0095-1137/95/$04.0010
Copyrightq1995, American Society for Microbiology
Isolation of Human Immunodeficiency Virus Type 1 (HIV-1) RNA
from Feces by a Simple Method and Difference between
HIV-1 Subpopulations in Feces and Serum
L.
VAN DERHOEK,
1R. BOOM,
1J. GOUDSMIT,
1* F. SNIJDERS,
2ANDC. J. A. SOL
1Human Retrovirus Laboratory
1and Division of Infectious Diseases, Tropical Medicine and AIDS,
2Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands
Received 24 August 1994/Returned for modification 29 September 1994/Accepted 6 December 1994
A simple method for the isolation and subsequent detection of human immunodeficiency virus type 1 (HIV-1)
RNA from feces is described. Viral RNA was isolated by the method developed by Boom et al. (R. Boom,
C. J. A. Sol, M. M. M. Salimans, C. L. Jansen, P. M. E. Wertheim-van Dillen, and J. van der Noordaa, J. Clin.
Microbiol. 28:495–503, 1990), which was adapted for feces. HIV-1 RNA was detected by reverse transcription
(RT) followed by a nested PCR encompassing the V3 region. Reconstruction experiments revealed that the
efficiencies of the extraction technique and the subsequent RT-PCR were not considerably affected by the varied
composition of feces. The method was applied on fecal specimens from 18 HIV-1-infected individuals, among
which were samples that had been stored for 9 years. It appeared that HIV-1 RNA was detectable in the feces
of 12 persons (67%). Viral RNA was present in the feces of persons who fulfilled the criteria for CDC class II
and CDC class III HIV infection as well as in patients who were diagnosed with AIDS (CDC class IV). Direct
sequencing of amplimers obtained from paired fecal and serum specimens showed that differences in sequence
heterogeneity existed. In one patient a remarkable difference in the HIV-1 sequences between isolates from
feces and serum was observed. In conclusion, HIV-1 RNA is frequently present in the feces of HIV-1-infected
individuals, and in some cases the HIV-1 subpopulation in feces differs from the HIV-1 subpopulation in serum.
Gastrointestinal symptoms are common in human
immuno-deficiency virus (HIV)-infected patients with advanced stages
of HIV infection (15, 20–22, 27, 30). In a majority of patients
with gastrointestinal diseases, opportunistic infectious agents
can be identified; however, in a significant number of cases the
etiology of diarrhea remains unexplained (3, 5, 12, 37). It has
been suggested that HIV itself might play a role in the
patho-genesis of diarrhea by infecting cells of the intestinal mucosa
(11, 29, 38, 44, 46). This hypothesis is supported by in vitro
studies in which a pathological effect of HIV on cultured
epi-thelial cells of the intestinal mucosa was observed (1, 4, 16).
Moreover, in vivo infection by HIV of intestinal mucosal cells
has been identified by in situ hybridization, PCR, and cell
culture (18, 23, 25, 28, 32, 33, 35, 45). Characterization of HIV
type 1 (HIV-1) isolates recovered from the bowel showed that
the biological and serological properties of these strains can
differ from those of HIV-1 strains recovered from serum (2).
Until now studies on HIV infection of intestinal cells in vivo
are limited to those of cells in intestinal biopsy specimens. A
longitudinal study at different stages of HIV-1 infection is
needed to determine whether the mere presence of HIV-1 in
the intestine is linked to the onset of gastrointestinal
symp-toms. However, for ethical reasons, intestinal biopsy specimens
are difficult to collect from patients at different stages of
dis-ease, particularly at the asymptomatic stage. In vitro studies
have shown that following HIV-1 infection, bowel epithelial
carcinoma cells excrete viral particles at both the apical and
basolateral surfaces of the cell, indicating that virus might be
detectable in feces (17, 48). Longitudinal collection of fecal
specimens is easy and noninvasive. Therefore, feces is a
suit-able specimen from which viral nucleic acids that could have
been produced in the gastrointestinal tract can be isolated and
characterized.
Purification of viral RNA from feces is difficult by the most
common extraction methods known to date, presumably
be-cause of its extremely varying composition. We have previously
reported on a reliable and rapid procedure for the purification
of nucleic acids from clinical specimens like serum and urine
(6, 7). This procedure is based on the lysing and
nuclease-inactivating properties of the chaotropic agent guanidinium
thiocyanate (GuSCN) and the concurrent binding of all nucleic
acid types (DNA and RNA, both single and double stranded)
by silica particles in the presence of this agent. In this report we
describe a simple though effective modification of the basic
procedure for the purification of HIV-1 RNA from fecal
spec-imens. We validated the method in reconstruction experiments
in which fecal specimens which strongly varied in color and
consistency were spiked with HIV-1. These experiments
re-vealed that the detection of HIV-1 RNA is not considerably
influenced by the fecal composition.
Fecal specimens from 18 HIV-1-infected individuals were
tested for the presence of HIV-1 nucleic acids. HIV-1 RNA
could be detected in the feces of 12 individuals (67%). The
specificities of the PCR products were confirmed by direct
sequencing. To determine whether the sequences present in
feces originated from blood leakage into the intestine or
rep-resent a distinct subpopulation, amplified HIV-1 RNA from
corresponding serum samples was sequenced.
MATERIALS AND METHODS
Subjects.One or two fecal specimens were collected from 18 HIV-1-infected individuals (the characteristics of the individuals in the study group are given in Table 1). Two individuals (A and B) participated in a longitudinal cohort study among homosexual men (14). Fecal and serum specimens were collected at the beginning of a study performed in 1984, when the patients were diagnosed as having CDC class II HIV infection (9). Fecal and serum specimens were also * Corresponding author. Mailing address: Human Retrovirus
Lab-oratory, Academic Medical Center, University of Amsterdam, Meiberg-dreef 15, 1105 AZ Amsterdam, The Netherlands. Phone: (31-20) 5664853. Fax: (31-20) 6916531.
581
on May 15, 2020 by guest
http://jcm.asm.org/
obtained from these two individuals 9 years later. During this period individual A developed AIDS and individual B showed persistent generalized lymphade-nopathy (CDC class III). One male (individual C) presented in the clinic with acute HIV-1 infection (lymphadenopathy, fever, exanthema, HIV-1 p24 present in serum, no detectable antibodies to HIV-1). Fecal and serum specimens were collected 3 weeks later, at which time the individual was asymptomatic and antibodies directed to p24, p55, and gp120/gp160 could be measured by Western blotting (immunoblotting; HIV blot 2.2; Diagnostic Laboratories); therefore, individual C was clinically staged at group CDC class II. A second fecal specimen was collected 8 months later. Individuals D to R presented in the clinic, and 13 of them were diagnosed with AIDS. Two fecal and serum specimens were collected from individual D within 3 months. Eight individuals in the study suffered from diarrhea at the time of fecal sampling. An opportunistic infection could be identified in a majority of these individuals: cryptosporidial infection in three individuals (individuals E, K, and N), microsporidial infection in one individual (individual O), and a gastrointestinal infection by adenovirus in one person (individual F). No pathogen could be identified as causative agent of diarrhea in individuals H and R. Individual I suffered from a mild diarrhea at the time of fecal sampling, and tests for enteric pathogens were not performed. Sixteen participants were males infected by homosexual contact; individual M was a male infected by intravenous drug use, and individual N was a woman who became infected by heterosexual contact with an intravenous drug user. The detection of HIV-1 RNA in feces and serum was performed with the informed consent of all persons.
Reconstruction experiments.The serum of an HIV-1-infected person with a known quantity of virus was used as a control for the reconstruction experiments. The quantity of virus in this serum specimen was measured by limiting dilution of the serum in a nested gag reverse transcription PCR (RT-PCR) with SK39, SK145, SK431, and SK102 primers (26, 36). The quantification method was validated with a virus preparation in which the particle counts had been quan-tified directly by electron microscopy (31). The concentration of viral RNA in the serum which was used in the reconstruction was 13107
HIV-1 RNA molecules per ml of serum (53106
virions per ml of serum), and 10ml of a 10-fold dilution (containing 13104
HIV-1 RNA molecules) in sterile H2O was used to spike fecal specimens. To determine the efficiency of extraction of HIV-1 RNA from fecal specimens, 17 fecal samples of various colors and consistencies which were negative for HIV-1 RNA were used. One fecal specimen from an HIV-1-sero-negative person was used in an experiment to monitor the stability of HIV-1 RNA during repeated freezing-thawing. Approximately 30% (vol/vol) suspen-sions were made from fecal specimens by mixing them with broth (containing
nutrient broth no. 2 supplied by Oxoid, 500 IU of penicillin per ml, 500mg of streptomycin per ml, and 3mg of amphotericin B per ml). For one fecal specimen a 30% (vol/vol) suspension was made by mixing it with 40% glycerol. This suspension was used for the comparison of different extraction techniques. Glyc-erol was added to this fecal specimen to preserve bacterial integrity upon storage at2208C.
Storage of patient specimens.Fecal specimens collected in 1984 were sus-pended in broth and were stored at2708C. Fecal specimens collected in 1993 and 1994 were stored as lysates in two ways: (i) feces was mixed with broth (30% [vol/vol]; see above) and subsequently, 100ml of the suspension was added to 1,200ml of L6lysis buffer (7) or (ii) approximately 50 mg of feces was added directly to 1,200ml of L6lysis buffer. Lysates were stored at2208C. Serum samples were stored at2708C.
Extraction of RNA from feces and serum. (i) Protocol Y/SC.Isolation of nucleic acids from serum specimens was performed by protocol Y/SC (7). Briefly, 100ml of serum was added to a tube containing 900ml of L6lysis buffer (see below) and 40 ml of silica suspension. The tube was immediately vortexed. During an incubation of 10 min at room temperature, the RNA in the specimens was allowed to complex with the silica particles. The tubes were subsequently vortexed again and centrifuged for 15 s at 10,000 rpm (24-hole Hettich KG centrifuge), the supernatant was discarded, and the silica-nucleic acid complexes were washed twice with 1 ml of buffer L2(see below), twice with 1 ml of 70% (vol/vol) ethanol, and once with 1 ml of acetone and were dried at 568C (10 min); the nucleic acids were then eluted at 568C (10 min) in 60ml of sterile H2O. Twenty microliters of the eluate was used in an RT reaction, and 20ml of eluate was used for the negative control reaction without reverse transcriptase.
The preparation of size-fractionated silica, lysis buffer L6, and wash buffer L2 has been described in detail by Boom et al. (7). In short, lysis buffer L6was made by dissolving 120 g of GuSCN in 100 ml of 0.1 M TriszHCl (pH 6.4), and subsequently, 22 ml of 0.2 M EDTA (pH 8.0) and 2.6 g of Triton X-100 were added. Washing buffer L2was made by dissolving 120 g of GuSCN in 100 ml of 0.1 M TriszHCl (pH 6.4).
(ii) Protocol F/SC.Nucleic acids were extracted from the feces-broth mixtures by protocol F/SC, which is a slightly modified version of protocol Y/SC. In short, fecal specimens previously lysed in L6buffer and stored at2208C were thawed, or 50 to 100ml of the feces-broth mixture was added to 1,200ml of lysis buffer L6 and the mixture was vortexed vigorously. After 10 min at room temperature, the tubes were centrifuged (3 min at 10,000 rpm [24-hole Hettich KG centrifuge]). The resulting supernatant is free of particulate debris which would otherwise cosediment with silica-nucleic acid complexes when the standard protocol (pro-TABLE 1. Detection by RT-PCR of HIV-1 RNA in feces and serum of HIV-1-infected individuals
Individual Yr sample was obtained
RT-PCR resulta
CDC class Sex
b Risk groupc Antiviral
therapyd Diarrheae 100ml of feces 50ml of feces Serum
A 1984 1 1 1 II m Ho 2 2
1993 1 1 1 IVC1 2 2
B 1984 1 1 1 II m Ho 2 2
1993 1 1 1 III AZT1ddI 2
C May 1993 1 1 1 II m Ho 2 2
January 1994 1 1 1 II 2 2
D October 1993 1 2 1 IVC1 m Ho AZT 2
January 1994 1 1 1 IVC1 AZT 2
E 1993 1 1 1 IVC1 m Ho AZT 1
F 1993 1 1 1 II m Ho Proteaseinhf 1
G 1993 1g 1 1 IVC1 m Ho AZT1ddC 2
H 1993 1g 2 1 IVC1 m Ho AZT 1
I 1993 1g 1 1 IVC1 m Ho AZT1
3TC 1
J 1993 2 2 1 IVC1 m Ho 2 2
K 1993 2 2 1 IVC1 m Ho 2 1
L 1993 2 2 1 IVC1 m Ho ddC 2
M 1993 2 2 1 IVC1 m I 2 2
N 1993 1 1 1 IVC1 f He 2 1
O 1994 1 1 1 IVC1 m Ho 2 1
P 1993 2 2 1 II m Ho 2 2
Q 1994 2 2 1 IVC1 m Ho AZT1ddC 2
R 1994 1 2 1 IVA m Ho 2 1
aSee Materials and Methods.
bm and f, male and female, respectively.
cHo, He, and I, risk behaviors of homosexual contact, heterosexual contact, and intravenous drug use, respectively. dAZT, zidovudine; 3TC, 29,39-dideoxy-39thiacytidine.
eIndividuals who suffered from diarrhea are indicated with a plus sign.
fProteaseinh, protease inhibitor, which was supplied by Abbott and which was administered parenterally. gFifty milligrams of feces was used.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.612.61.555.83.328.2]tocol Y/SC) described above would have been used. Nine hundred microliters of the supernatant was then added to 40ml of silica suspension and was immediately vortexed, and the nucleic acids were allowed to bind during incubation for 10 min at room temperature. The tube was subsequently vortexed again and centrifuged for 15 s at 10,000 rpm (24-hole Hettich KG centrifuge). The supernatant was discarded and the silica-nucleic acid complexes were washed, dried, and eluted as described above for protocol Y/SC. In an isolated experiment an input of 1 ml of fecal suspension which was lysed in 9 ml of L6was used. Following centrifuga-tion, 9 ml of supernatant was added to 40ml of the silica suspension, and the tube was vortexed, incubated for 10 min at room temperature, and vortexed again. After centrifugation for 20 min at 3,000 rpm (Rotanta/AP Hettich centrifuge) the supernatant was discarded and the nucleic acids were purified as described above. In experiments in which the stability of rRNA during extraction was monitored 100ml of thawed feces-glycerol mixture lysed in 1 ml of L6was used. In these particular experiments nucleic acids were eluted in 80ml of TE con-taining an inhibitor of RNases (20 mM ribonucleoside-vanadyl complexes [RVCs]; Sigma).
(iii) Proteinase K-SDS procedure.One hundred microliters of a thawed feces-glycerol mixture was lysed by adding 100ml of lysis buffer containing 250 mM Tris (pH 7.5), 250 mM EDTA (pH 7.5), 1% sodium dodecyl sulfate (SDS), and 500mg of proteinase K (Sigma) per ml. After an incubation for 1 h at 568C, the samples were extracted twice with phenol-chloroform-isoamyl alcohol (25:24:1) and once with chloroform-isoamyl alcohol (24:1), and the nucleic acids were precipitated by adding 2.5 volumes of ethanol (100%) in the presence of 0.1 M NaCl. The nucleic acids were washed with 70% ethanol and were redissolved in 80ml of TE containing 20 mM RVCs.
RT.Annealing of the primer with template RNA was performed in a mixture containing 16 ng of antisense primer (L10, 59-GCGCCC-39; nucleotides 7831 to 7836 [HIV-1MN] [34]), 67 mM Tris (pH 8.8), 17 mM (NH4)2SO4, 1 mMb -mer-captoethanol, 6mM EDTA, 0.2 mg of bovine serum albumin (BSA) (Boehr-inger) per ml, 500mM (each) deoxynucleoside triphosphates (dNTPs), and 20 U of RNAsin (Promega) (39). After annealing for 3 min at room temperature, 200 U of Superscript (BRL GIBCO) and MgCl2(to a final concentration of 6 mM) were added and RT was performed for 1 h at 378C in a total volume of 40ml. Reverse transcriptase was denatured by incubation for 5 min at 958C (40), and RNase H treatment was performed for 30 min at 378C with 0.5 U of RNase H (Boehringer). As a control, reactions without reverse transcriptase were per-formed as described for the RT reaction but without the addition of Superscript and RNAsin and without the RNase H treatment.
PCR.Twenty microliters of cDNA was added to 80ml of the PCR mixture containing 100 ng of antisense primer (L9, 59-CCCAAGGAACAAAGCTCC-39; nucleotides 7787 to 7804 [HIV-1MN]), 100 ng of sense primer (59 V3NOT; 59-GCGCGGCCGCACAGTACAATGTACACATGG-39; nucleotides 6983 to 7002 [HIV-1MN]), 62.5 mM Tris (pH 8.3), 25 mM KCl, 0.125 mg of BSA per ml, 125mM (each) dNTPs, 1.25 mM MgCl2, and 2 U of Taq polymerase (Perkin-Elmer Cetus). After incubation for 5 min at 948C the sample was subjected to 35 cycles of amplification in a DNA thermal cycler (type 480; Perkin-Elmer Cetus). A cycle consisted of denaturation for 15 s at 948C, annealing for 45 s at 558C, and extension for 1 min at 728C. After the cycling program the samples were incu-bated for 10 min at 728C. Five microliters of the first PCR mixture was amplified in a nested PCR of 30 cycles in 100ml containing 100 ng of antisense primer (39V3NOT; 59-GCGCGGCCGCCCCCTCTACAATTAAAACTGTG-39; nucle-otides 7367 to 7388 [HIV-1MN]), 100 ng of sense primer (59KSI, 59-ATAAGCT TGCAGTCTAGCAGAAGAAGA-39; nucleotides 7041 to 7059 [HIV-1MN]), 50 mM Tris (pH 8.3), 20 mM KCl, 0.1 mg of BSA per ml, 100mM (each) dNTPs, 2 mM MgCl2, and 2 U of Taq polymerase (Perkin-Elmer Cetus). The same cycle program used for the first PCR was performed. In reconstruction experiments a single PCR was used, and 37 cycles of amplification were performed. Ten mi-croliters of the amplified product was analyzed by agarose gel electrophoresis. PCR products derived from the stability and efficiency tests (a single PCR) were detected after Southern blotting by hybridization with a32P-labeled probe made from a plasmid containing the entire gp160 of HIV-1MN(plasmid KL-8; see below). The PCR product of the nested PCR was detected by ethidium bromide staining of an agarose gel. Negative and positive extraction controls were in-cluded in each experiment. As a positive control, 105HIV-1 RNA molecules isolated from a virus culture were used (31).
To construct the gp160-containing plasmid, a PCR was performed on chro-mosomal DNA containing HIV-1MNproviral DNA with the primers VIF-N (59-GATGCGGCCGCTCGCGATTATGGAAAACAGANGGCAGGTG ATGATTGTG 39; nucleotides 5054 to 5085 [HIV-1MN]) and 39END-N (59-GA GGCGGCCGCGGCAAGCTTTATTGAGGCTTAAG 39; nucleotides 9621 to 9643 [HIV-1MN]). This PCR product was cloned into plasmid PSP73 (Promega) and was subsequently subcloned in PSP64polyA (Promega). The final plasmid, KL-8, contained gp160 of HIV-1MNfrom nucleotides 5758 to 8917. Underlining in the primers indicates endonuclease restriction sites.
Sequencing.Ten microliters of the nested PCR product was purified from a neutral agarose gel by dissolving approximately 50ml of a gel slice in 900ml of lysis buffer L7T (containing 1 mg of casein [purified from bovine milk; catalog no. C5890 Sigma]) per ml of L6) and 2.5ml of the silica suspension. The tubes were vortexed every 10 min during incubation for 40 min at 378C. DNA-silica com-plexes were washed twice with 1 ml of L2, twice with 1 ml of 70% ethanol, and once with 1 ml of acetone. After drying at 568C for 10 min the purified DNA was
eluted at 568C (10 min) in 15ml of TE, and 6ml of purified DNA was used in a sequence reaction. Both strands were sequenced by using the sense primer 59PVUII-J (59-GTACAGCTGAATGAATCTGTAGAAATTAATTGT-39; nu-cleotides 7102 to 7126 [HIV-1MN]) and the antisense primer 39V3NOT. In the presence of one of the sequence primers the PCR product was denatured by incubating for 3 min at 1008C; this was followed by immediate freezing at2788C in dry ice with ethanol. Dideoxynucleotide sequencing was performed by using Sequenase 2.0 (U.S. Biochemicals) by applying the following modifications to the manufacturer’s protocol. Labeling reactions with [35S]dATP and termination reactions were performed simultaneously at 378C for 5 min in the presence of 10% dimethyl sulfoxide.
Nucleotide sequence accession numbers.The sequences presented in this report have been submitted to GenBank (accession no. L38720-L38733).
RESULTS
Adaptation of standard protocol Y/SC for feces.
A serious
drawback for the isolation of RNA from feces is the presence
of very high concentrations of RNases. To address this issue, a
recently described nucleic acid extraction technique was used
(7). The technique efficiently inactivates RNases during lysis of
the sample (protocol Y/SC). However, to isolate nucleic acids
from crude feces or feces-broth mixtures an adaptation of the
standard procedure for extraction appeared to be necessary,
since preliminary experiments showed that debris which
co-sedimented with silica-nucleic acid complexes caused RNA
degradation (data not shown). To overcome this problem we
included a centrifugation step which removed debris from the
lysate prior to binding of nucleic acids to silica particles
(pro-tocol F/SC).
Inactivation of RNases during nucleic acid extraction.
RNA
isolation from feces by protocol F/SC was compared with the
isolation of RNA by a classic method that uses proteinase
K-SDS-mediated lysis and several extractions with organic
sol-vents. The integrity of bacterial rRNA was used as a marker for
the inactivation of RNases in the different purification
proce-dures. The nucleic acids isolated by the two different methods
were examined by agarose gel electrophoresis and ethidium
bromide staining. In Fig. 1 the degradation of rRNA extracted
from feces by the classic procedure is apparent, while the
rRNA isolated by protocol F/SC remained intact.
Efficiency of RNA extraction and RT-PCR from feces.
The
influence of the composition of feces on the extraction and the
detection technique was monitored for 17 different fecal
spec-imens. Virions (5
3
10
3) were added to 50
m
l of feces-broth
[image:3.612.352.517.70.190.2]mixtures, and the nucleic acids were purified by protocol F/SC.
FIG. 1. Purified nucleic acids from feces isolated by different extraction tech-niques. Thirty microliters of nucleic acids which were purified from a feces-glycerol suspension was electrophoresed through a 0.8% agarose gel containing ethidium bromide, and the gel was subsequently photographed under UV illu-mination. Lanes 1 and 7, 2mg of phage lambda DNA digested with HindIII; lanes 2 and 8, 2mg of Escherichia coli rRNA; lanes 3 and 4, nucleic acids purified by the proteinase K-SDS procedure; lanes 5 and 6, nucleic acids purified by protocol F/SC. The photograph was scanned by using a Scanjet IIC scanner (Hewlett-Packard), the layout was done with MacDrawPro 1.5 (Claris Corporation), and the picture was printed on a Kodak Colorease PS printer.
on May 15, 2020 by guest
http://jcm.asm.org/
In parallel, nucleic acids were purified from positive (5
3
10
3virions in H
2O) and negative (no HIV added) samples.
Fol-lowing extraction, HIV-1 RNA was amplified by a single
RT-PCR and the RT-PCR products were analyzed by Southern blot
hybridization (Fig. 2). In all specimens the input HIV-1 RNA
was amplified, albeit to a different extent. The variation
be-tween duplicates could have been due to the small number of
spiked virions, which was close to the lower limit of detection
(see below). A comparable experiment was performed on
seven fecal samples with an input of 100
m
l of feces-broth
suspension. Again, from all fecal specimens the spiked input of
5
3
10
3virions could be amplified in a single PCR (data not
shown).
The lower limit of detection was determined for two fecal
specimens (specimens F9 and F12 in Fig. 2). To determine
whether the variability of the PCR signal as shown in Fig. 2 was
the result of inefficient amplification in a single PCR or
inef-ficient extraction, fecal specimen F9 was included in the
ex-periment since specimen F9 clearly exhibited a diminished
signal in the single PCR. Twofold serial dilutions of virus were
added to 50
m
l of the feces-broth mixtures of specimens F9 and
F12. Nucleic acids were extracted by protocol F/SC, and the
detection level was determined in two ways: in a single PCR, in
conjunction with detection by Southern blot hybridization, and
in a nested PCR, with product detection by ethidium bromide
agarose gel electrophoresis. The lowest concentration of virus
which could be amplified and monitored was equal for a single
PCR and a nested PCR and was 300 virions (600 HIV-1 RNA
molecules) in both fecal specimens (data not shown). The
detection level turned out to be the same for both fecal
spec-imens, indicating that the variability seen in a single PCR (Fig.
2) is most likely the result of a feces-dependent variable
am-plification efficiency in a single PCR and not an effect of
inef-ficient extraction of the viral RNA.
Stability of HIV-1 RNA in feces upon freezing-thawing.
The
stability of virus during repeated freezing-thawing was
moni-tored in a reconstruction experiment for one fecal specimen.
To 600
m
l of feces-broth mixture, 3
3
10
4virions were added.
This mixture was freeze-thawed four times. One freeze-thaw
cycle included contact for 5 min with dry ice (
2
78
8
C) and
incubation at room temperature until the specimen was
thawed. Before each cycle of freezing, 100
m
l of the mixture
was added to 1 ml of L
6. After extraction of nucleic acids and
amplification by a single RT-PCR the amplified products were
detected by Southern blot hybridization. The PCR signals
ob-tained after each freezing-thawing cycle are shown in Fig. 3.
The signal obtained after four cycles of freezing-thawing was
equal to the signal obtained with the samples which were not
frozen, indicating that RNA molecules within virions were not
degraded during repeated freezing-thawing.
Detection of HIV-1 RNA in feces of HIV-1-infected
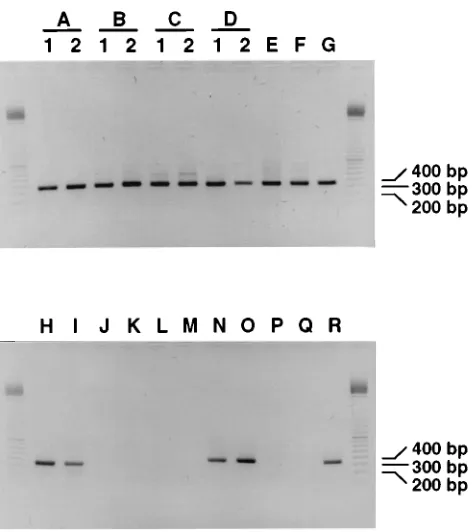
individ-uals.
Twenty-two fecal specimens from 18 HIV-1-infected
in-dividuals were tested for the presence of HIV-1 RNA. In 1993
to 1994 three of the individuals fulfilled the criteria for CDC
class II, one fulfilled the criteria for CDC class III, and 14 were
diagnosed with AIDS (Table 1). HIV-1 RNA could be
de-tected in the feces of 12 of the 18 individuals (67%) with an
input of either 100
m
l of a fecal suspension or approximately 50
mg of feces (Table 1 and Fig. 4). HIV-1 RNA was detected in
the feces of two of the three persons belonging to CDC class II
(individuals C and F), among which was one individual who
had recently been infected. HIV-1 RNA was also detected in
the feces of the one person (individual B) who was clinically
classified in CDC class III. Furthermore, HIV-1 RNA was
present in the feces of 9 of the 14 individuals who were
diag-nosed with AIDS (individuals A, D, E, G, H, I, N, O, and R).
Two fecal specimens were collected from four individuals at
different times. A second fecal specimen was collected 8
months later from the person whose fecal specimen was
ob-tained a few weeks after primary infection (individual C). In
this specimen HIV-1 RNA was again detectable. Furthermore,
a second fecal specimen from one of the persons with AIDS
(individual D), which was collected 3 months after the first
specimen was obtained, was again positive for HIV-1 RNA.
Fecal samples had also been collected from two individuals
(individuals A and B) 9 years earlier. At that time both persons
were asymptomatic and fulfilled the criteria for CDC class II.
[image:4.612.74.282.69.341.2]FIG. 2. Efficiency of HIV-1 RNA extraction and amplification. Duplicate extractions and amplifications were performed with 17 fecal specimens spiked with virions and control samples without feces. Ten microliters of the product from the single PCR was electrophoresed through a 1% agarose gel, blotted onto nitrocellulose, and hybridized with a 32P-labeled gp160-containing plasmid probe. Lanes C1 to C4, amplified control input HIV-1 RNA; lanes f1 to f17, amplified HIV-1 RNA extracted from 17 different fecal specimens; lane N1, negative extraction control in the extraction series containing C1, C2, and f1 to f8; lane N2, negative extraction control in the extraction series containing C3, C4, and f9 to f17. The picture was scanned as described in the legend to Fig. 1.
FIG. 3. Stability of virions during freezing-thawing. A fecal suspension was spiked with virions and freeze-thawed several times, and nucleic acids were extracted and amplified in a single PCR. Ten microliters of PCR product was electrophoresed on a 1% agarose gel, blotted onto nitrocellulose, and hybridized with a32
P-labeled gp160-containing plasmid probe. Lane 1, PCR product with-out freezing-thawing of the sample; lanes 2 to 5, PCR product after one to four cycles of freezing-thawing, respectively; lane 6, PCR product of spiked fecal suspension which was incubated at room temperature for 1 h, lane 7, negative extraction control. The picture was scanned as described in the legend to Fig. 1.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.612.359.510.70.133.2]HIV-1 RNA could also be extracted and amplified from these
early specimens. For all fecal specimens the reaction without
reverse transcriptase was negative, suggesting that the detected
signals originated from HIV-1 RNA rather than from proviral
DNA (data not shown).
With an input of 50
m
l of a fecal suspension, HIV-1 RNA
could be detected in 9 of the 18 (50%) HIV-1-infected
indi-viduals (Table 1), indicating that in three persons the amount
of HIV-1 RNA in feces was close to the detection level. HIV-1
RNA could not be detected in feces from six HIV-1-infected
individuals, while HIV-1 RNA was detectable in their sera
(Table 1). This indicates that the primers used in the RT-PCR
bound to the viral RNA extracted from the sera of those
persons. However, to ensure that inefficient annealing of the
envelope primers to the viral RNA extracted from feces was
not the cause of the negative results, a nested gag RT-PCR
with primers SK39, SK145, SK431, and SK102 (26, 36) was also
performed on nucleic acids extracted from the feces of the six
individuals. The gag RT-PCR was not positive for any of the
specimens (data not shown). In reconstruction experiments in
which these six fecal specimens were spiked with virions (5
3
10
3), this input was efficiently purified and amplified by
RT-PCR (data not shown).
Negative and positive controls accompanied the clinical
specimens in the extractions. In none of the experiments did
these controls test either false positive or false negative.
Direct sequencing.
The specificities of the PCR products
from feces and serum were confirmed by direct sequencing of
the amplimers. From 16 fecal specimens from the 12 persons in
whose feces HIV-1 RNA was detectable, HIV-1 sequences
confirmed the specificity of the nested RT-PCR product (data
not shown). A comparison of the HIV-1 sequences obtained
from eight paired fecal and serum specimens by direct
se-quencing revealed that the subpopulation of HIV-1 found in
feces can differ from the HIV-1 subpopulation found in serum.
In most cases the HIV-1 RNA sequence of virus from feces
appeared to be more homogeneous than the sequence of virus
from serum (individuals A [1984 and 1993], B [1984 and 1993],
G, and H; Fig. 5B and 6). Although the sequence of virus from
the feces of individual A (1993) was more homogeneous when
the entire sequence of the V3 region was compared, there were
some positions at which the sequence of virus from feces
dis-played heterogeneity whereas the sequence of virus from
se-rum did not (Fig. 5A and 6). Sequences of virus from fecal and
serum specimens which were obtained shortly after primary
infection of individual C were identical (data not shown).
Re-markably, the sequences of HIV-1 from feces and serum of
individual I obtained by direct sequencing were clearly distinct
(Fig. 5C). Besides a codon deletion, some positions displayed
different nucleotides. These nucleotide substitutions resulted
in a different deduced amino acid sequence of the V3 loop
(Fig. 7). Since we analyzed both the sequences and the
heter-ogeneities of the sequences of HIV-1 from feces and serum by
direct sequencing of the PCR products, the reproducibility of
direct sequencing was crucial. Therefore, duplicate extractions,
amplifications, and direct sequencing of two serum samples
was performed. Of the 220 nucleotides that were sequenced, a
difference in heterogeneity was measured between the two
duplicate sequences at only a single position, which was a very
small percentage of the total heterogeneous nucleotide
posi-tions (3.2%) (data not shown). Furthermore, the effect of the
input amount on the heterogeneity in a sequence determined
by direct sequencing was examined. HIV-1 RNA was extracted
from 1-ml and 100-
m
l fecal suspensions of individual B (1993)
and was then amplified. The sequence with 10 times more
input did not result in a substantial change in the heterogeneity
(0.9%) of the sequence (data not shown).
DISCUSSION
We have presented a rapid and simple method (protocol
F/SC) for the purification of HIV-1 RNA from feces which is
unique in its simplicity. HIV-1 RNA can easily be purified from
large series of fecal specimens (12 specimens in about 1.5 h)
and has appeared to be a good substrate in RT-PCR. The
variability in the composition of fecal specimens did not seem
to play a major role in the extraction of HIV-1 RNA. A nested
RT-PCR instead of Southern blot hybridization of a single
PCR was chosen for the detection of HIV-1 RNA because it is
more rapid and the amplimers are produced in such quantities
that they can be used for direct sequencing.
HIV-1 RNA has previously been detected in feces from
HIV-1-infected children (49). Our data extend these
observa-tions to HIV-1-infected adults. We could detect HIV-1 RNA
in the feces of the majority (67%) of our study group. Among
the individuals in whose feces HIV-1 RNA was detectable
were individuals who were clinically staged in CDC classes II,
III, and IV. So the detection of HIV-1 RNA was not restricted
to those with a certain stage of HIV-induced disease.
In theory, the HIV-1 RNA detected in feces could be either
RNA in HIV-1-infected cells or genomic HIV-1 RNA within
viral particles. The HIV-1 RNA detected in feces probably
originated from virions, because we did detect HIV-1 RNA in
feces which had been stored for 9 years at
2
70
8
C. It is unlikely
[image:5.612.61.299.68.334.2]that cells could have survived these storage conditions, and
FIG. 4. Compilation of different experiments showing nested PCR signals of feces from HIV-1-infected individuals A to R. Ten microliters of nested PCR products from different experiments were electrophoresed on a 1% agarose gel containing ethidium bromide and were subsequently photographed under UV illumination. Lanes A1 to D2, feces collected at the first (lanes 1) and second (lanes 2) time points from individuals A to D; lanes E to R, feces from individuals E to R, respectively. Lanes on the left and right sides of the PCR products contain a 100-bp ladder (Pharmacia), the 200-, 300-, and 400-bp fragments of which are indicated. The picture was scanned as described in the legend to Fig. 1.
on May 15, 2020 by guest
http://jcm.asm.org/
mRNA would probably have been degraded. Moreover, we did
not detect HIV-1 proviral DNA in feces, which also suggests
that HIV-1-infected cells were not present. However, we did
not determine the efficiency of our technique for the extraction
of DNA from feces.
[image:6.612.94.514.72.283.2]It is possible that the presence of HIV-1 RNA in feces
reflects the production of virions by epithelial cells that excrete
viral particles by apical budding, as postulated by Yahi et al.
(48). It is also feasible that HIV-1 produced by lymphocytes or
macrophages in the lamina propria can pass the epithelial
FIG. 5. Differences in sequences of HIV-1 RT-PCR products from paired feces and serum of HIV-1-infected individuals. Autoradiograms are of sequences of virus from feces (F) and serum (S) obtained by direct sequencing. (A) Specimens from individual A (1993); grey arrows indicate nucleotide positions where the sequence is heterogeneous in feces and homogeneous in serum; black arrows indicate positions where the serum sequence is heterogeneous and the feces sequence is homogeneous. (B) Specimens from individual B (1993); black arrows indicate positions where the serum sequence is heterogeneous and the feces sequence is homogeneous. (C) Specimens from individual I. The codon deletion in the sequence of virus from feces in comparison with that in the sequence of virus from serum is indicated. Other major sequence differences are presented in Fig. 7. Numbering is as described previously for HIV-1MN(34). The picture was scanned as described in the legend to Fig. 1.
FIG. 6. Direct nucleotide sequences of HIV-1 RNA in serum and feces of individuals A (1984 and 1993), B (1984 and 1993), G, and H. Part of the V3 region is shown; dashes indicate identity. Heterogeneous nucleotide positions are indicated by R (A and G), Y (C and T), M (A and C), S (G and C), W (A and T), D (G, A, and T), and V (G, A, and C);p, the sequence of virus from serum is heterogeneous, while the sequence of virus from feces is homogeneous at this position;F, the sequence of virus from serum is homogeneous, while the sequence of virus from feces is heterogeneous at this position; #, sites where sequences of virus from serum and feces display different nucleotides. Numbering is as described previously for HIV-1MN(34).
on May 15, 2020 by guest
http://jcm.asm.org/
[image:6.612.53.557.438.684.2]barrier because of the increased permeability of the epithelial
cell layer (1, 4, 16, 21, 27). Future research on the
character-ization of variants in different cell types in the intestinal
mu-cosa is needed to determine the origins of HIV-1 particles in
feces.
We can rigorously exclude the possibility that the high
per-centage of HIV-1 RNA-positive fecal specimens was the result
of aspecific amplification or contamination in the PCR. In all
experiments negative controls were added in the extraction,
and they never turned out to be positive. Furthermore, in all
amplifications from fecal and serum specimens the RT-PCR
performed without reverse transcriptase was negative,
indicat-ing not only that the PCR signal originated from RNA but also
that no contamination by PCR products or HIV-containing
plasmids had occurred. Furthermore, the PCR products of the
fecal specimens were sequenced, and this excluded the
possi-bility of intersample contamination.
The nucleotide sequences showed a more homogeneous
subpopulation of HIV-1 in feces in comparison with that in
serum for a majority of the individuals studied. In principle,
this decreased level of heterogeneity could be caused by the
amplification of only a few molecules of HIV-1 RNA.
How-ever, in the one case studied a 10-fold increase in the amount
of feces input did not result in a significant difference in the
heterogeneity of the direct sequence.
At some positions in the HIV-1 RNA from feces the
nucle-otides displayed heterogeneity which was not detectable in
serum, suggesting that a distinct HIV-1 subpopulation is
present in feces. Moreover, in one case a clear-cut difference
between the sequences of virus from feces and serum obtained
by direct sequencing was observed. It is therefore unlikely that
the presence of viral RNA in feces is solely the result of
leakage from blood. Our characterization of HIV-1 variants in
feces by direct sequencing confirms the findings of Barnett et
al. (2), who showed that HIV isolated from the bowel can differ
in biological and serological properties from HIV strains in
blood. The cell tropism of HIV-1 is dependent on the amino
acid sequence of the envelope (8, 10, 13, 19, 41), among which
is the V3 region (24, 42, 43, 47). The direct sequence of the V3
region of virus from individual I showed a deduced amino acid
sequence which differed substantially from that of virus in
serum. Therefore, it could be that the viruses detected in the
feces of individual I are produced in a cell type different from
the cells in the blood that produce virus.
In the study described here a relatively small group of
HIV-1-infected patients was studied; therefore, a correlation
be-tween gastrointestinal symptoms and the detection of HIV-1
RNA cannot be made. In addition, most individuals in our
study group were males infected by homosexual contact, and
the chance that they were infected by passive anal sexual
in-tercourse is great. Future research will reveal whether there is
a link between the route of HIV-1 transmission and the
pres-ence of HIV-1 in the intestinal lumen and whether there is a
link between the onset of gastrointestinal symptoms and the
detection of HIV-1 RNA in feces.
ACKNOWLEDGMENTS
We thank Wim van Est for expert technical assistance, J. Veenstra for providing fecal specimens, M. Cornelissen for quantifying HIV-1 RNA in serum, A. van Strien for preparing the buffers for nucleic acid extraction, and P. Nara, Virus Biology Unit, National Cancer Institute, Frederick, Md., for providing an HXB3 virus stock.
REFERENCES
1. Asmuth, D. M., S. M. Hammer, and C. A. Wanke. 1994. Physiological effects of HIV infection on human intestinal epithelial cells: an in vitro model for HIV enteropathy. AIDS 8:205–211.
2. Barnett, S. W., A. Barboza, C. M. Wilcox, C. E. Forsmark, and J. A. Levy. 1991. Characterization of human immunodeficiency virus type 1 strains re-covered from the bowel of infected individuals. Virology 182:802–809. 3. Bartlett, J. G., P. C. Belitsos, and C. L. Sears. 1992. AIDS enteropathy. Clin.
Infect. Dis. 15:726–735.
4. Batman, P. A., S. C. Fleming, P. M. Sedgwick, T. T. MacDonald, and G. E.
Griffin.1994. HIV infection of human fetal intestinal explant cultures in-duces epithelial cell proliferation. AIDS 8:161–167.
5. Batman, P. A., A. R. O. Miller, S. M. Forster, J. R. W. Harris, A. J. Pinching,
and G. E. Griffin.1989. Jejunal enteropathy associated with human immu-nodeficiency virus infection: quantitative histology. J. Clin. Pathol. 42:275– 281.
6. Boom, R., C. J. A. Sol, R. Heijtink, P. M. E. Wertheim-van Dillen, and J. van
der Noordaa.1991. Rapid purification of hepatitis B virus DNA from serum. J. Clin. Microbiol. 29:1804–1811.
7. Boom, R., C. J. A. Sol, M. M. M. Salimans, C. L. Jansen, P. M. E.
Wertheim-van Dillen, and J. Wertheim-van der Noordaa.1990. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 28:495–503.
8. Cann, A. J., M. J. Churcher, M. Boyd, W. O’Brien, J.-Q. Zhao, J. Zack, and
I. S. Y. Chen.1992. The region of the envelope gene of human immunode-ficiency virus type 1 responsible for determination of cell tropism. J. Virol.
66:305–309.
9. Centers for Disease Control. 1987. Revision of the CDC surveillance case definition of acquired immunodeficiency syndrome. Morbid. Mortal. Weekly Rep. 36:3s–15s.
10. Cheng-Mayer, C., T. Shioda, and J. Levy. 1991. Host range, replicative, and cytopathic properties of human immunodeficiency virus type 1 are deter-mined by very few amino acid changes in tat and gp120. J. Virol. 65:6931– 6941.
11. Clayton, F., S. Reka, W. J. Cronin, E. Torlakovic, S. H. Sigal, and D. P.
Kotler.1992. Rectal mucosal pathology varies with human immunodefi-ciency virus antigen content and disease stage. Gastroenterology 103:919– 933.
12. Connolly, G. M., D. Shanson, D. A. Hawkins, J. N. Harcourt Webster, and
B. G. Gazzard.1989. Non-cryptosporidial diarrhoea in human immunodefi-ciency virus (HIV) infected patients. Gut 30:195–200.
13. Cordonnier, A., L. Montagnier, and M. Emerman. 1989. Single amino-acid changes in HIV envelope affect viral tropism and receptor binding. Nature (London) 340:571–574.
14. de Wolf, F., J. M. A. Lange, J. T. M. Houweling, R. A. Coutinho, P. Th. A.
Schellekens, J. van der Noordaa, and J. Goudsmit.1988. Numbers of CD41 cells and the levels of core antigens of and antibodies to the human immu-nodeficiency virus as predictors of AIDS among seropositive homosexual men. J. Infect. Dis. 158:615–622.
15. Dworkin, B., G. P. Wormser, W. S. Rosenthal, S. K. Heier, M. Braunstein,
L. Weiss, R. Jankowski, D. Levy, and S. Weiselberg.1985. Gastrointestinal FIG. 7. Sequences obtained by direct nucleotide sequencing (A) and deduced amino acid sequence (B) of HIV-1 RNA in serum and feces of individual I. Part of the V3 region is shown; dashes indicate identity, and dots indicate deletions. Numbering is as described previously for HIV-1MN(34).
on May 15, 2020 by guest
http://jcm.asm.org/
manifestations of the acquired immunodeficiency syndrome: a review of 22 cases. Am. J. Gastroenterol. 80:774–778.
16. Fantini, J., N. Yahi, S. Baghdiguian, and J.-C. Chermann. 1992. Human colon epithelial cells productively infected with human immunodeficiency virus show impaired differentiation and altered secretion. J. Virol. 66:580– 585.
17. Fantini, J., N. Yahi, and J.-C. Chermann. 1991. Human immunodeficiency virus can infect the apical and basolateral surfaces of human colonic epithe-lial cells. Proc. Natl. Acad. Sci. USA 88:9297–9301.
18. Fox, C. H., D. Kotler, A. Tierney, C. S. Wilson, and A. S. Fauci. 1989. Detection of HIV-1 RNA in the lamina propria of patients with AIDS and gastrointestinal disease. J. Infect. Dis. 159:467–471.
19. Fujita, K., J. Silver, and K. Peden. 1992. Changes in both gp120 and gp41 can account for increased growth potential and expanded host range of human immunodeficiency virus type 1. J. Virol. 66:4445–4451.
20. Gazzard, B. G. 1992. Diarrhea in human immunodeficiency virus antibody-positive patients. Semin. Liver Dis. 12:154–166.
21. Gillin, J. S., M. Shike, N. Alcock, C. Urmacher, S. Krown, R. C. Kurtz, C. J.
Lightdale, and S. J. Winawer.1985. Malabsorption and mucosal abnormal-ities of the small intestine in the acquired immunodeficiency syndrome. Ann. Intern. Med. 102:619–622.
22. Griffin, G. E. 1990. Malabsorption, malnutrition and HIV disease, p. 361– 373. In B. G. Gazzard (ed.), Gastroenterological aspects of AIDS, Baillie`re’s clinical gastroenterology. Baillie`re Tindall, London.
23. Heise, C., S. Dandekar, P. Kumar, R. Duplantier, R. M. Donovan, and C. H.
Halsted.1991. Human immunodeficiency virus infection of enterocytes and mononuclear cells in human jejunal mucosa. Gastroenterology 100:1521– 1527.
24. Hwang, S. S., T. J. Boyle, H. K. Lyerly, and B. R. Cullen. 1991. Identification of the envelope V3 loop as the primary determinant of cell tropism in HIV-1. Science 253:71–74.
25. Jarry, A., A. Cortez, E. Rene´, F. Muzeau, and N. Brousse. 1990. Infected cells and immune cells in the gastrointestinal tract of AIDS patients. An immu-nohistochemical study of 127 cases. Histopathology 16:133–140.
26. Kellogg, D. E., and S. Kwok. 1990. Detection of human immunodeficiency virus, p. 337–347. In M. A. Innes, D. H. Gelfand, J. J. Sninsky, and T. J. White (ed.), PCR protocols: a guide to methods and applications. Academic Press, Inc., San Diego, Calif.
27. Kotler, D. P., H. P. Gaetz, M. Lange, E. B. Klein, and P. R. Holt. 1984. Enteropathy associated with the acquired immunodeficiency syndrome. Ann. Intern. Med. 101:421–428.
28. Kotler, D. P., S. Reka, A. Borcich, and W. J. Cronin. 1991. Detection, localization and quantitation of HIV-associated antigens in intestinal biop-sies from patients with HIV. Am. J. Pathol. 139:823–830.
29. Kotler, D. P., S. Reka, and F. Clayton. 1993. Intestinal mucosal inflammation associated with human immunodeficiency virus infection. Dig. Dis. Sci. 38: 1119–1127.
30. Laughon, B. E., D. A. Druckman, A. Vernon, T. C. Quinn, B. F. Polk, J. F.
Modlin, R. H. Yolken, and J. G. Bartlett.1988. Prevalence of enteric patho-gens in homosexual men with and without acquired immunodeficiency syn-drome. Gastroenterology 94:984–993.
31. Layne, S. P., M. J. Merges, M. Dembo, J. L. Spouge, S. R. Conley, J. P.
Moore, J. L. Raina, H. Renz, H. R. Gelderblom, and P. L. Nara.1992. Factors underlying spontaneous inactivation and susceptibility to neutraliza-tion of human immunodeficiency virus. Virology 189:695–714.
32. Levy, J. A., W. Margaretten, and J. Nelson. 1989. Detection of HIV in enterochromaffin cells in the rectal mucosa of an AIDS patient. Am. J.
Gastroenterol. 84:787–789.
33. Mathijs, J. M., M. Hing, J. Grierson, D. E. Dwyer, C. Goldschmidt, D. A.
Cooper, and A. L. Cunningham.1988. HIV infection of rectal mucosa. Lancet i:1111.
34. Myers, G., A. B. Rabson, J. A. Berzofsky, T. F. Smith, and F. Wong-Staal. 1990. Human retroviruses and AIDS. A compilation and analysis of nucleic and amino acid sequences. In Theoretical biology and biophysics. Los Alamos National Laboratory, Los Alamos, N.M.
35. Nelson, J. A., C. Reynolds-Kohler, W. Margaretten, C. A. Wiley, C. E. Reese,
and J. A. Levy.1988. Human immunodeficiency virus detected in bowel epithelium from patients with gastrointestinal symptoms. Lancet i:259–262. 36. Portegies, P. 1993. Ph.D. thesis. University of Amsterdam, Amsterdam. 37. Riecken, E. O., M. Zeitz, and R. Ullrich. 1990. Non-opportunistic causes of
diarrhoea in HIV infection, p. 385–403. In B. G. Gazzard (ed.), Gastroen-terological aspects of AIDS, Baillie`re’s clinical gastroenterology. Baillie`re Tindall, London.
38. Rodgers, V. D., R. Fassett, and M. F. Kagnoff. 1986. Abnormalities in intestinal mucosal T cells in homosexual populations including those with the lymphadenopathy syndrome and acquired immunodeficiency syndrome. Gastroenterology 90:552–558.
39. Sellner, L. N., R. J. Coelen, and J. S. Mackenzie. 1992. A one-tube, one manipulation RT-PCR for detection of Ross River virus. J. Virol. Methods
40:255–264.
40. Sellner, L. N., R. J. Coelen, and J. S. Mackenzie. 1992. Reverse transcriptase inhibits Taq polymerase activity. Nucleic Acids Res. 20:1487–1490. 41. Shioda, T., J. A. Levy, and C. Cheng-Mayer. 1991. Macrophage and T
cell-line tropisms of HIV-1 are determined by specific regions of the enve-lope gp120 gene. Nature (London) 349:167–169.
42. Shioda, T., J. A. Levy, and C. Cheng-Mayer. 1992. Small amino acid changes in the V3 hypervariable region of gp120 can affect the T-cell-line and mac-rophage tropism of human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 89:9434–9438.
43. Takeuchi, Y., M. Akutsu, K. Murayama, N. Shimizu, and H. Hoshino. 1991. Host range mutant of human immunodeficiency virus type 1: modification of cell tropism by a single point mutation at the neutralization epitope in the env gene. J. Virol. 65:1710–1718.
44. Ullrich, R., M. Zeitz, W. Heise, M. L’age, G. Ho¨ffken, and E. O. Riecken.
1989. Small intestinal structure and function in patients infected with human immunodeficiency virus (HIV): evidence for HIV-induced enteropathy. Ann. Intern. Med. 111:15–21.
45. Ullrich, R., M. Zeitz, W. Heise, M. L’age, K. Ziegler, C. Bergs, and E.-O.
Riecken.1990. Mucosal atrophy is associated with loss of activated T cells in the duodenal mucosa of human immunodeficiency virus (HIV)-infected pa-tients. Digestion 46(Suppl. 2):302–307.
46. Ullrich, R., M. Zeitz, and E.-O. Riecken. 1992. Enteric immunologic abnor-malities in human immunodeficiency virus infection. Semin. Liver Dis. 12: 167–174.
47. Westervelt, P., D. B. Trowbridge, L. G. Epstein, B. M. Blumberg, Y. Li, B. H.
Hahn, G. M. Shaw, R. W. Price, and L. Ratner.1992. Macrophage tropism determinants of human immunodeficiency virus type 1 in vivo. J. Virol.
66:2577–2582.
48. Yahi, N., S. Baghdiguian, C. Bolmont, and J. Fantini. 1992. Replication and apical budding of HIV-1 in mucous-secreting colonic epithelial cells. J. Acquired Immune Defic. Syndr. 5:993–1000.
49. Yolken, R. H., S. Li, J. Perman, and R. Viscidi. 1991. Persistent diarrhea and fecal shedding of retroviral nucleic acids in children infected with human immunodeficiency virus. J. Infect. Dis. 164:61–66.