Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Genetic and Phenotypic Features of
Streptococcus pyogenes
Strains
Isolated in Brazil That Harbor New
emm
Sequences
LU´ CIA M. TEIXEIRA,1* ROSANA R. BARROS,1ANGELA CHRISTINA D. CASTRO,1JOSE´ MAURO PERALTA,1 MARIA DA GLO´ RIA S. CARVALHO,1DEBORAH F. TALKINGTON,2ADRIANA M. VIVONI,1
RICHARD R. FACKLAM,2ANDBERNARD BEALL2
Instituto de Microbiologia, Universidade Federal do Rio de Janeiro, Rio de Janeiro 21941, Brazil,1and
Division of Bacterial and Mycotic Diseases, Respiratory Diseases Branch, Centers for
Disease Control and Prevention, Atlanta, Georgia 303332
Received 13 April 2001/Returned for modification 2 June 2001/Accepted 5 July 2001
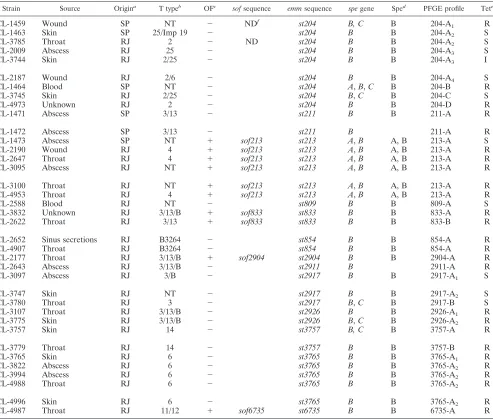
In the present study, 37 group A Streptococcus(GAS) strains belonging to 13 new emmsequence types identified among GAS strains randomly isolated in Brazil were characterized by using phenotypic and geno-typic methods. The new types were designatedst204,st211, st213, st809, st833, st854, st2904, st2911,st2917,
st2926, st3757,st3765, andst6735. All isolates were susceptible to the antimicrobial agents tested, except to tetracycline. They all carried thespeBgene, and 94.6% produced detectable SpeB. Most strains belonging to a givenemmtype had similar or highly related pulsed-field gel electrophoresis profiles that were distinct from profiles of strains of another type. The other characteristics were variable from isolate to isolate, although some associations were consistently found within someemmtypes. Unlike the other isolates, all typest213
isolates werespeApositive and produced SpeA. Strains belonging tost3765were T6 and opacity factor (OF) negative. Individual isolates within OF-positiveemmtypes were associated with uniquesofgene sequence types, while OF-negative isolates weresofnegative by PCR. This report provides information on newemmsequence types first detected in GAS isolates from a geographic area not extensively surveyed. Such data can contribute to a better understanding of the local and global dynamics of GAS populations and of the epidemiological aspects of GAS infections occurring in tropical regions.
Streptococcus pyogenes, frequently referred to as group A
Streptococcus(GAS), is a significant cause of human morbidity
worldwide, and it is associated with a variety of mild and severe diseases that may occur either in areas where the diseases are endemic or as epidemics (10). Most of the knowledge that has been accumulated concerning GAS epidemiology is based upon serologic M and T typing. However, many GAS isolates are nontypeable due to the lack of appropriate type-specific antisera or possibly due to loss of antigen expression under cultivation. In recent years, DNA sequencing-based methods for characterizing GAS strains have been used, including se-quence analysis of emm gene-specific PCR products (emm
typing) (1, 2). This methodology has allowed the recognition of several previously unknown GAS types in different geographic areas, demonstrating the usefulness ofemmtyping for detect-ing genetic diversity among GAS isolates and for tracdetect-ing GAS infections (1, 2, 5). A complementary molecular methodology (softyping) is also based upon sequence analysis of a hyper-variable virulence gene (3).
In the present study, we describe the characteristics of GAS isolates belonging to newemmtypes identified among strains recovered from patients living in Brazil. These strains were further characterized through phenotypic tests (T-protein typ-ing, detection of opacity factor [OF], antimicrobial suscepti-bility, and production of streptococcal pyrogenic exotoxin A [SpeA] and SpeB) and additional genotypic tests (the presence
ofspeA,speB,speC, andsofgene PCR products,sofsequence types, and analysis of genomic DNA restriction profiles).
MATERIALS AND METHODS
Bacterial strains.A total of 37 GAS isolates belonging to newemmtypes were
included in the present study. They were recognized among random GAS iso-lates recovered from sterile- and nonsterile-site clinical specimens obtained be-tween 1995 and 1999 from patients living in the southeast region of Brazil. The isolates were identified on the basis of colony morphology and-hemolysis on blood agar plates, gram stain characteristics, susceptibility to bacitracin, produc-tion of leucine aminopeptidase and pyrrolidonyl arylamidase, and reactivity with group A-specific antiserum.
Phenotypic characterization of the isolates.Serologic T types and opacity
factor (OF) reactions were determined as previously described (7, 8). Suscepti-bility to antimicrobial agents was tested by a broth microdilution assay using the CDC Strep panel (PML Microbiologicals, Wilsonville, Oreg.). The following 14 antimicrobial agents were tested: penicillin, amoxicillin, cefotaxime, cefuroxime, erythromycin, trimethoprim-sulfamethoxazole, clindamycin, chloramphenicol, tetracycline, vancomycin, trovafloxacin, rifampin, meropenem, and levofloxacin. The MIC results were interpreted according to NCCLS guidelines (9).
Detection of exotoxin production by immunoblotting assay. Strains were
tested for their ability to produce SpeA and SpeB in vitro by using procedures for exotoxin preparation and for exotoxin detection by a immunoblotting assay derived from those outlined by Talkington et al. (11).
Detection of streptococcal pyrogenic exotoxin genes by PCR.PCRs were
per-formed to detect the presence of thespeA,speB, andspeCgenes. DNA extraction and PCR conditions were based on previously described methods (4, 11).
Analysis of chromosomal DNA restriction profiles by PFGE.Genomic DNA
was prepared with agarose plugs, based on previously described recommenda-tions (12). For lysis, plugs were treated with lysis solution containing 1 mg of lysozyme and 5 U of mutanolysin per ml. After digestion withSmaI, the frag-ments were resolved by pulsed-field gel electrophoresis (PFGE) in 1.3% agarose gels in 0.5⫻Tris-borate-EDTA buffer in a CHEF-DR III system (Bio-Rad Laboratories, Hercules, Calif.). The following parameters were used: running time, 25 h; temperature, 13°C; voltage gradient, 6 V/cm; initial pulse time, 5.3 s; final pulse time, 34.9 s. The DNA fragments in the gels were stained with ethidium bromide and photographed under UV light. Analysis of chromosomal
* Corresponding author. Mailing address: Instituto de Microbiolo-gia, Universidade Federal do Rio de Janeiro, CCS, Bloco I, Cidade Universita´ria, Rio de Janeiro, RJ 21941, Brazil. Phone: 55 21 260 4193. Fax: 55 21 560 8344. E-mail: [email protected].
3290
on May 15, 2020 by guest
http://jcm.asm.org/
DNA fragmentation profiles was performed by visual inspection of photographic registers, considering the criteria suggested by Tenover et al. (13). PFGE profiles were also analyzed and compared by using the Molecular Analyst Fingerprint Plus software package, version 1.12 (Bio-Rad). The percentages of similarities of the PFGE banding profiles were estimated with the Dice coefficient and clus-tered by the unweighted pair group method with arithmetic averages.
Detection ofsofgenes andsofsequence analysis.Amplification of 5⬘sofgene
fragments andsofsequence analysis were performed as previously described (3). GenBank accession numbers forsofgene sequences are as follows:sof213, AF 139743;sof833(actually an allele ofsof90; see reference 3), AF139741;sof6735
(also described in reference 3, with a 5⬘sequence very similar in overlap to
sof90).
Sequencing ofemmgene-specific PCR products (emmtyping).Harvesting of
the bacterial cells and the procedures for PCR and sequence analysis ofemm -specific PCR products were essentially those already published (1, 2). DNA sequences were subjected to homology searches against all known emm se-quences present in the GenBank and in the Centers for Disease Control and
Prevention (CDC) database (http://www.cdc.gov/ncidod/biotech/infotech_hp .html) with Genetics Computer Group software, version 9, with the NCBI Blast programs. GenBank accession numbers are as follows:st204, AF056301;st211, AF 96179; st213, AF049855; st809, AF296180; st833, AF052425; st854, AF281048;st2904, AF192768;st2917, AF082864;st3757, AF074875; andst3765, AF074877.
Phylogram construction of M protein sequences.Sequences with 103 residues
(including 23 signal sequence residues and 80 amino acids of the mature N termini) were analyzed by sequential use of the Wisconsin Package (version 10.1) PileUp (gap creation and extension penalties of 8 and 2, respectively), Distances (uncorrected distance), and GrowTree (neighbor-joining) programs.
RESULTS AND DISCUSSION
The 37 GAS isolates selected for the present study were found to correspond to 13 previously undocumentedemm
se-CL-2647 Throat RJ 4 ⫹ sof213 st213 A,B A, B 213-A R
CL-3095 Abscess RJ NT ⫹ sof213 st213 A,B A, B 213-A R
CL-3100 Throat RJ NT ⫹ sof213 st213 A,B A, B 213-A R
CL-4953 Throat RJ 4 ⫹ sof213 st213 A,B A, B 213-A R
CL-2588 Blood RJ NT ⫺ st809 B B 809-A S
CL-3832 Unknown RJ 3/13/B ⫹ sof833 st833 B B 833-A R
CL-2622 Throat RJ 3/13 ⫹ sof833 st833 B B 833-B R
CL-2652 Sinus secretions RJ B3264 ⫺ st854 B B 854-A R
CL-4907 Throat RJ B3264 ⫺ st854 B B 854-A R
CL-2177 Throat RJ 3/13/B ⫹ sof2904 st2904 B B 2904-A R
CL-2643 Abscess RJ 3/13/B ⫺ st2911 B 2911-A R
CL-3097 Abscess RJ 3/B ⫺ st2917 B B 2917-A1 S
CL-3747 Skin RJ NT ⫺ st2917 B B 2917-A2 S
CL-3780 Throat RJ 3 ⫺ st2917 B,C B 2917-B S
CL-3107 Throat RJ 3/13/B ⫺ st2926 B B 2926-A1 R
CL-3775 Skin RJ 3/13/B ⫺ st2926 B,C B 2926-A2 R
CL-3757 Skin RJ 14 ⫺ st3757 B, C B 3757-A R
CL-3779 Throat RJ 14 ⫺ st3757 B B 3757-B R
CL-3765 Skin RJ 6 ⫺ st3765 B B 3765-A1 R
CL-3822 Abscess RJ 6 ⫺ st3765 B B 3765-A2 R
CL-3994 Abscess RJ 6 ⫺ st3765 B B 3765-A2 R
CL-4988 Throat RJ 6 ⫺ st3765 B B 3765-A2 R
CL-4996 Skin RJ 6 ⫺ st3765 B B 3765-A2 R
CL-4987 Throat RJ 11/12 ⫹ sof6735 st6735 B B 6735-A R
aLocality of isolation: RJ, Rio de Janeiro; SP, Sa˜o Paulo. bNT, T nontypeable.
c⫹, OF positive;⫺, OF negative.
dProduction of SpeA and or SpeB as detected by immunoblotting assay.
eResults of antimicrobial susceptibility tests for tetracycline: R, resistant; S, susceptible; I, intermediate. fND, not determined.
on May 15, 2020 by guest
[image:2.612.57.550.88.507.2]quence types. They comprised 12.3% of a total of 302 random GAS isolates recovered in Brazil, which are the subject of a study in progress. Phenotypic and genotypic characteristics of the 37 GAS isolates representing newemmtypes are shown in Table 1, in addition to their sources and city of isolation.
All strains were susceptible to most of the antimicrobial agents tested. The exception was tetracycline, to which 26 strains were resistant and 1 was intermediate. ThespeBgene was detected in all isolates, while speAand speCgenes were found in seven and six isolates, respectively. Most isolates (94.6%) produced SpeB. PFGE profiles of isolates belonging to a givenemmtype were usually similar or highly related. In some instances, PFGE profiles within a givenemmtype were not related, but they were always distinct from profiles of strains of anotheremmtype (Fig. 1 and 2). This data indicated that the majority of isolates within each individualemmtype shared a high degree of genetic relatedness. This high degree of relatedness within certainemmtypes might hold true only within this particular geographical area, sinceemmsequence type 854 (st854) isolates recovered in Egypt as this study was being completed had PFGE profiles clearly distinct from those of the Brazilianst854isolates (Fig. 1C). It should also be noted that after its initial discovery in 1997 in Brazilian isolates (this study), the 549-base emm st854 sequence was found to be identical over its overlap with material recently assigned Gen-Bank accession number AF018179 (225 bases), derived from an impetigo isolate in Northern Australia (6).
Several characteristics were variable from strain to strain, although some associations were found withinemmtypes. Six of nine isolates representing the predominant new type st204
had related PFGE profiles, despite differences in susceptibility to tetracycline and T-typing results. T types and OF pheno-types can vary extensively even between different passages of the same strain, possibly due to the effects of laboratory growth conditions on gene expression (unpublished observations). All
sixst213isolates contained bothspeAand speBgenes and, in
contrast to all of the other isolates, produced both SpeA and SpeB. All sixst213isolates were OF positive and contained the unique sof213 sequence. Threest213 isolates were T type 4, while the other three were T nontypeable. The five st3765
isolates were T type 6 and OF negative. All threest2917 iso-lates were susceptible to tetracycline and were OF negative.
Two st2917 isolates typed with components of the T 3/13/B
complex while one was T nontypeable.
Characteristics varied among isolates with the other new
emm types represented by one or two isolates (st211, st809,
st833,st854,st2904,st2911,st2926,st3757, andst6735). Most of
these isolates typed with components of the T 3/13/B complex, except forst3757isolates (T type 14) and st6735isolates (T type 11/12).
Representative isolates from 4 of the 13 new emm types shown in Table 1 were found to be sof PCR positive. The partialsof213 sequence from the st213 isolate CL-1473 was unique from all othersofsequences analyzed to date (at least 80 at present time [3]), with only 79% sequence identity to its closest match. Similarly, the best match tosof2904 was only 80% identical over 547 bases. It was interesting to find that the 5⬘ sof sequences associated with bothst833and st6735types were nearly identical tosof90from anemmtype M serotype 90 isolate (3). While thest833sequence is very similar toemm90, it differs fromemm90by the deletion of the codons encoding mature M residues 2 to 5 and 17 to 21 and does not confer the M90 serotype. Nonetheless, ast833 sof833isolate was found to be anti-OF type 90 (3). The similarity between this pair of chromosomally unlinked genes with these genes from the
emm90reference strain, together with related T-agglutination
[image:3.612.70.542.74.242.2]patterns (either T 3/13 or T 3/13/B3264) suggests that sequence typesst833,sof833, andemm90 sof90strains are highly related genomically (see http://www.cdc.gov/ncidod/biotech/infotech _hp.html for information concerningemm90and other known FIG. 1. Representative PFGE profiles ofSmaI-digested genomic DNA ofStreptococcus pyogenesbelonging to newemmsequence types isolated in Brazil. Lanes 1 (A through C), 11 (B), and 20 (A), molecular mass markers expressed in kilobases (lambda DNA concatemers ranging from 48.5 to 1,018.5 kb). (A) Lanes: 2 to 10,st204strains (lane 2, CL-1459; lane 3, CL-1463; lane 4, CL-3785; lane 5, CL-2009; lane 6, CL-3744; lane 7, CL-2187; lane 8, CL-4973; lane 9, CL-1464; lane 10, CL-3745); 12 and 13,st211strains (lane 12, CL-1471; lane 13, CL-1472); 14 to 19,st213strains (lane 14, CL-1473; lane 15, CL-2190; lane 16, CL-2647; lane 17, CL-3095; lane 18, CL-3100; lane 19, CL-4953). (B) Lanes: 2,st809strain CL-2588; 3 and 4,st833strains (lane 3, CL-3832; lane14, CL-2622); 5,st2904strain CL-2177; 6,st2911strain CL-2643; 7 and 8,st2917strains (lane 7, CL-3097; lane 8, CL-3780); 9 and 10,st2926strains (lane 9, CL-3107; lane 10, CL-3775); 11 and 12,st3757strains (lane 11, CL-3757; lane 12, CL-3779); 13 to 15,st3765strains (lane 13, CL-3765; lane 14, CL-3822; lane 15, CL-3994). (C) Lanes: 2 and 3,st854strains from Brazil (lane 2, CL-2652; lane 3, CL-4907); 4 to 6,st854strains from Egypt; 7,st6735strain CL-4987; 8,st3765strain CL-4996.
on May 15, 2020 by guest
http://jcm.asm.org/
emmtypes). In contrast, whilesof6735andsof90(orsof833) 5⬘ sequences were highly similar (3), thest6735sequence and T types were found to be quite divergent from these markers in
theemm90reference strain. The PFGE results clearly indicate
that CL-4987 (st6735) and CL-2622 (st833) had distinct chro-mosomal restriction profiles (Table 1; Fig. 1 and 2).
Figure 3 shows a dendrogram based upon sequence com-parisons of 114 residue N-terminal portions of the various deduced M proteins representing the new sequence types first encountered in Brazil. These 114 residues include 19 signal sequence amino acids and the 95 mature N terminal residues. For reference, the corresponding sequence from the closest “classical” M protein sequence match was included in the analysis. It is striking that all of the 17 sequences, representing OF- andsof-negative strains, were neatly segregated from the 4 sequences derived from OF- andsof-positive strains, with the single exception ofemmtypest211, which was obtained from an OF- andsofPCR-negative isolate. This relationship, where
the majority of OF-positiveemmtypes are segregated from the majority of OF-negativeemmtypes, holds true with a similar analysis representing the majority of known deduced M se-quences (14; www.cdc.gov/ncidod/biotech/strep/strepindex.html). These results support the notion that OF-negative and -posi-tive strains represent two distinct GAS lineages between which little horizontal exchange occurs. As also shown in Fig. 3, the branch point foremmtypest211is much deeper than the other branch points in the OF-positive section of the dendrogram, which indicates that this sequence is the most divergent se-quence included in this analysis.
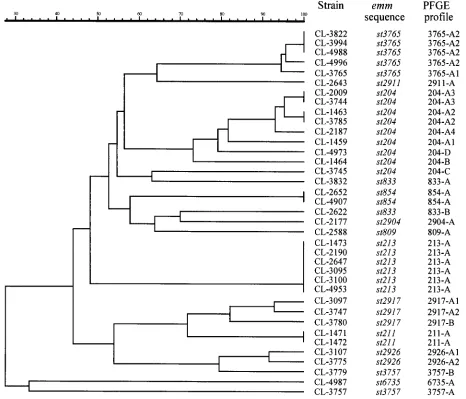
[image:4.612.70.536.84.483.2]This report presents the phenotypic and genotypic charac-teristics of GAS isolates belonging to 13 newemmsequence types that were first detected among randomly collected Bra-zilian isolates. The data indicate the existence of a significant proportion of newly recognizedemm types with a variety of characteristics, suggesting that strains circulating in Brazil, an area not previously extensively surveyed, may represent a large FIG. 2. Dendrogram resulting from a computer-assisted analysis of the PFGE profiles ofSmaI-digested genomic DNA ofS. pyogenesbelonging to newemmsequence types isolated in Brazil. The scale represents average percentages of similarity.
on May 15, 2020 by guest
pool of M serotypes and clonal groups different from those known to circulate in other geographic areas. Such information can contribute to a better understanding of the local and global dynamics of GAS populations, and of the epidemiological as-pects of GAS infections occurring in tropical regions.
ACKNOWLEDGMENTS
This study was supported in part by Conselho Nacional de Desen-volvimento Cientı´fico e Tecnolo´gico (CNPq), Coordenac¸a˜o de Aper-feic¸oamento de Pessoal de Nı´vel Superior (CAPES), Financiadora de Estudos e Projetos (FINEP), Fundac¸a˜o de Amparo a` Pesquisa do Estado do Rio de Janeiro (FAPERJ), and Ministe´rio da Cieˆncia e Tecnologia (MCT/PRONEX), Brazil.
We thank Theresa Hoenes, Raji Viswanathan, and Zhongya Li for their excellent technical assistance in the CDC Streptococcal Genetics Laboratory. We also thank the CDC Biotechnology Core Facility Branch computing group, most notably Scott Sammons, Elizabeth Neuhaus, and Sarah McKneally, for assistance in constructing and maintaining the web site http://www.cdc.gov/ncidod/biotech/infotech _hp.html.
REFERENCES
1.Beall, B., R. Facklam, T. Hoenes, and B. Schwartz.1997. Survey ofemmgene
sequences and T-antigen types from systemicStreptococcus pyogenes infec-tion isolates collected in San Francisco, California; Atlanta, Georgia; and Connecticut in 1994 and 1995. J. Clin. Microbiol.35:1231–1235.
2.Beall, B., R. Facklam, and T. Thompson.1996. Sequencingemm-specific
PCR products for routine and accurate typing of group A streptococci. J. Clin. Microbiol.34:953–958.
3.Beall, B., G. Gherardi, M. Lovgren, R. R. Facklam, B. A. Forwick, and G. J.
Tyrrell.2000.emmandsofgene sequence variation in relation to serological
typing of opacity-factor-positive group A streptococci. Microbiology146:
1195–1209.
4.Black, C. M., D. F. Talkington, T. O. Messmer, R. R. Facklam, E. Hornes,
and O. Olsvik.1993. Detection of streptococcal pyrogenic exotoxin genes by
a nested polymerase chain reaction. Mol. Cell. Probes7:255–259.
5.Facklam R., B. Beall, A. Efstratiou, V. Fischetti, D. Johnson, E. Kaplan, P.
Kriz, M. Lovgren, D. Martin, B. Schwartz, A. Totolian, A., D. Bessen, S.
Hollingshead, F. Rubin, J. Scott, and G. Tyrrell.1999.emmtyping and
validation of provisional M types for group streptococci. Emerg. Infect. Dis.
5:247–253.
6.Gardiner, D. L., A. M. Goodfellow, D. R. Martin, and K. S. Sriprakash.1998.
Group A streptococcal Vir types are M-protein gene (emm) sequence type specific. J. Clin. Microbiol.36:902–907.
7.Maxted, W. R., J. P. Widdowson, C. A. M. Fraser, L. C. Ball, and D. C. J.
Bassett.1973. The use of serum opacity reaction in the typing of group A
streptococci. J. Med. Microbiol.68:83–90.
8.Moody, M. D., J. Padula, D. Lizana, and C. T. Hall.1965. Epidemiologic
characterization of group A streptococci by T-agglutination and M-precipi-tation tests in the public health laboratory. Health Lab. Sci.2:149–162.
9.National Committee for Clinical Laboratory Standards.1999. Performance
standards for antimicrobial susceptibility testing. Ninth informational sup-plement, M100–S9. National Committee for Clinical Laboratory Standards. Wayne, Pa.
10. Stevens, D. L.2000. Group A beta-hemolytic streptococci: virulence factors,
pathogenesis, and spectrum of clinical infections, p. 19–36.InD. L. Stevens, and E. L. Kaplan (ed.), Streptococcal infections; clinical aspects, microbiol-ogy, and molecular pathogenesis. Oxford University Press, New York, N.Y.
11. Talkington, D. F., B. Schwartz, C. M. Black, J. K. Todd, J. Elliott, R. F.
Breiman, and R. R. Facklam.1993. Association of phenotypic and genotypic
characteristics of invasiveStreptococcus pyogenesisolates with clinical com-ponents of streptococcal toxic shock syndrome. Infect. Immun.61:3369– 3374.
FIG. 3. Dendrogram based upon sequence comparisons of 114 residue N-terminal portions of the various deduced M proteins representing new
emmsequence types first encountered amongS. pyogenesisolated in Brazil (*St211 was an exception within this group of M N-terminal sequences in that the strain harboringst211was OF andsofPCR negative. Note that the branch point forst211is much deeper than the other branch points
in the OF-positive section of the dendrogram).