Differentiation of neuronal cells using a murine embryonic stem cell based method
Full text
Figure
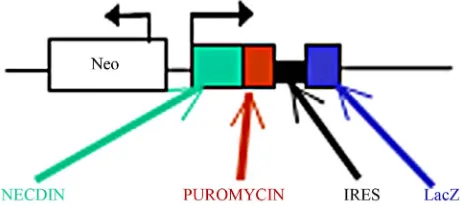
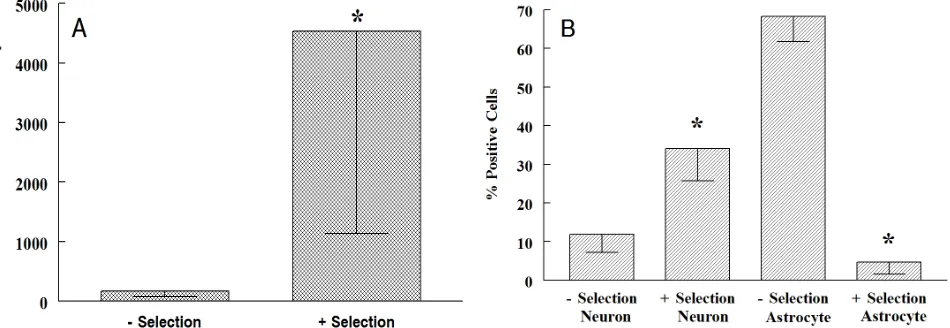
Related documents
However, a Maximum Residual Disinfectant Level Goal (MRDLG) and Maximum Residual Disinfectant Level (MRDL) of 4.0 mg/L was established by US EPA as the enforceable maximum
The present study revealed the qualitative phytochemical analysis and in vitro antioxident activities of ethanolic fruit extract partitioned in different solvents (ethanol,
Chemists use their knowledge of synthesis, decomposition, single replacement, and double replacement to predict what will happen in chemical reactions?. When predicting the
With an objective to find the nexus between nexus between Brand Equity and Consumer’s Purchase intention a theoretical framework has been formed where consumers’ purchase
PUSH and ACK Flood: A PUSH or ACK flood DDoS attack inundates the server with fake PUSH and ACK requests to prevent the server from responding to legitimate traffic.. SYN Flood:
Since dead hosts are unable to mount an inducible immune re- sponse, we reasoned that sexual dimorphism in bacterial growth would be maintained in dead hosts if it was due
Webmasterworld hosts a forum that discusses general domain issues, such as: domains as a brand, domain values, negotiating domain sales, domain inventory management, and
To answer the first research question, the respondents were asked to rank their perceptions of the extent to which they think they had competence in general language knowledge and
