIT Laby J 2006, 6, 302–303.
[16] Murphy T, Bartos LC: Surface-exposed and antigenically conserved determinants of outer membrane proteins of
Branhamella catarralis. Infect Immun 1989, 10, 2938.
[17] Rockwood D, Wilson MT, Darley-Usmar VM: Isolation and characteristic of intact mitochondria. pp. 1–16. In: Mitochondria: a practical approach. Eds.: Darley-Usmar VM, Rickwood D, Wilson MT. IRL Press, Oxford, 1987.
[18] Bradford MM: A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976, 72, 248–254.
[19] Luche S, Santoni V, Rabilloud T: Evaluation of nonionic and zwitterionic detergents as membrane protein solu-bilizers in two-dimensional electrophoresis. Proteomics 2003, 3, 249–253.
[20] Futoma-Kołoch B, Bugla-Płoskońska G, Doroszkiewicz W: Isolation of outer membrane proteins (OMP) from
Salmonella cells using zwitterionic detergent and their separation by two-dimensional electrophoresis (2-DE). Pol
J Microbiol 2009, 58, 363–366.
[21] Laemmli UK: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 15, 680–685.
[22] Kustos I, Kocsis B, Kilar F: Bacterial outer membrane protein analysis by electrophoresis and microchip technol-ogy. Expert Rev Proteomics 2007, 4, 91–106.
[23] Zollinger WD, Boslego J, Froholm LO, Ray JS, Moran EE, Brandt BL: Human bactericidal antibody response to meningococcal outer membrane protein vaccines. Ant Leeuwenhoek 1987, 53, 403–411.
[25] Alberti S, Marquez G, Camprubi S, Merino T, Vivanco F, Benedi J: C1q binding and activation of the comple-ment classical pathway by Klebsiellapneumoniae outer membrane proteins. Infect Immun 1993, 61, 852–860.
[26] White CD, Leduc I, Olsen B, Jeter C, Harris C, Elkins C: Haemophilusducreyi outer membrane determinants, including DsrA, define two clonal populations. Infect Immun 2005, 4, 2387–2399.
[27] Attia AS, Lafontaine ER, Latimer JL, Aebi C, Syrogiannopoulos GA, Hansen EJ: The UspA2 protein of Moraxella
catarrhalis is directly involved in the expression of serum resistance. Infect Immun 2005, 73, 2400–2410.
[28] Bugla G, Korzeniowska-Kowal A, Gamian A, Doroszkiewicz W: Bactericidal activity of serum against Salmonella
O48 serovars. Int. J Antimicrob Agents 2004, 24, 604.
[29] Mielnik G, Gamian A, Doroszkiewicz W: Bactericidal activity of normal cord serum (NCS) against Gram-negative rods with sialic acid-containing lipopolysaccharides (LPS). FEMS Immunol Med Microbiol 2001, 31, 169–173
[30] Würzner R: Evasion of pathogens by avoiding recognition or eradication by complement, in part via molecular mimicry. Mol Immunol 1999, 36, 249–260.
[31] Seelen MA, Roos A, Wieslander J, Mollnes TE, Sjöholm AG, Wurzner R, Loos M, Tedesco F, Sim RB, Garred P, Alexopoulos E, Turner MW, Daha MR: Functional analysis of the classical, alternative, and MBL pathways of the complement system: standardization and validation of a simple ELISA. J Immunol Methods 2005, 1–2, 187–198.
[32] Arlaud GJ, Barlow PN, Gaboriaud C, Gros P, Narayana SV: Deciphering complement mechanisms: the contri-butions of structural biology. Mol Immunol 2007, 44, 3809–3822.
[33] Harmat V, Gál P, Kardos J, Szilágyi K, Ambrus G, Végh B, Náray-Szabó G, Závodszky P: The structure of MBL-associated serine protease-2 reveals that identical substrate specificities of C1s and MASP-2 are realized through different sets of enzyme-substrate interaction. J Mol Biol 2004, 342, 1533–1546.
[34] Jankowski S, Cisowska A: Plasmids R577 and R785 decrease the resistance of Escherichia coli K12 strain W1485 to bactericidal action of normal serum. J Appl Genet 1998, 39, 211–216.
[35] Cirillo DM, Heffernan EJ, Wu L, Harwood J, Fierer J, Guiney DG: Identification of a domain in Rck, a product of the Salmonella typhimurium virulence plasmid, required for both serum resistance and cell invasion. Infect Immun 1996, 64, 2019–2023.
[36] Chaffer M, Heller ED, Schwartsburd B: Relationship between resistance to complement, virulence and outer membrane protein patterns in pathogenic Escherichia coli O2 isolates. Vet Microbiol 1999, 64, 323–332.
[37] Lin J, Huang S, Zhang Q: Outer membrane proteins: key players for bacterial adaptation in host niches. Microb Infect 2002, 4, 325–331.
[38] Galdiero F, Tufano MA, Sommese L, Folgore A, Tedesco F: Activation of the complement system by porins extracted from Salmonella typhimurium. Infect Immun 1984, 46, 559–563.
[39] Negm RS, Pistole TG: Macrophages recognize and adhere to an OmpD-like protein of Salmonella typhimurium. FEMS Immunol Med Microbiol 1998, 20, 191–199.
[40] Bugla-Płoskońska G, Kiersnowski A, Futoma-Kołoch B, Doroszkiewicz W: Killing of Gram-negative bacteria with normal human serum and normal bovine serum: use of lysozyme and complement protein in the death of Salmonella
strains O48. Microb Ecol 2009, 58, 276–289.
Address for correspondence:
Jolanta Sarowska
Department of Biology and Medical Parasitology Wroclaw Medical University
Mikulicza-Radeckiego 9 50-367 Wrocław Poland
Tel.: +4871 784-1519
E-mail: [email protected] Conflict of interest: None declared Received: 9.09.2010