Evaluation of Vitek MS for Differentiation of
Cryptococcus
neoformans
and
Cryptococcus gattii
Genotypes
Lumena P. Machado Siqueira,
aViviane M. Favero Gimenes,
aRoseli Santos de Freitas,
aMárcia de Souza Carvalho Melhem,
bLucas Xavier Bonfietti,
bAfonso Rafael da Silva, Jr.,
cLetícia B. Souza Santos,
aAdriana L. Motta,
cFlavia Rossi,
cGil Benard,
aJoão N. de Almeida, Jr.
a,caLaboratorio de Micologia Médica (LIM 53), Instituto de Medicina Tropical, Universidade de São Paulo, São Paulo, Brazil bDivisão Técnica de Biologia Médica, Seção de Micologia, Instituto Adolfo Lutz, São Paulo, Brazil
cLaboratório Central (LIM 03), Hospital das Clínicas da Faculdade de Medicina da Universidade de São Paulo, São Paulo, Brazil
ABSTRACT
Cryptococcus neoformans
and
Cryptococcus gattii
are the main
patho-genic species of invasive cryptococcosis among the
Cryptococcus
species. Taxonomic
studies have shown that these two taxa have different genotypes or molecular types
with biological and ecoepidemiological peculiarities. Matrix-assisted laser desorption
ionization–time of flight mass spectrometry (MALDI-TOF MS) has been proposed as
an alternative method for labor-intensive methods for
C. neoformans
and
C. gattii
genotype differentiation. However, Vitek MS, one of the commercial MALDI-TOF MS
instruments, has not been yet been evaluated for this purpose. Thus, we constructed
an in-house database with reference strains belonging to the different
C. neoformans
(VNI, VNII, VNIII, and VNIV) and
C. gattii
(VGI, VGII, VGIII, and VGIV) major molecular
types by using the software Saramis Premium (bioMérieux, Marcy-l’Etoile, France).
Then, this new database was evaluated for discrimination of the different genotypes.
Our in-house database provided correct identification for all
C. neoformans
and
C.
gattii
genotypes; however, due to the intergenotypic mass spectral similarities, a
careful postanalytic evaluation is necessary to provide correct genotype
identifica-tion.
KEYWORDS
Cryptococcus gattii
,
Cryptococcus neoformans
, MALDI-TOF, VITEK MS,
genotypic identification
T
he genus
Cryptococcus
comprises encapsulated yeasts that are divided in two main
pathogenic species,
Cryptococcus neoformans
and
Cryptococcus gattii
(1) However,
phylogenetic analyses have shown intraspecific genomic diversity among
C.
neofor-mans
and
C. gattii
strains, dividing them into eight major molecular types or genotypes,
as follows:
Cryptococcus neoformans
VNI, VNII, VNIII (AD hybrid), and VNIV and
C. gattii
VGI, VGII, VGIII, and VGIV (2, 3). Recent studies have shown that these eight major
molecular types have differences in their biology, ecoepidemiology, antifungal
suscep-tibility, and disease characteristics, making genotypic differentiation relevant (4).
Molecular methods, such as restriction fragment length polymorphism (RFLP),
am-plified fragment length polymorphism (AFLP), and multilocus or whole-genome
se-quence analysis, have been used to characterize these major eight molecular types;
however, these methods are labor costly, labor-intensive, and restricted to reference
laboratories (5). In 2011 and 2012, studies using the Bruker’s matrix-assisted laser
desorption ionization–time of flight mass spectrometry (MALDI-TOF MS) instrument
showed that this technique was able to discriminate the major molecular types of
C.
neoformans
and
C. gattii
(5–7). Thus, MALDI-TOF MS, an easy, fast, and inexpensive
method used in research and clinical laboratories, became an attractive alternative for
C. neoformans
and
C. gattii
genotype differentiation. The Vitek MS (bioMérieux,
Marcy-CitationSiqueira LPM, Gimenes VMF, de Freitas RS, Melhem MDSC, Bonfietti LX, da Silva AR, Jr, Souza Santos LB, Motta AL, Rossi F, Benard G, de Almeida JN, Jr. 2019. Evaluation of Vitek MS for differentiation ofCryptococcus neoformansandCryptococcus gattiigenotypes. J Clin Microbiol 57:e01282-18.https://doi.org/
10.1128/JCM.01282-18.
EditorDavid W. Warnock
Copyright© 2019 American Society for Microbiology.All Rights Reserved. Address correspondence to João N. de Almeida, Jr., [email protected]. Received9 August 2018
Returned for modification7 September 2018
Accepted31 October 2018
Accepted manuscript posted online14 November 2018
Published
crossm
2 January 2019
on May 17, 2020 by guest
http://jcm.asm.org/
l’Etoile, France), another MALDI-TOF MS instrument that has been used in clinical
laboratories, has shown performance for yeast identification similar to that of Bruker’s
instrument (8, 9). However, there are differences regarding mass spectrum acquisition,
identification algorithms, and reference spectrum creation between the instruments,
which makes relevant the evaluation of Vitek MS for
C. neoformans
and
C. gattii
major
molecular type differentiation (10, 11). Thus, aiming to fill this gap and to consolidate
the previous promising findings, we evaluated the Vitek MS (bioMérieux) and provided
technical details for future application of this technology for the differentiation of
C.
neoformans
and
C. gattii
molecular types.
MATERIALS AND METHODS
Fungal organisms. A total of 44 isolates belonging to the majorC. neoformans and C. gattii molecular types were used to construct in-house SuperSpectrum library, with 24 isolates from the National Institute of Quality Control in Health (INCQS; https://portal.fiocruz.br/en/unidade/national -institute-quality-control-health-incqs) and 20 clinical isolates maintained at the culture collection from the Tropical Medical Institute from São Paulo (University of São Paulo). To test the performance of the new database, 32 other isolates also representative of the majorC. neoformansandC. gattiimolecular types were included in the study, with 26 isolates from the mycological collection of the Federal University from Rio Grande do Norte, Brazil, and 6 additional isolates from the Tropical Medicine Institute (see Table S1 in the supplemental material). Clinical isolates were genotyped by URA5restriction fragment length polymorphism (RFLP) analysis, as previously described (12).
Sample preparation for MALDI-TOF MS analysis.Strains and isolates maintained as frozen stocks at⫺80°C in yeast extract-peptone-dextrose medium were subcultured on Sabouraud’s dextrose agar (SDA) plates and incubated for 48 h at 30°C before MALDI-TOF MS analysis. Then, a standard protein extraction protocol with ethanol and formic acid was carried out. In brief, one loop of yeast biomass was transferred into a 1.5-ml microcentrifuge tube containing 300l of purified water and mixed thoroughly. Subsequently, 900l of absolute ethanol was added to each tube and mixed for 1 min. The samples were centrifuged for 2 min at 13,000 rpm, and the supernatant was removed. The pellet was dried at room temperature, and 50l of formic acid (70%) was added. In addition, an equivalent volume of acetonitrile was added, and the mixture was centrifuged for 2 min at 13,000 rpm. Finally, 1l of the clear supernatant was spotted in quadruplicate onto a disposable MALDI target slide composed of a polypropylene carrier with a stainless steel layer (bioMérieux). After air-drying, each spot was overlaid with 1l of␣ -cyano-4-hydroxycinnamic acid (HCCA) matrix (bioMérieux).
MALDI-TOF MS analysis.Measurements were performed on a Vitek MS instrument (bioMérieux). For each acquisition group, a standard (Escherichia coliATCC 8739) was included to calibrate the instrument and validate the run. The spectra were generated using the Launchpad version 2.8 software (bioMérieux) and analyzed using the research use only (RUO) software Saramis Premium version 4.12 (bioMérieux). The in vitrodiagnostic (IVD) system was not evaluated due to the lack of C. neoformansand C. gattii genotype-specific mass spectrum profiles in the database.
Construction of an in-house SuperSpectrum library. For SuperSpectrum construction, mass spectra of 44 isolates belonging to the majorC. neoformansandC. gattiigenotypes (see Table S1) were imported into the Saramis Premium software package (bioMérieux). Then, 10 high-quality mass spectrum replicates (ⱖ120 masses,ⱖ70% similarity) of a given organism were selected to create a genotype-specific SuperSpectrum with the Saramis Premium SuperSpectrum tool (bioMérieux), according to the manufacturer’s instructions. The specificity of the potential biomarker masses was determined by comparison against the whole Saramis spectral archive (bioMérieux), and each SuperSpectrum had 40 biomarkers (masses) selected. Then, each biomarker was designated 35 points, given a total of 1,400 points for the final SuperSpectrum. Mass spectrum profiles from other Cryptococcus species (e.g., Cryptococcus laurentii[n⫽11],Cryptococcus curvatus[n⫽8],Cryptococcus uniguttulatus[n⫽12]) and from the genusTrichosporon(n⫽94) that are present in the original Saramis spectral archive version 4.12 (bioMérieux) were submitted to identification against the new in-house library to attest its specificity.
Genotype identification by MALDI-TOF MS.We initially evaluated the clustering of the different genotypes by mass spectrometry by creating a SuperSpectrum dendrogram and correlation matrix through the Saramis Premium software (bioMérieux). The dendrogram was generated based on whole spectra, with a single-link clustering algorithm and a binary mass list with an error of 800 ppm. Then, the 44 isolates that were used to construct the in-house SuperSpectrum library were subcultured once again and submitted to identification. Self-identifications (mass spectra identified by SuperSpectra from the same isolate) were not taken into account. Finally, the second set of 32 isolates, which were not used to construct the in-house SuperSpectrum library, were also submitted to identification (performance analysis step).
The Saramis Premium software (bioMérieux) compares the sample’s mass spectrum against Super-Spectra, and the sum of peak weights is computed after matching mass signals of each SuperSpectrum and is transformed into a confidence value and points as follows (13): dark green with 99.9% confidence values or 1,000 to 1,400 points, light green with 90% to 99% confidence values or 900 to 999 points, yellow with 80% to 89.9% confidence values or 800 to 899 points, and white with 75% to 79.9% confidence values or 750 to 799 points.
Siqueira et al. Journal of Clinical Microbiology
on May 17, 2020 by guest
http://jcm.asm.org/
RESULTS
The dendrogram split the SuperSpectra of
C. neoformans
and
C. gattii
into two
species-specific branches (Fig. 1). These main branches showed a clear distinction
between the two species, with SuperSpectra showing more than 20 species-specific
biomarkers (Fig. 1). Moreover, the cryptococcal isolates were separated into clusters
corresponding to the main genotypes of the two
Cryptococcus
species (Fig. 1). No single
mass spectrum or SuperSpectrum from
Cryptococcus
species other than
C. neoformans
and
C. gattii
or from
Trichosporon
species showed cross-identification with the new
in-house library.
The correlation matrix of the
C. neoformans
SuperSpectra (each one with 40
bio-FIG 1Saramis Premium dendrogram clustering of SuperSpectra from eachCryptococcus neoformansand Crypto-coccus gattiiorganism, with distances displayed in number of identical masses. Filter was tolerance of 0.08%, absolute intensity ofⱖ0, relative intensity ofⱖ0, and mass range 3,000 to 20,000 Da.
on May 17, 2020 by guest
http://jcm.asm.org/
[image:3.585.41.395.68.557.2]markers) showed higher (twice) intragenotype than intergenotype similarity, as there
was a mean of 24 intragenotype common biomarkers compared with only 12
inter-genotype common biomarkers (Fig. S1A). The correlation matrix of the
C. gattii
Super-Spectrum (each one with 40 biomarkers) also showed higher intragenotype than
intergenotype similarity; there were means of 27 and 13 intragenotype and
intergeno-type common biomarkers, respectively (Fig. S1B).
All isolates that were used to construct the in-house SuperSpectrum library had
correct species assignment as
C. neoformans
and
C. gattii
by the new in-house
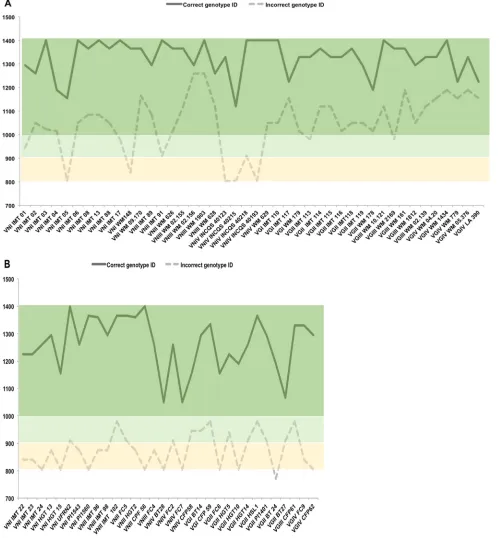
Super-Spectrum library. Regarding the results for genotype identification, the provided
con-fidence intervals were not discriminatory, and ambiguous results with
⬎
90%
confi-dence level values occurred in 20 of the 24 (83%) and in 20 of the 20 (100%)
C.
neoformans
and
C. gattii
organisms, respectively (Fig. 2A). However, the identification
points were able to correctly discriminate all isolate genotypes, with the higher point
values corresponding to the accurate genotype.
Among the 32 additional isolates used in the performance analysis step, all had
correct species assignment as
C. neoformans
and
C. gattii
by the new in-house
Super-Spectrum library. For genotype identification, the provided confidence intervals were
also not discriminatory, and ambiguous results with
⬎
90% confidence level values
occurred in 5 of the 19 (26%) and in 7 of the 13 (54%)
C. neoformans
and
C. gattii
organisms, respectively (Table 1). Yet, genotype identification of all isolates was
effi-ciently achieved by the reported point values, with higher point values corresponding
to the accurate genotype (Table 1).
Figure 2 illustrates that interpretation of the results provided by Saramis Premium
software (bioMérieux) using point values instead of confidence levels is able to correctly
discriminate all
C. neoformans
and
C. gattii
genotypes.
DISCUSSION
In the last 20 years, taxonomic studies with the help of molecular tools have
recognized several new (cryptic) species among the pathogenic fungi (14, 15). The new
taxonomic evidence added to biological traits, and to epidemiological and clinical data,
has resulted in powerful and holistic information that is helping better understand
transmission routes and mechanisms of acquisition, implement disease control
mea-sures, and finally, achieve a better organism-targeted therapy (16). For example, it was
shown that VGII and VGIV isolates show higher MICs for azole derivatives than do
isolates of
C. neoformans
and the other
C. gattii
(17) genotypes, and VGII isolates have
been linked to outbreaks of invasive disease in immunocompetent hosts in North
America (18) and Brazil (19). Indeed, the assembly of these relevant data with
genea-logical analyses has led experts to propose that the
C. neoformans
and
C. gattii
major
molecular types are indeed at least seven different species, including
C. neoformans
(genotypes VNI and VNII),
C. deneoformans
(VNIV),
C. gattii
(VGI),
C. deuterogattii
(VGII),
C. bacillisporus
(VGIII),
C. tetragattii
(VGIV), and
C. decagattii
(2, 20).
Since MALDI-TOF MS is becoming the method of choice for microorganism
identi-fication in clinical laboratories all over the world, systematic identiidenti-fication of
C.
neofor-mans
and
C. gattii
major molecular types by using this technology may be achieved in
the future. However, validated reference spectrum libraries including several
represen-tatives of each genotypes/species are necessary to produce accurate results and to
minimize the number of misidentifications (21). Moreover, the identification algorithms
of the different MALDI-TOF systems may not have sufficient discriminatory power to
separate some closely related taxa. Thus, additional postanalytical steps may be
re-quired for the correct organisms’ identification (8, 22). Dendrograms generated with the
RUO tools (e.g., BioTyper and Saramis) provide preliminary yet useful information
regarding the relatedness of the different species by its main mass spectrum profiles.
As previously reported by studies that used the Bruker’s MALDI-TOF MS apparatus
(5–7), the dendrogram created with the Saramis Premium software segregated the
major molecular types of
C. neoformans
and
C. gattii
into different and
genotype-specific clades. Despite clear genotype distinction provided by the dendrogram, initial
Siqueira et al. Journal of Clinical Microbiology
on May 17, 2020 by guest
http://jcm.asm.org/
evaluation showed that 91% of the organisms had ambiguous identifications with
⬎
90% confidence values. As shown in the correlation matrix, some SuperSpectra
belonging to different genotypes had more than 20 common biomarkers. These
intergenotypic similar mass spectrum profiles when analyzed by the Saramis’s
match-FIG 2Identification points provided for each isolate by the Saramis Premium software (bioMérieux). The values for the correct genotype identifications are depicted in a continuous line, while those for incorrect genotype identifications are provided in a dashed line. Color boxes correspond to the confidence level identifications, as follows: dark green with 99.9% confidence values, light green with 90% to 99% confidence values, and yellow with 80% to 89.9%. As noticed in the light and dark green boxes, most of the isolates had cross-identification with other genotypes with 90% to 99.9% confidence levels. However, higher identification points provided correct genotype identifications. (A) Identifications for the isolates that were used for the construction of the in-house library. (B) Identifications for the isolates that were used during the performance step.
on May 17, 2020 by guest
http://jcm.asm.org/
[image:5.585.39.538.75.613.2]ing signals method may result in ambiguous identifications if only the confidence level
percentage is taken into account. However, by using an alternative algorithm that took
into account only the higher identification points, all organisms had unambiguous
genotype assignment by the Saramis software.
Alternatively, the Vitek MS IVD system (bioMérieux) uses an identification algorithm
method based on a process called “mass binning” that has shown better performance
for microorganisms identification than Saramis’s method of matching mass signals (13,
23, 24). Thus, although not evaluated here, an upgrade of the IVD database with the
major
C. neoformans
and
C. gattii
genotypes by the manufacturer could enhance the
power of Vitek MS to well discriminate these microorganisms.
In conclusion, we showed that Vitek MS (bioMérieux) instrument has the potential
to well discriminate the major molecular types of
C. neoformans
and
C. gattii
. The
Saramis software enables the construction of genotype-specific SuperSpectra, but a
robust in-house library and an alternative identification algorithm are necessary to
achieve accurate performance.
SUPPLEMENTAL MATERIAL
Supplemental material for this article may be found at
https://doi.org/10.1128/JCM
.01282-18
.
SUPPLEMENTAL FILE 1
, PDF file, 8.5 MB.
ACKNOWLEDGMENTS
We thank Márcia dos Santos Lazéra and Ivano de Filippis (National Institute of
Quality Control in Health, Instituto Oswaldo Cruz-FIOCRUZ) and Raquel Cordeiro
The-odoro (Federal University of Rio Grande do Norte, Brazil) for kindly providing isolates.
This research received no specific grant from any funding agency in the public,
commercial, or not-for-profit sector. The work of G.B. is supported by a grant from the
National Council of Technological and Scientific Development (CNPQ 421374/2016-0).
We declare no conflicts of interest.
REFERENCES
1. Chang YC, Kwon-Chung KJ. 1994. Complementation of a capsule-deficient mutation of Cryptococcus neoformans restores its virulence. Mol Cell Biol 14:4912– 4919.https://doi.org/10.1128/MCB.14.7.4912. 2. Hagen F, Khayhan K, Theelen B, Kolecka A, Polacheck I, Sionov E, Falk
R, Parnmen S, Lumbsch HT, Boekhout T. 2015. Recognition of seven species in the Cryptococcus gattii/Cryptococcus neoformans species complex. Fungal Genet Biol 78:16 – 48.https://doi.org/10.1016/j.fgb .2015.02.009.
3. Severo CB, Xavier MO, Gazzoni AF, Severo LC. 2009. Cryptococcosis in
children. Paediatr Respir Rev 10:166 –171.https://doi.org/10.1016/j.prrv .2009.06.009.
4. Del Poeta M, Casadevall A. 2012. Ten challenges on cryptococcus and cryptococcosis. Mycopathologia 173:303–310.https://doi.org/10.1007/ s11046-011-9473-z.
5. Hagen F, Huijsmans A, Klaassen CH, Boekhout T, Meis JF. 2011. MALDI-TOF based identification of the pathogenic yeast species C. neoformans and C. gattii. Mycoses 54:78.
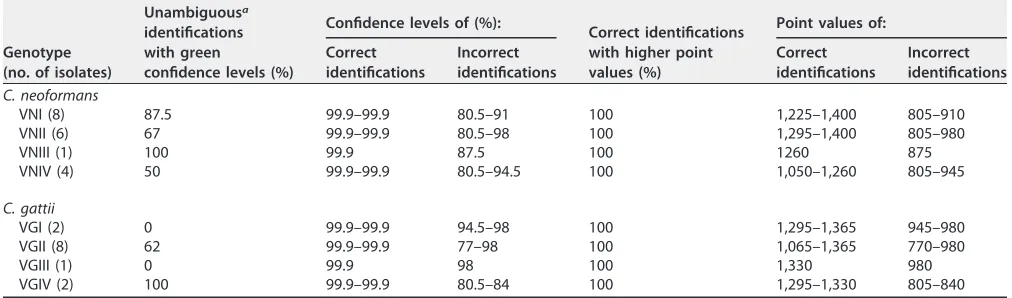
[image:6.585.40.546.94.243.2]6. Firacative C, Trilles L, Meyer W. 2012. MALDI-TOF MS enables the rapid TABLE 1Cryptococcus neoformansandCryptococcus gattiigenotypes identification provided by the Saramis Premium software equipped with the in-house SuperSpectrum library during the performance step
Genotype (no. of isolates)
Unambiguousa
identifications with green
confidence levels (%)
Confidence levels of (%):
Correct identifications with higher point values (%)
Point values of: Correct
identifications
Incorrect identifications
Correct identifications
Incorrect identifications
C. neoformans
VNI (8) 87.5 99.9–99.9 80.5–91 100 1,225–1,400 805–910
VNII (6) 67 99.9–99.9 80.5–98 100 1,295–1,400 805–980
VNIII (1) 100 99.9 87.5 100 1260 875
VNIV (4) 50 99.9–99.9 80.5–94.5 100 1,050–1,260 805–945
C. gattii
VGI (2) 0 99.9–99.9 94.5–98 100 1,295–1,365 945–980
VGII (8) 62 99.9–99.9 77–98 100 1,065–1,365 770–980
VGIII (1) 0 99.9 98 100 1,330 980
VGIV (2) 100 99.9–99.9 80.5–84 100 1,295–1,330 805–840
aOnly the correct genotype was named with confidence levels above 90%.
Siqueira et al. Journal of Clinical Microbiology
on May 17, 2020 by guest
http://jcm.asm.org/
identification of the major molecular types within the Cryptococcus neoformans/C. gattii species complex. PLoS One 7:e37566.https://doi .org/10.1371/journal.pone.0037566.
7. Posteraro B, Vella A, Cogliati M, De Carolis E, Florio AR, Posteraro P, Sanguinetti M, Tortorano AM. 2012. Matrix-assisted laser desorption ionization–time of flight mass spectrometry-based method for discrim-ination between molecular types of Cryptococcus neoformans and Cryp-tococcus gattii. J Clin Microbiol 50:2472–2476.https://doi.org/10.1128/ JCM.00737-12.
8. de Almeida JN, Jr, Favero Gimenes VM, Francisco EC, Machado Siqueira LP, Gonçalves de Almeida RK, Guitard J, Hennequin C, Colombo AL, Benard G, Rossi F. 2017. Evaluating and improving Vitek MS for identi-fication of clinically relevant species of Trichosporon and the closely related genera Cutaneotrichosporon and Apiotrichum. J Clin Microbiol 55:2439 –2444.https://doi.org/10.1128/JCM.00461-17.
9. Zhou L, Chen Y, Xu Y. 2017. Performance of VITEK mass spectrometry V3.0 for rapid identification of clinical Aspergillus fumigatus in different culture conditions based on ribosomal proteins. Infect Drug Resist 10: 499 –506.https://doi.org/10.2147/IDR.S148121.
10. Pappalardo MC, Melhem MS. 2003. Cryptococcosis: a review of the Brazilian experience for the disease. Rev Inst Med Trop Sao Paulo 45:299 –305.https://doi.org/10.1590/S0036-46652003000600001. 11. Lacaz CS, Porto E, Martins JEC, Heins-Vaccari EM, Takahashi De Melo N.
2002. Tratado de micologia médica. Sarvier, São Paulo, Brazil. 12. Meyer W, Castañeda A, Jackson S, Huynh M, Castañeda E, IberoAmerican
Cryptococcal Study Group. 2003. Molecular typing of IberoAmerican Cryptococcus neoformans isolates. Emerg Infect Dis 9:189 –195.https:// doi.org/10.3201/eid0902.020246.
13. Welker M. 2011. Proteomics for routine identification of microorganisms. Proteomics 11:3143–3153.https://doi.org/10.1002/pmic.201100049. 14. Hibbett DS, Binder M, Bischoff JF, Blackwell M, Cannon PF, Eriksson OE,
Huhndorf S, James T, Kirk PM, Lücking R, Thorsten Lumbsch H, Lutzoni F, Matheny PB, McLaughlin DJ, Powell MJ, Redhead S, Schoch CL, Spatafora JW, Stalpers JA, Vilgalys R, Aime MC, Aptroot A, Bauer R, Begerow D, Benny GL, Castlebury LA, Crous PW, Dai Y-C, Gams W, Geiser DM, Griffith GW, Gueidan C, Hawksworth DL, Hestmark G, Hosaka K, Humber RA, Hyde KD, Ironside JE, Kõljalg U, Kurtzman CP, Larsson K-H, Lichtwardt R, Longcore J, Miadlikowska J, Miller A, Moncalvo J-M, Mozley-Standridge S, Oberwinkler F, Parmasto E, Reeb V, et al. 2007. A higher-level phylogenetic classification of the Fungi. Mycol Res 111: 509 –547.https://doi.org/10.1016/j.mycres.2007.03.004.
15. Hittinger CT, Rokas A, Bai F-Y, Boekhout T, Gonçalves P, Jeffries TW, Kominek J, Lachance M-A, Libkind D, Rosa CA, Sampaio JP, Kurtzman CP. 2015. Genomics and the making of yeast biodiversity. Curr Opin Genet Dev 35:100 –109.https://doi.org/10.1016/j.gde.2015.10.008.
16. Chen SC-A, Meyer W, Sorrell TC. 2014. Cryptococcus gattii infections. Clin Microbiol Rev 27:980 –1024.https://doi.org/10.1128/CMR.00126-13. 17. Espinel-Ingroff A, Aller AI, Canton E, Castañón-Olivares LR, Chowdhary A,
Cordoba S, Cuenca-Estrella M, Fothergill A, Fuller J, Govender N, Hagen F, Illnait-Zaragozi MT, Johnson E, Kidd S, Lass-Flörl C, Lockhart SR, Martins MA, Meis JF, Melhem MSC, Ostrosky-Zeichner L, Pelaez T, Pfaller
MA, Schell WA, St-Germain G, Trilles L, Turnidge J. 2012. Cryptococcus neoformans-Cryptococcus gattii species complex: an international study of wild-type susceptibility endpoint distributions and epidemiological cutoff values for fluconazole, itraconazole, posaconazole, and voricona-zole. Antimicrob Agents Chemother 56:5898 –5906.https://doi.org/10 .1128/AAC.01115-12.
18. Kidd SE, Hagen F, Tscharke RL, Huynh M, Bartlett KH, Fyfe M, Macdougall L, Boekhout T, Kwon-Chung KJ, Meyer W. 2004. A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proc Natl Acad Sci U S A 101: 17258 –17263.https://doi.org/10.1073/pnas.0402981101.
19. Trilles L, Lazéra MS, Wanke B, Oliveira RV, Barbosa GG, Nishikawa MM, Morales BP, Meyer W. 2008. Regional pattern of the molecular types of Cryptococcus neoformans and Cryptococcus gattii in Brazil. Mem Inst Oswaldo Cruz 103:455– 462. https://doi.org/10.1590/S0074 -02762008000500008.
20. Hagen F, Lumbsch HT, Arsic Arsenijevic V, Badali H, Bertout S, Billmyre RB, Bragulat MR, Cabañes FJ, Carbia M, Chakrabarti A, Chaturvedi S, Chaturvedi V, Chen M, Chowdhary A, Colom M-F, Cornely OA, Crous PW, Cuétara MS, Diaz MR, Espinel-Ingroff A, Fakhim H, Falk R, Fang W, Herkert PF, Ferrer Rodríguez C, Fraser JA, Gené J, Guarro J, Idnurm A, Illnait-Zaragozi M-T, Khan Z, Khayhan K, Kolecka A, Kurtzman CP, Lagrou K, Liao W, Linares C, Meis JF, Nielsen K, Nyazika TK, Pan W, Pekmezovic M, Polacheck I, Posteraro B, de Queiroz Telles F, Romeo O, Sánchez M, Sampaio A, Sanguinetti M, Sriburee P, et al. 2017. Importance of resolv-ing fungal nomenclature: the case of multiple pathogenic species in the Cryptococcus genus. mSphere 2:e00238-17.
21. Normand A-C, Cassagne C, Ranque S, L’ollivier C, Fourquet P, Roesems S, Hendrickx M, Piarroux R. 2013. Assessment of various parameters to improve MALDI-TOF MS reference spectra libraries constructed for the routine identification of filamentous fungi. BMC Microbiol 13:76.https:// doi.org/10.1186/1471-2180-13-76.
22. Grenfell RC, da Silva Junior AR, Del Negro GMB, Munhoz RB, Gimenes VMF, Assis DM, Rockstroh AC, Motta AL, Rossi F, Juliano L, Benard G, de Almeida Júnior JN. 2016. Identification of Candida haemulonii complex species: use of ClinProTools to overcome limitations of the Bruker Biotyper, VITEK MS IVD, and VITEK MS RUO databases. Front Microbiol 7:940.
23. Leyer C, Gregorowicz G, Mougari F, Raskine L, Cambau E, de Briel D. 2017. Comparison of Saramis 4.12 and IVD 3.0 Vitek MS matrix-assisted laser desorption ionization–time of flight mass spectrometry for identi-fication of mycobacteria from solid and liquid culture media. J Clin Microbiol 55:2045–2054.https://doi.org/10.1128/JCM.00006-17. 24. Rychert J, Burnham C-AD, Bythrow M, Garner OB, Ginocchio CC,
Jenne-mann R, Lewinski MA, Manji R, Mochon AB, Procop GW, Richter SS, Sercia L, Westblade LF, Ferraro MJ, Branda JA. 2013. Multicenter evaluation of the Vitek MS matrix-assisted laser desorption ionization-time of flight mass spectrometry system for identification of Gram-positive aerobic bacteria. J Clin Microbiol 51:2225–2231. https://doi.org/10.1128/JCM .00682-13.