With 12 text-figures Printed in Great Britain
DIFFUSION IN INSECT WJNG MUSCLE,
THE MOST ACTIVE TISSUE KNOWN
BY TORKEL WEIS-FOGH
Zoophysiological Laboratory B, Juliane Maries Vej 36, Copenhagen University, Denmark
(Received 12 July 1963)
INTRODUCTION
During flight, insect wing muscle is a strictly aerobic tissue. Per unit protein it consumes fuel and oxygen at rates paralleled only in freely suspended bacteria but, in contrast to the microbiological systems, this activity takes place in a very concen-centrated and highly organized tissue so that the steady-state rates per unit volume represent an absolute record in biology (see Weis-Fogh, 1961, and Table 1). From the point of view of diffusion, many wing muscles are large and may exceed 10 mm in length and 2 mm. in width. Moreover, in some Diptera the fibres are of giant dimensions. In the very active Rutilia, for instance, the cross-section is 1800 fi long and more than 500 fi wide. The transport of fuel and oxygen from the surroundings to the sites of consumption and the reverse transport of catbon dioxide therefore repre-sent a challenge to the biologist both in relation to transport in the liquid phase and in the intricate system of air tubes, i.e. in the tracheal system.
Since Krogh's studies on tracheal respiration (1920 a, b) it has been quite clear that diffusion in the air tubes plays a major role in the transport of the respiratory gases even over quite long distances, often supplemented by ventilation of the larger trunks or air sacs. Recently, however, Krogh's deductions have been heavily criticized partly on the basis of very detailed measurements of diffusion pathways in silk-worms (Nunome, 1944, 1951) and partly on the basis of a number of simplifications in Krogh's treatment (Buck, 1962). I have found most of these arguments unfounded, as will be shown, but it is true that the number of quantitative concrete examples published is still so limited that there is a danger in relying even upon established qualitative principles. I hope to remedy this situation by analysing the most extreme case in existence.
Fortunately, some wing muscles are constructed according to simple geometrical principles so that it has been possible to reduce the diffusion problems to one-dimen-sional, steady-state equations (Appendix).
MATERIAL AND METHODS
It was then fixed in 4 % formaldehyde for 2 days, washed for 1 day, soaked in 15 % gelatine at 370 C. for 1 day, transferred to 25 % gelatine for 1 day, embedded and fixed in 4 % formaldehyde for 2 days. The preparation was washed briefly before being sectioned on the freezing microtome. The section thickness was adjusted so that it corresponded approximately to the distance between the origins of the secondary tracheae from the primary trunk. The sections were transferred to glycerol-water 1:1 and after evaporation of most of the water, they were mounted in glycerol-gelatine between two cover slips placed on a slide. In this way, distortion and shrinkage was so small that it could be neglected.
THE PROBLEMS
(a) Metabolic rate
When insects do not fly the metabolic rate m usually varies between o-oi and 0-04 ml. O2 per gram body weight per minute so that we may assume that resting wing muscles consume about 0-02 ml. O2/g. muscle/min. During steady-state flight the rates are 50-400 times greater, as it is seen from Table 1. It is apparent that there is no general relationship between size and rate. In the calculated examples, the diffusion equations were therefore solved for m = 1, 2, 4 and 8 ml. Oj/g./min. (and for 0-02, in brackets). The corresponding figures for the consumption of glucose are given in Table 6. The smallest rates are probably representative for insects which do not perform sustained flight but the muscles of most good flyers, whether small or large, require 1*5-3 ml. O2/g./min. Some of the large Hymenoptera and Diptera approach the exceptional rate of 8.
Table 1. Oxygen uptake of insect wing muscle during steady-state flight (For some of the insects, the necessary exchange of air between muscle and surroundings per contraction cycle ( = per wing stroke) has been calculated as a percentage of the muscle volume, assuming that the muscle absorbs 1/4 of the oxygen present in atmospheric air (1/4x21% = S%).)
(ml. O,/g. (100 x ml. muscle/min.) air/g./stroke)
Locust, Schiitocerca, (Weis-Fogh, 1952) Dragonfly, Aethna, (Weis-Fogh, 19646) Butterflies and moths, (Zebe, 1954) Aphid, Aphis (Cockbain, 1961)
Fruit fly, Drotophila, (Chadwick & Gilmour, 1940) Wasp, Vespa erabro, (Weis-Fogh, 1964ft) Blowfly, Lucilia, (Davis & Fraenkel, 1940) Honeybee, ApU mellifera, (Hocking, 1953)
(b) Supplies and supply routes
During a single performance the wing muscles may consume more fuel than that corresponding to the total dry matter of the active fibres so that the stores of glycogen and fat in the muscle may be ignored in this context. The fuel is therefore transported to the muscles by means of the blood which bathes them and consists of carbohydrates, fats or their break-down products (Weis-Fogh, 1952). It is most likely that trehalose in the blood is split to glucose on its passage through the fibre membrane and it is unlikely that the fatty acids are broken down as far as to acetate before they diffuse in,
1 -4-2-8
i-8
1-4-3-5 1-4-1-8 2-0-2-3 2-6-3-3 5-6
7-3
2-6-S-2 2 — — 0-3-0-4 0-9-1-1
because this would mean a great waste of energy. We may therefore choose glucose as a model substance for the diffusion of fuels.
A typical wing muscle receives its supply of air from a longitudinal, primary trachea or air sac which is directly connected with a spiracle (Weis-Fogh, 1964a). From the primary supply, a system of secondary tracheae branch off transversely between the fibres and give off tertiary tracheae at regular intervals (see dragonfly muscle) which eventually split up into tracheoles some of which may become ' internal' by identing the fibre membrane (Edwards, Ruska & de Harven, 1958; Vogell, Bishai, Busher, Klingenberg, Pette & Zehe, 1959; Smith, 1961a). A wing muscle is a prismatic structure of parallel fibres and its total volume consisting of fibres, blood and air tubes therefore decreases with shortening by a fraction corresponding to the relative shorten-ing. This must lead to muscular pumping of blood and air in and out. In the second column of Table 1, the amount of fresh air is calculated which is needed per stroke if 25 % of its original oxygen content is used up. In the different insects this amount is of the same magnitude as the change in volume per stroke or even larger, and only an intricate system of unidirectional valves in the primary tracheae could ensure an adequate supply by muscular pumping. Such valves are not present, however, and in large insects morphological inspection and direct experimental evidence demonstrate that the primary supply routes are ventilated strongly and in such a way that the com-position of the air is almost the same as in the major air sacs, in locusts about 5 % CO2 and 15 % Oa (Weis-Fogh, 1964 a, b). The pumping therefore causes blood movements and, as will be indicated, some small ventilation of the secondary tracheae which, how-ever, is not essential but leads to a greater exchange of blood between the interior and the exterior of the muscle.
The problems are then confined mainly to (a) the exchange of fuel between blood and tissue and (b) to the exchange by diffusion of O2 and CO2 between the primary tubes and the sites of combustion. The transport across fibre membranes and tracheolar walls will not be dealt with.
(c) On diffusion
We are dealing only with diffusion in the stationary or steady state where there is no change in concentration at any point in the system. In a mixture of substances the net transport J of a given substance passing perpendicularly through a unit reference area per unit time is given by Fick's first law.
where c is the concentration of the substance, x the co-ordinate normal to the reference surface, and D the diffusion coefficient for the substance in the given system. In the c.G.s. units it has the dimensions of cm.2 sec."1. In most biological systems D is not known but only the permeability constant P which is the flow of a substance through unit area per unit time when the average concentration gradient is unity and when the flow is measured in the steady state under certain conditions and with the assumption that D remains independent of c (cf. Barrer, 1941; Jost, 1952).
gas at N.T.P. dissolved per unit volume of liquid is V = apj'760, where a. is the absorp-tion coefficient. The volume of gas at N.T.P. transported according to equation (1) is
The diffusion coefficient of CO2 is about 15 % lower than for O2 in most systems but since the absorption coefficient for C0a in pure water is 28 times greater than that of O2> the permeability constant P for COa has been found to be about 36 times greater than for oxygen in animal tissues which usually absorb more CO2 than water (Krogh, 1919). For the same difference in partial pressure Ap, CO2 is then transported about 36 times quicker in tissues, which we shall call tissue diffusion, than Ot but 15 % slower
in the tracheal system, which we shall call air-tube diffusion.
The permeability constant P for 02 in air at 200 C. and atmospheric pressure is 11 ml. min."1 cm.~2 atm."1 cm."1, while it is 3-4 x io"6 in water and 1-^x io"6 in muscle from the abdominal wall of the frog (Krogh, 1919). Like insect wing muscle, frog muscle is poorly vascularized and contains little myoglobin so that the facilitating effect on O2-diffusion of high concentrations of haemoglobin and similar oxygen carriers should be absent (cf. Scholander, i960; Hemmingsen & Scholander, i960). Moreover, a strictly aerobic muscle can hardly afford to decrease the oxygen tension to the low values necessary for this facilitated transport to be operative. For a given Ap tissue diffusion then requires io6 times more area than air-tube diffusion in order to transport a given amount of oxygen. It is therefore convenient to introduce the hole fraction a as the summed cross-sectional area of all air tubes in a given tissue surface divided by the total area of that surface. In insect wing muscle, a usually ranges between io"1 and io"3 (see later) which means that the permeability for O2 is io8 to io6 times greater than in non-tracheated muscle and for CO2, 50-5000 times greater.
The permeability constant for glucose in muscle is not known but if we adopt the diffusion coefficient D in water (0-57 x io"6 cm.2 sec."1; see Jost, 1952), we shall certainly not overestimate the exchange but rather underestimate the necessary con-centration difference by a factor of two or more.
DRAGONFLY WING MUSCLE
important feature is that the secondary air tubes radiate out from the primary tubes in an exceedingly regular fashion. Thus, for every 20-25 fi along the axis each lobe receives one secondary trachea which enters the central edge of the lobe and runs almost perpendicular to the fibres towards the peripheral edge, branching only in the transverse plane to form a structure resembling the fruit grower's 'espalier tree'. Moreover, at different levels the ' espalier trees' are almost identical in shape and they branch between the same fibres so that it is possible to reconstruct the tracheal supply in detail from rdktively few transverse sections.
0-5 1-0
Fig. 1 Fig. 3
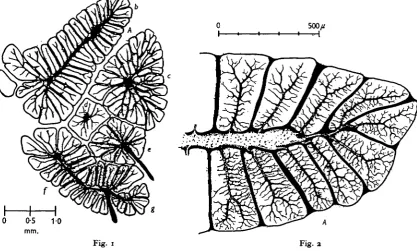
Fig. 1. Transverse section of the dorso-ventral wing muscles in the left half of the metathorax of a large dragonfly Aeshna cyanea. The injected tracheae are drawn in black. Nomenclature according to Neville (i960); (a) first basalar muscle, (6) second basalar, <c) tergosternal, (d) anterior coxoalar, (e) posterior coxoalar, if) first subalar, and (g) second subalar muscle. Lobe A in Figs. 2 and 4 is indicated.
Fig, 2. Part of the second basalar wing muscle in Fig. 1. The blood-filled spaces are in black and the injected trachea* are faintly dotted.
From, Figs. 1-3, it is seen that in some muscles the distance from the primary trachea to the periphery may approach 1 mm. On the other hand, the blood (black in Figs. 2 and 3) bathes the lobes completely and no point is more than 75 ft from a free blood pool. As to volumes, 75 % of the cross-section in Fig, 2 corresponds to fibres and 68% in Fig. 3, The blood occupies 12 and 18 %, the secondary air tubes about 8 and 10% and the primary tubes 5 and 4 %, respectively. More than half the volume of the clefts through which the trunks of the ' espalier tree' run consists of air tubes and the rest is blood. The blood then constitutes 15-20% of the total muscle volume.
[image:5.451.19.437.185.436.2]234
[image:6.451.135.318.116.392.2]and the two large radial lobes B and C in Fig. 3. For each step we shall try to generalize the results obtained so as to facilitate the comparison with other muscles. During flight the oxygen uptake is i-8 ml. Og/g./min.
Fig. 3. Transverse section of the tergosternal muscle from the left mesothorax of Aeskna cyanea, showing lobes B and C. The blood-filled spaces are in black and the injected tracheae are faintly dotted. The circles indicate the estimated equivalent diameter of the ventilated primary supply tubes for the two lobes in question. Further distally, the transport of re-spiratory gases takes place only by diffusion.
(a) Diffusion in primary tracheae
In small insects with no ventilation during flight, the centro-radial type would be adequate only in. a thorax less than 1 mm. high and only at moderate metabolic rates. In the latero-linear case, however, where the primary supply is an air sac which spreads out over the surface of the muscle and has a cross-sectional area similar to that of the muscle, diffusion is adequate up to the size of a locust, as is probably the case with the controller-depressor muscles of Schistocerca (Weis-Fogh, 1964a).
Table 2. The maximum distance L (in /i) for the diffusion of oxygen in the longitudinal
direction of the axially placed primary tube of a centro-radial type muscle, according to Fig. 12 D and equation (7) when Ap = 0-05 atm.
Active
Rest , * > Metabolic rate m (ml. Ot/g./min.)... 002 1 2 4 8
T = R/io (7>4°o)
(14.800) 1,050 2,100
740 1,480
520
1,040
37O 74O
In the latero-linear type of muscle whose primary supply may have a cross-sectional area similar to that of the muscle, these distances should be multiplied by 5 to 10, cf. Drosopfula on p. 248.
The necessity for ample ventilation of the primary tracheae in the centro-radial type of muscle will be apparent also from the analysis of gas diffusion in the subsequent branches of the tracheal tree.
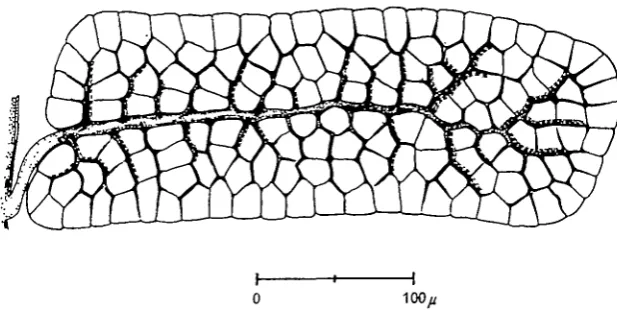
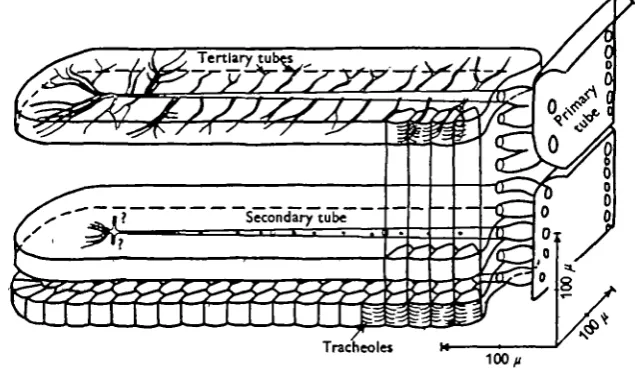
Fig. 4. Transverse section of lobe A in Figs. 1 and 2. The thickness is 29 ft. The primary tube is indicated at the extreme left, the tapering secondary tube runs horizontally, and the tertiary tubes branch off in the vertical direction, ending in tracheoles. Note the close packing of the fibres. The broken lines indicate the number of tertiary branches in the section in addition to the top branch.
(b) Air-tube diffusion in the small lobe A
3
[image:8.451.68.386.188.372.2]lobe. These long tapering tubes follow a longitudinal cleft which occupies 3 % of the volume. One secondary tube is given off for every 21 /i on the average. The diameters were measured in 17 sections at eight relative distances £ from the entrance ( x 800 linear magnification, eye-piece 9crew micrometer). Fig. 6 shows that the diameter of the secondary tubes decreases linearly from the entrance (£ = o) to the tip (£ = i-o). However, at £ = 0-84, detailed measurements of the tertiary system under oil immer-sion ( x 1600) showed that each secondary tube ends and continues as three tertiary tubes each of which soon branch into two, giving rise to an increase in equivalent
Fig. s. Reconstruction of the tracheal supply to a lobe of a wing muscle in a dragonfly (lobe A in Figs, i, 2 and 4), as explained in the text.
Diffusion in the gas phase may now be divided into two steps. For both we choose
m = 2 ml. Oa/g./min.
Diffusion in secondary tubes. The length L of the lobe is 407 x io~*cm. and the
hole fraction a = 1-5 x io~2, calculated as the summed cross-section area of the secondary tubes at their entrance into the lobe divided by the corresponding area of the lobe perpendicular to the tubes. Since the cross-section of the lobe is approxi-mately rectangular, the necessary difference in partial pressure A/> was calculated for oxygen according to equation (6). The upper curve in Fig. 6 shows that from the entrance at £ => o to where the tertiary tubes begin at £ = 0-84, diffusion is adequate if A/> ^ 0-035 a t m- Ventilation due to muscular pumping of the sac-like parts between the lobe and the primary tubes will increase the effective diameter of the central tubes but neither this nor some ventilation of the proximal part of the secondary tubes will have much effect, Ap being so small.
to
Q
004
002
6
4
2
i - i
-" . . o
-1 -1
1 1 1 1 1
Secondary tube
1 1 1 1 1
1
o
•
Tertiary tubes
1
X
X
0-2 0-4 0-6
Relative distance
[image:9.451.67.381.246.513.2]0-8 1 0 = £
Fig. 6. Diameter of the secondary tube in lobe A (lower curve) and the necessary difference in partial pressure Ap of oxygen (upper curve) as functions of the relative distance £ from the primary tube.
Judged merely from their appearance, tracheal systems with linearly decreasing diameter like that in Fig. 6 are probably quite common, but so are systems with con-stant diffusion area (Krogh, 1920a, for Cossus larva; Thorpe & Crisp, 1947, for
Apheiocheirus; Locke, 1958, for Rhodnius). In Table 3, I have therefore calculated
IO~2, 0-5 mm. is about the limit. The small value of io~3 is relevant for the next step and also for diffusion in dipteran muscle fibres.
Diffusion in terminal tubes. The depth of tissue to be served at either side of the
median cleft is 65 fi but, because of branching, we shall use 75 /i. The hole fraction of the tertiary tubes at the cleft is a = i-o x io~3i According to equations (4), (5) and (6) Ap is 0-005 atm. in case of constant diffusion area, which is the most likely, o-oi atm. in case of linearly decreasing area and 0-023 a t m- m the quite unrealistic case of linearly decreasing diameter. This step is therefore particularly favourable for diffusion.
[image:10.451.73.393.306.427.2]As far as the terminal part of the lobe is concerned, these low figures for Ap were checked by considering the end of the lobe as a half cylinder with tubes radiating out from the axis towards the periphery (Fig. 12 J and equations (16) and (17)). The hole area a was 3-5 x io"3 and the resulting Ap was 0-003 a t m- f°r a constant diffusion area and 0-007 for a linearly decreasing area.
Table 3. Maximum distance {in fi)for the diffusion of oxygen in air tubes of wing muscles
supplied according to the latero-Unear type and when Ap = 0-05 atm.
(In (A) the diffusion area of the air tubes is constant, Fig. 12 B and equation (4), and in (B) the diffusion area decreases linearly with the distance from the surface of the muscle, equation (5)-)
Active Rest ,
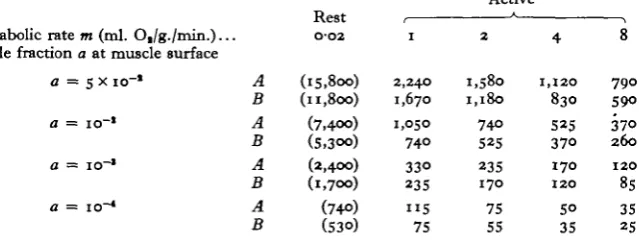
Metabolic rate m (ml. O,/g./min.)... Hole fraction a at muscle surface
a = s x io"1 A B A B A B A B O'O2 (I5,8oo) (11,800) (7,4OO) (5,300) (2,4OO) (l,7OO) (740) (530) I 2,240 1,670 1,050 74O 33O 235 " 5 75 2 1,580 1,180 74° 525 235 170 75 55 4 1,120 830 525 370 170 120 50 35 8 790 59O 37O 260 120 85 35 25
In other words, it is only necessary to establish a difference of less than 0-05 atm. in the partial pressure between the primary supply and the terminal end of the tracheoles in order that diffusion alone may account both for the inwards transport of oxygen and for the outwards transport of carbon dioxide.
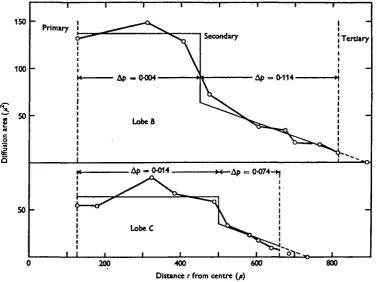
(c) Air-tube diffusion in lobes B and C
and tracheolar tubes (the third step). According to equation ( n ) , the first step with constant area requires that A/> is only 0-004 atm. in lobe B (a = 6-6 x io~2) and 0-014 in lobe C (a = 2-8 x io~J). This means that ventilation by muscular pumping of these partially exposed, wide tracheae should have little significance for the transport of oxygen but may be of some consequence for the transport of carbon dioxide by means of blood. The second step, however, is critical since Ap is 0-114 atm. in lobe B and 0-074 m l°be C (equation (136)), a being 8-8 x io"3 and 3-9 x io"3 respectively. In other words, the drop in partial pressure necessary is 0-12 atm. in B and 0-09 atm. in C. If we adopt the results from lobe A, the total drop in tension in the air tubes from the primary supply to the fibre surface would then amount to as much as 0-13 and 010 atm. respectively. There is no obvious possibility for any improvement because the main diffusion resistance is located in the peripheral thin tubes.
150
100
SO
50
Primary
1 Tertiary
400
Distance r from centre (p)
[image:11.451.39.417.223.505.2]600 800
Fig. 7. Summed cross-sectional area of the tracheal tubes in lobes B and C of Fig. 3, as a function of the distance from the equivalent axes of the primary tube. The thin continuous lines show the variation of the area adopted for calculating Ap.
Taking account of tissue diffusion, the total difference in partial pressure from the primary tube to the sites of combustion must be greater by about 0-05 atm. which means that these muscles cannot become larger unless the tracheal system is re-organized and, also, that the central trachea must be ventilated very efficiently. In
Schistocerca the air in the primary trunks contains about 15% O2 during flight but this is close to the possible limit in dragonflies since 13 % may be needed to ensure an adequate exchange in certain lobes. We must therefore expect that the dragonfly
thorax is ventilated more strongly than the locust thorax, as is also borne out by measurements (Weis-Fogh, 1964 ft). Moreover, the carbon dioxide tension in the interior of dragonfly muscle fibres must be high during flight so that transport by blood may be more important than in locusts. This is considered a more probable explanation than cooling for the synchronized movements of the thoracic spiracles after cessation of flight (Miller, 1962) until the tension has decreased to a low value again.
Table 4. Maximum radius R (in fi) for the diffusion of oxygen in the radiating air tubes
of a cylindrical, tracheated muscle of the centro-radial type (Fig. 12H, I)
(The radius of the primary trachea is r0 = Rjio and the possible drop in oxygen tension in the radiating secondary tracheae is Ap = 0-05 atm. The diffusion area decreases linearly with the distance from the axis (equation (13a)) but if it is considered constant as in equation (11), all distances should be increased by about 55 %).
Active
Rest , • , Metabolic rate m (ml. OJg./min.)... 0-02 1 2 4 8
Hole fraction a of wall of primary trachea
a = io"1 (6300) 890 630 440 310
a = 5 x 10"1 (4400) 630 440 310 220
a = io~* (2000) 280 200 140 100
Since the centro-radial type of muscle is so common among large insects, Table 4 was computed and it is seen that even at medium rates with a large number of holes in the central tube, such a muscle may hardly exceed 1 mm. in diameter unless, as in dragonflies, the primary system is exceedingly well ventilated with the consequent risk of an impaired water balance. If a of the central tube is as large as o-i, the maximum possible ratio / between tube and muscle diameter can be estimated from equations (12) and (14). In order that the central tube should not occupy too large a volume, we shall choose / = 0*2 as the upper limit. The maximum diameter for such muscles is then 2-4 mm. when the consumption is 2 ml. Og/g./min. but decreases to 1-2 mm. at the higher rates. The dimensions of insect wing muscles are consistent with this estimate except for those of the giant belostomid bug Lethocerus uhleri. According to Moller (1921), the tracheal supply is similar to that of dragonfly muscles but the diameter sometimes exceeds 5 mm. However, the secondary radiating tracheae do not taper. On the contrary, they expand into terminal air sacs when they reach the periphery, covering the surface like intestinal villi. In such exceptional cases the secondary tracheae are undoubtedly ventilated.
The above examples stress that the diffusion area of tracheal systems often varies in a complicated way with distance, as was also found by Nunome (1944), so that it must be analysed in every case considered.
(d) Tissue diffusion
It is characteristic of dragonfly wing muscle that the fibres of a lobe adhere closely to one another so that the diffusion of respiratory gases and of fuels must be treated separately.
Dissolved gases in dragonfly muscle. The tracheoles do not indent the fibre membrane
equation (18) in Thorpe & Crisp (1947), the dense meshwork of tracheoles makes it reasonable to use the more simple expression of equation (96), i.e. to consider the fibres as being completely surrounded by a layer of air. In this way we obtain the maximum possible radius R for a fibre seen in Table 5. Since the average cross-sectional area of a dragonfly fibre was 350 /J2, the maximum diffusion distance is about 10 /i, i.e. close to the possible limit when m = 2 ml. Oj/g./min. and Ap = 0-05 arm. We saw from the previous examples that Ap cannot be significantly greater than this value and, without internal tracheoles, the fibres of most insect wing muscles must be less than 20 fi in diameter. It is possible, however, that the arrangement of the dragonfly fibre into alternating radiating lamellae of ribbon-like fibrils and flattened sarcosomes (Smith, 1961 b) is an adaptation to this situation since surfaces of equal oxygen tension become crenelated rather than cylindrical with valleys at the non-consuming fibrils and crests where the consuming sarcosomes are located. Since the necessary tension usually depends on the second power of the distance to be served, this tends to facilitate exchange by diffusion and such measures could be expected to have evolved in a system pressed to the limit (see also cockroaches in the Discussion).
Table 5. Maximum distance (in n)for the diffusion of oxygen in the dissolved state from
a source into active and resting muscle tissue {figures in brackets) when Ap = 0-05 atm.
((A) distance L for rectilinear diffusion from a flat surface, Fig. 12 A and equation (3). (£)
radius -R for diffusion from the surface of a cylinder to the axis, Fig. 13 F and equation (96). (C) R — r0 for radial diffusion from an air tube of diameter 1 /* (r0 = 0-5 /*) into the tissue
cylinder surrounding it, Fig. 12 E and equation (8).)
Active
metabolic rate m Rest , * \
(ml. Oj/g./min.)... 0-02 1 2 4 8
A: L(ji) (84) 11-8 8-4 5'9 4 2 B: R(ji) (118) 167 n-8 84 59
C: R-ro(]i) (41) 7-4 5 4 4-6 3-6
General comments on tissue diffusion of gas. Table 5 also illustrates some general
properties of tracheal systems. In his recent review, Buck (1962) claims that if simple diffusion were to account for the transport the 'mean distance from tracheole wall to cytoplasmic sites of Oa consumption cannot exceed o-z/i' under any conceivable condition even when Ap = 0-2 atm. He uses this and other arguments to show that there must be a considerable diffusion out from the larger tracheal trunks and also to cast doubt upon the principles of simple diffusion as the main transport mechanism for gas in tracheated tissues.
However, he does not disclose his method of calculation and Table 5 clearly demonstrates that his conclusions are untenable even in the most active tissue known. In example A, oxygen diffuses in freely from the flat surface and it is seen that when Ap = 0-05 atm. a sheath of active muscle will be served which is 4-12 fi thick. This illustrates how far out from a large tube direct diffusion is sufficient (still far in excess of 0-2 fi) and it is obvious that large air tubes are completely inadequate because they cannot come close enough to the sites of consumption unless they occupy the major part of the muscle volume. In contrast, example C shows that under similar con-ditions a tube only 1 fi in diameter can serve a cylinder, which is 7-15 ft in diameter,
of which 2-6 % of the volume is air. In other words, if, as we have seen, exchange of air within the tube system is sufficient to permit a difference in oxygen tension between the tracheolea and the sites of combustion of maximally 0-05 atm. the distance between the tracheoles need not be less than about 6—8 fi in a block of the most active muscle tissue. In many muscles it is about 3 fi, as discussed later, so that the safety factor is 2-3. It is also clear that during evolution flying insects must either have decreased the fibre diameter considerably (example B), which is not the case, or they must have 'invented* indenting tracheoles, which is the case. The wall thickness of tracheoles amounts to only 100-300 A and has therefore not been taken into account (cf. Smith,
1961a).
[image:14.451.45.402.382.475.2]Since the permeability constant of carbon dioxide in animal tissue is about 36 times greater than for oxygen, all distances in Table 5 should be increased by a factor of 36^ = 6. There is therefore no problem with respect to tissue diffusion. As to diffusion in the tracheoles and in the tertiary tubes, tissue diffusion clearly represents a shunt but it will not be very effective because of the relatively large hole fraction invariably found in wing muscle. The main problem concerns exchange in the secondary system which becomes the major factor determining the carbon dioxide tension in the tissue, as already discussed.
Table 6. Diffusion of glucose from the surface of a cylindrical muscle fibre to the axis for
various values of its radius R
(The figures show the minimum difference in concentration At between surface and axis in mg. glucose per ioo g. muscle (mg. %) and are based on diffusion in pure water, according to equation (96). The corresponding figures for a flat sheet of tissue supplied from the one side and whose thickness L equals it are twice as big, according to equation (3).)
Active Metabolic rate m
in mL Ot/g./rrun.... in mg. glu./g./sec
R = 500/* R = 200/1
R = 100/*
R = 50/* R = 20/* R = 10/*
Rest
0-02 4-S x io-«
(49) (08) (02) (005) (0008) (0-002) 1 2-25 x io~*
MS 4 0 1 0
2-s
0 - 4 O-I
a
4-5 x 10-'
4 9 5
79
2 0 S O
o-8
0 - 2
4
9 x 10-1
9 9 0 158 4 0 9 9 1-6 0-4 8
18 x i o - '
1980 3 1 6
79
2 0 3-2 0 8
Diffusion and fuel concentration in various muscles. In dragonflies there is no effective
that c must be about 79/0-41 === 200 mg. %. In the very active tachinid fly Rutilia
potinia, the distance from the membrane to the central parts of the flat fibre (cf.
equation (3)) is about 260 ji (Tiegs, 1955), giving the exceptionally high value of Ac = 535/0-41 === 1300 mg. % for a metabolic rate of only 4 ml. Og/g./min. In Diptera with such giant fibres, but not in Hymenoptera, recent investigations indicate that the plasma membrane is invaginated into deep narrow longitudinal clefts (Auber, 1961; D. S. Smith, personal communication) so that some exchange takes place by muscular pumping (see below). But in most flying insects, there is no doubt that the difference in fuel concentration inside the fibres must be very high since mixing of the sarcoplasm of these close-packed parallel systems is unlikely in view of the small degree of shortening (0-5-1 %).
In this context, the exact nature of the fuel is of little significance. It could be argued that, although Ac between plasma membrane and fibre axis is large, the blood concentration need not be particularly high since, for example, glucose may become phosphorylated in passing the membrane and thus being prevented from diffusing back. This could hardly be so in Aeskna muscle because the fuel must diffuse both in and out of the peripheral fibres in order to reach the central ones, indicating that it permeates the membranes freely. Consequently, the fuel concentration in the blood must be considerable. There is, however, a more compelling reason why flying insects should contain large amounts of trehalose and other fuels dissolved in the haemolymph.
Muscular pumping and fuel concentration. In many insects the wing muscles are so
large that diffusion is quite inadequate for transport from the surface to the interior, as is seen from Table 6. The only other mechanism for exchange is muscular pumping but, as an example will show and because of the inefficiency of such a system, the concentration of fuel in the blood must be increased much above the level necessary for diffusion into active fibres. The bee muscle, for instance, consumes 18 mg. glucose per 100 g. muscle per second and if 20% of the volume is blood this means 90 mg. consumed per second per 100 ml. blood entrapped in the muscle. The muscle shortens about 200 times per second but only by 0-5 % of its length, giving rise to a pumping of maximally 100 ml. per 100 g. muscle per second. Under ideal conditions, the blood surrounding the muscle must therefore contain at least 90 mg. % more glucose than the amount necessary for simple diffusion into the fibres. However, muscular pumping moves air as well as blood and its tidal character will make it a very ineffective mechanism when, as here, the relative stroke volume is considerably smaller (40 times) than the relative amounts of air and blood entrapped. We can there-fore conclude that the maximum concentration in Apis blood must be many hundred mg. %, in accordance with measurements of blood sugar (2% blood sugar or more, Beutler, 1937).
244
to flight. It also follows from all that has been said that the main function of muscular pumping is to move blood and not air in contrast to what I thought earlier (Weis-Fogh, 1956). The limited solubility of lipids presents a special problem which is discussed on p. 247.
DISCUSSION AND SOME COMPARATIVE RESULTS
In connexion with the different steps in the transport processes in dragonfly muscle we have already discussed several problems of general interest and illustrated them by means of Tables 2-6, but a more comprehensive and comparative survey remains.
Visible end of tracheoles 160
120
80
-40
o
i
a
10 15 20 25
[image:16.451.58.398.193.412.2]Distance from splrade (arbitrary units)
Fig. 8. The variation of the summed cross-sectional area of the tracheae in a segment of a "ilk worm as a function of the distance from the spiracle (open circles), according to Nunome (1944). The filled circles indicate the diameter of the equivalent air tube.
(a) Diffusion in insect larvae
(b) Diffusion in locust wing muscles
Gas. The tracheal supply of locust muscle is exceedingly rich but much less regular
than in dragonflies so that exact estimates are difficult to make. As an example, we shall consider the large dorsal longitudinal wing muscle. In the mesothorax it belongs to the latero-linear type and, according to fig. 8 in Weis-Fogh (1964a), the hole fraction a at the surface of the muscle is as large as 0-07. The mesothoracic muscle is supplied in a similar way although the primary tubes consist mainly of large parallel tracheae on the mesial surface. In transverse sections (Fig. 9) it is seen that the fibres are much more loosely packed than in the dragonfly muscles so that more than half their surface area is in direct contact with the haemolymph (Fig. 10A). All major clefts and most of the minor ones are packed with branching tracheae (not drawn). The summed area of the secondary tubes therefore seems to remain more or less constant with the distance from the mesial surface of the muscle. Even in the case of a linearly decreasing area, however, Table 3 shows that for a = 0-05 diffusion in the secondary, system is sufficient.
Fig. 9. Cross-section of the dorsal-longitudinal wing muscle in the metathroax of the locust Schistocerca gregaria (a) being the mesial and (b) the lateral side. Only the major tracheae are shown. Note the abundant system of crevices filled with blood and tracheae.
actual case 1-5 x io~*). It was assumed that the entering tracheoles are o-Zfi wide and that they taper gradually towards the interior. There is therefore no problem with respect to the diffusion of gases. In Chortoicetes terminifera which is also a good flyer, Tiegs (1955) found the tracheoles arranged as in Schistocerca and, according to his photographs figs. 125-126, with approximately the same density.
1001
I I I I I I
[image:18.451.92.363.144.357.2]0 SO//
Fig. 10. Cross-sections of fibres of the dorsal-longitudinal wing muscle of the locust
Schisto-cerca gregaria. (A) shows the loose packing of a group of fibres and (B) shows the number of
tracheoles which indent a fibre section 25 /* tall.
Table 7. Calculated radius R (in fi) of a cylindrical muscle fibre supplied with oxygen by
diffusion in internal tracheoles which radiate from surface towards axis with linearly decreasing cross-sectional area (Fig. 12G and equation (10)
(Ap = 0-05 atm. Compare R in Table 5, case B.)
Active
Metabolic rate m (ml. O,/g./min)..
Hole fraction a at fibre surface
Rest
O-O2
(10400) (3300) (1050)
1480 470 150
1050 320 105
740 230 75
520 165 5°
Fuels. As has already been discussed, the transport of fuel to the interior of muscles
[image:18.451.55.394.448.526.2]meta-bolism, since fatty acids consist of large molecules of limited solubility in haemolymph and fibre plasma. If the fibres are about 50 /i in diameter and if the rather large mole-cular weight of fatty acids compared with that of glucose is outweighed, as far as dif-fusion speed is concerned, by the higher caloric value, Table 6 shows that the con-centration difference between the plasma membrane and the axis need amount only to 1 mg. % in locusts (diameter 50 fi), 5-20 mg. % in most butterflies (George & Bhakthan, i960; 80-180 fi) and 5-10 mg. % in some beetles (Darwin & Pringle, 1959; 80-140 /i). The solubility of stearic acid in water corresponds to about 30 mg. % at 250 C. and increases with temperature to 100 mg. % at 370 C. (Handbook of Physics and Chemistry, 42nd ed.). There is therefore no difficulty in providing the individual fibres with sufficient fuel and since Tietz (1962) found that the haemolymph of resting locusts (Locusta) contains 800 mg. % esterified fatty acids (mainly in combina-tion with protein) and 2-6 mg. % free fatty acid, diffusion and muscular pumping together may account for the transport of even long-chain fatty acids. Moreover, in wing muscles of Lepidoptera and Coleoptera there are many fat-storing cells located between the fibres. They possess a considerable capability for fat synthesis (Zebe, 1958) and also a high lipase activity (George & Bhakthan, i960).
0 0-5 mm.
1 I I I I I
Anastomosis 1
Head Abdomen
Leg1.
Leg
3-Anastomosis 2. Leg 2.
[image:19.451.38.416.296.451.2]B
Fig. 11. The major thoracic air sacs and tracheae of Drosopfrila melanogtuter drawn from the original injection preparations of Wigglesworth (1950). (A) a young adult prior to the ex-pansion of the air sacs and (B) an adult after exex-pansion. In contrast to locusts there are two large anastomoses between the right and the left side of the thorax, as indicated.
(c) Diffusion in Drosophila
Ventilatory movements have not been recorded in these small insects either at rest or during flight although C M . Williams (unpublished) saw small rhythmic pulsations of the dorsal air sacs at a frequency unrelated to the wing movements (quoted by Chadwick, 1953). Chadwick & Gilmour (1940) clearly demonstrated that the flight of
D. repleta does not involve an oxygen debt and that the performance can go on at
tensions corresponding to 6 % oxygen in the ambient air. It is therefore almost certain that diffusion accounts fully for the exchange of gases in such small insects even during flight, as already suggested by Krogh, (1920a). In order further to test this, Prof. V. B. Wigglesworth kindly lent me some of his injection preparations of D.
expanded and appear flattened while Fig. 11B is a fully developed female in which the air sacs are greatly enlarged and expanded (Wigglesworth, 1950) to an extent which, as to general organization and magnitude, is reminiscent of the intricate system found in locusts (Weis-Fogh, 1964a). Although more exact estimates are lacking, a com-parison between Fig. 11 and Tables 2 and 3 (m = 2 ml. O2/g./min.) indicates that with such large primary and secondary supplies diffusion should be completely adequate in itself, the total diffusion distance from spiracle to fibre being less than 0-5 mm. throughout the thorax. As to the tracheoles, I have counted the density in a dorso-ventral muscle at different levels (cf. Wigglesworth, 1950, plate II, fig. 2, 75 ft thick section) and found that the average cross-section area supplied by a tracheole is 55 p? corresponding to a radius of 4-2 fi, i.e. somewhat smaller than the figure of 5-4 ft in Table 5C. Since it is unlikely that all existing tracheoles have been successfully injected or counted, the combined results strongly indicate that the entire gaseous exchange can take place by diffusion during rest as well as during flight.
The question remains why such small insects have air sacs. Wigglesworth has recently suggested (1963) that the expanding air sacs tend to reduce the volume of blood to about one-third and therefore to reduce the work of circulation as well as the total amount of dissolved blood sugar without decreasing its concentration. To this should be added the advantage of flexible air sacs with respect to a distributed and thorough mixing of the blood within the thorax and inside the muscles, as discussed in the present paper. Although the flight movements of Diptera hardly result in any net change of thoracic volume, they must give rise to pressure changes and some move-ment of blood and air within the thorax. While the air movemove-ments are of little functional significance, the blood movements may be essential for an adequate supply of fuel. The rhythmic pulsations of the dorsal air sacs observed by Williams may then reflect the work of the aorta and the accessory pulsating organs.
[d) Other vmg muscles
Many data on wing-muscle tracheation offered by Tiegs (1955) are relevant in the present context.
Like dragonflies, the cockroach Periplaneta americana has radial lamellar fibres 30-60 ji in diameter and with no trace of internal tracheoles, but the thickness of the cortical part containing sarcosomes and fibrils is only 10-15 /*• According to Polacek & Kubista (1960) this species can perform flight of some minutes' duration and with an oxygen consumption as high as 4 ml. Og/g. muscle/min. However, unless the lamellar arrangement offers some special advantage as to diffusion (as suggested for dragonfly muscle on p. 241), Table 5 shows that the limiting thickness is 8-4/* for
Ap = 0-05 arm. In other words, the system is working near to the limit. This
accords well with Polacek & Kubista's (i960) observation that pyruvate (and probably glycerophosphate) accumulates in the muscles and the insect soon becomes exhausted in contrast to locusts and other good flyers.
{Table 5). In the less active jassid Erytkroneura ix the many internal tracheoles follow a longitudinal path each supplying about 80 /i2 of the fibre cross section which should permit metabolic rates of about 2 ml. Og/g./min. (Table 5).
The most interesting group consists of the large-fibred Diptera already discussed in connexion with the diffusion of dissolved fuel (p. 243). Here the fibres are served by a regular system of small transverse tracheae about 2 /i in diameter at the surface, tapering and giving off tracheolar side branches in the interior. At the surface the distances between tracheae vary from 30-40 fi in Musca and CalHphora to 20 /i in
Trichophthalma. They serve a depth of 100 and 50 /i respectively. Thus the supply
is of the latero-linear type with a hole fraction of about io~a which is sufficient for intensely working fibres which are two to four times thicker than the fibres found in these insects (Table 3). The tracheoles form a space network (although actual anastomoses may not occur) with a distance between the tubes of from 2-3 fi
{Calliphora, Gastrophilus, Trichophthalma) to 3-5 fi (Musca) and this will ensure
adequate tissue diffusion at the highest metabolic rates encountered as yet (Table 5), the safety factor being of the order of 2-3.
(e) General
So far, all estimates are based on the assumption that diffusion in air is not in-fluenced by the proximity of solid walls (normal diffusion). However, the diameter of the smallest tracheoles, o-i /i, approaches the mean free path of most gas molecules at atmospheric pressure, about 0-06 fi, so that it is reasonable to discuss the possible •effects of the small dimensions. I have been unable to find any literature bearing on my particular problem since the so-called Knudsen flow is related to pore diameters approaching those of the molecules themselves, i.e. smaller by at least two orders of magnitude (cf. Hewitt & Sharratt, 1963). According to discussions with Prof. J. Kofoed and Dr Sten-Knudsen it seems more likely that the diffusion is unhampered because gas molecules adsorbed to the walls should behave as perfectly elastic bodies and be replaced almost instantaneously if removed. In fact, the replacement may take place by lateral displacement within the adsorbed layer at speeds which in some systems may approach that of sound. As far as tracheoles are concerned, Prof. Kofoed sug-gested that, due to this effect, the transport may even be facilitated to some extent compared with diffusion through wider tubes of similar cross-sectional area.
The conclusions drawn in this paper should therefore not be significantly influenced by the smallness of the tracheolar tubes.
CONCLUSIONS
Transport of respiratory gases and of fuel in insect wing muscle depends on three <lifferent mechanisms: ventilatory movements executed by thorax and abdomen, muscular pumping due to length changes, and diffusion. The first is analysed elsewhere (Weis-Fogh, 1964 a, b) while the other two are treated here.
(1) The arrangement of muscle fibres, blood spaces and air tubes was found to be sufficiently regular in dragonflies (Aeshna spp.) to make possible a detailed analysis. The account is extended so as to include other tracheated systems, partly by means of a number of calculated examples (Tables 2-7), partly from observations on
(2) During flight the primary tracheae supplying the wing muscles in dragonflies and other large insects must be strongly ventilated, but in small insects like Drosophila diffusion of respiratory gases is sufficient to account for the entire transport between the spiracles and the end of the tracheoles even at the highest rates of metabolism. (3) Beyond the primary tracheae and air sacs, air-tube diffusion is sufficient in all insects studied except in the giant belostomid bugs where the secondary tracheae are also ventilated. It is shown that muscular pumping plays an insignificant role in the transport of respiratory gases.
(4) The efficiency of the tracheal system is due to the large hole fraction charac-teristic of wing muscle. It varies from io~x to io~2 as far as the secondary supply is concerned, and from icr2 to io"3 in the tertiary and tracheolar systems. The rate of diffusion of O2 is then io3 to io6 times greater than in the liquid phase and for CO2 it is about 50-5000 times greater.
(5) Diffusion in the liquid phase between the tracheoles and the sites of com-bustion is sufficient in all fibres investigated. It is shown that the main exchange must occur via the tracheoles since, at the high metabolic rates, the diffusion distances are much too long for larger tubes to be effective.
(6) In giant and medium-sized fibres with ' internal' tracheoles indenting the surface the safety factor is of the order of 2-3, but in fibres supplied only from the surface the oxygen-consuming zone has reached the limiting thickness of about 8-10 fi in Odonata and Blattidae. It is argued that the radial-lamellar fibres in the latter groups are an adaptation which improves oxygen diffusion inside the fibre.
(7) Glucose was used as a model substance for the diffusion of fuel into whole muscle and into the interior of the fibres. Muscular pumping of the blood is essential in almost all active wing muscles but, because of its tidal nature, the concentration of dissolved fuel in the haemolymph must be of the order of 0-5-1 %. It is therefore suggested that the high content of trehalose and of lipid materials in insect blood has evolved as an adaptation to flapping flight.
(8) In many Hymenoptera and in Diptera with giant fibres the difference in fuel concentration between the inside of the fibre membrane and the interior must be about 0-5-1 %. In insects depending on fat for sustained flight (Orthoptera, Lepidoptera, Coleoptera), the fibres are smaller in diameter and appear to be loosely packed so that relatively small amounts of free fatty acids in the sarcoplasma should be sufficient (0-001-0-02 %) while muscular pumping and protein-bound lipid serve to transport fuel into the muscle.
SUMMARY
1. The tracheal system of insect wing muscle is so dense that between 1 cr1 and 1 o"3 of any cut area is occupied by air tubes. In most cases, air tube diffusion of O2 and C08 through the muscle is therefore several thousand times quicker than diffusion in the liquid phase.
2. In large insects the primary tracheal supply must be strongly ventilated while diffusion is sufficient in the remaining part of the air tubes, even at the highest metabolic rates encountered in any insect.
3. The tracheoles represent the main site of exchange between the gaseous and the liquid phase while the tracheae are of little significance in this respect. The fibres cannot exceed about 20/1 in diameter unless the tracheoles indent the surface and become 'internal'.
4. Muscular pumping of air and blood due to shortening is of little importance for the exchange of gases but of major importance for the supply with fuel for combustion. However, the large fibre diameters and the tidal nature of the pumping necessitates a very high concentration of fuel in the haemolymph. The high concentration of trehalose in insect blood is considered to be an essential adaptation to flapping flight. 5. The transport by diffusion of O2 and COa was followed in detail in a number of concrete examples in the gaseous as well as in the liquid phase. Within a safety factor of 2-3, the rate of transport was always found to be adequate. There is no reason to suggest other mechanisms than a simple, normal diffusion.
I am indebted to Prof .V. B. Wigglesworth for the loan of injection preparations of
Drosophila, to Dr J. Buck for a translation of Nunome'a paper, and to Prof. J. Kofoed,
Dr P. L. Miller, and Dr O. Sten-Knudsen for valuable discussions.
R E F E R E N C E S
AUBER, J. (1961). Observations sur la structure des fibres musculaire geantes (muscle du vol) chez les Dipteres. C.R. Acad. Sci. 353, 3137-29.
BARBER, R. M. (1941). Diffusion in and through Solids. Cambridge University Press. BEUTLER, R. (1937). Uber den Blutzucker der Bienen. Z. vargl. Physiol. 24, 71-115. BUCK, J. (1962). Some physical aspects of insect respiration. Ann. Rev. Ent. 7, 27-56.
CHADWICK, L. E. (1953). In Insest Physiology. Ed. K. D. Roeder. New York: John Wiley and Sons, Inc.; London: Chapman & Hall, Ltd.
CHADWICK, L. E. & GILMOUR, D. (1940). Respiration during flight in Drosophila repleta Wollaston: the oxygen consumption considered in relation to the wing-rate. Physiol. Zool. 13, 398—410. CLARK, H. W. (1940). The adult musculature of the anisopterous dragonfly thorax (Odonata, Anisoptera).
J. Morph. 67,
523-65-COCKBAIN, A. J. (1961). Fuel utilization and duration of tethered flight in Aphis fabae Scop. J. Exp. Biol. 38, 163-74.
DARWIN, F. W. & PRINGLE, J. S. W. (1959). The physiology of insect fibrillar muscle. I. Anatomy and innervation of the basalar muscle of lamellicorn beetles. Proc. Roy. Soc. B, 151, 194-203.
DAVIS, R. A. & FRAENKEL, G. (1940). The oxygen consumption of flies during flight. J. Exp. Biol. 17, 402-7.
EDWARDS, A., RUSKA, H. & DE HARVEN, E. (1958). The fine structure of insect tracheoblast, tracheae and tracheoles. Arch. biol. 69, 351-69.
GEORGE, J. C. & BHAKTHAN, N. M. G. (i960). A study of the fibre diameter and certain enzyme con-centrations in the flight muscles of some butterflies. J. Exp. Biol. 37, 308-15.
HEMMINGSEN, E. & SCHOLANDER, P. F. (i960). Specific transport of oxygen through haemoglobin solutions. Science, 133, 1379-81.
HOCKING, B. (1953). The intrinsic range and speed of flight in insects. Trans. R. Ent. Soc. Land. 104, 223-345.
HOWDEN, G. F. & KILBY, B. A. (1961). Biochemical studies on insect haemolymph. II. The nature of the reducing material present. J. Ins. Physiol. 6, 85-95.
JOST, W. (1952). Diffusion in Solids, Liquids, Gases. New York: Academic Press Inc.
KROGH, A. (1919). The rate of diffusion of gases through animal tissues, with some remarks on the coefficient of invasion. J. Physiol. 53, 391-408.
KROGH, A. (1920a). Studien ttber Tracheenrespiration. II. Ober Gasdiffusion in den Tracheen. PflOg. Arch. ges. Physiol. 179, 95-112.
KROGH, A. (19206). Studien Uber Tracheenrespiration. III. Die Kombination von mechanischer Ventilation mit Gasdiffusion nach Versuchen an Dytiscuslarven. Pflug. Arch. ges. Physiol. 179,113-20. KROGH, A. (1929). The Anatomy and Physiology of Capillaries. New Haven: Yale University Press. KROGH, A. & WEIS-FOGH, T. (1951)- The respiratory exchange of the desert locust. (Schistocerca
gregaria) before, during and after flight. J. Exp. Biol. 38, 344-57.
LOCKE, M. (1958). The co-ordination of growth in the tracheal system of insects. Quart. J. Micr. Sci. 99.
373-91-MILLER, P. L. (1962). Spiracle control in adult dragonflies (Odorxata). J. Exp. Biol. 39, 513-35. MOLLKR, H. (1921). Uber Lethoceris Uhleri Mont. Zool. Jb. 4a, 43-90.
NEVILLE, A. C. (1960). Aspects of flight mechanics in anisopterous dragonflies. J. Exp. Biol. 37, 631-S6.
NUNOME, J. (1944). Studies on the respiration of the silkworm. Part I. Diffusion of oxygen in the respiratory system of the silkworm. Bull. Seric. Exp. Sta. Chosen, 13, (1), 17-39. (Translated from the Japanese for Dr J. Buck.)
NUNOME, J. (1951). Studies on the respiration of the silkworm. Part III. On the air current of respira-tion. J. Seric. Sci. Japan, 20 (2), 111-27 (Translated from the Japanese for Dr J. Buck.)
POLACEK, I. & KUBISTA, V. (i960). Metabolism of the cockroach Periplaneta americana during flight. Physiol. Bohemoslov. 9, 228-34.
PRINGLE, J. W. S. (1957). Insect Flight. Cambridge University Press.
SCHOLANDER, P. F. (i960). Oxygen transport through haemoglobin solutions. Science, 131, 585-90. SMITH, D. S. (1961a). The structure of insect fibrillar muscle. J. Biophys. Biochem. Cyt. 10, No. 4
(suppl.), 123-58.
SMITH, D. S. (19616). The organization of the flight muscle in dragonfly, Aeshna sp. (Odonata). J. Biophys. Biochem. Cyt. 11, 119-44.
THORPE, W. H. & CRISP, D. J. (1947). Studies on plastron respiration. II. The respiratory efficiency of the plastron in Aphelocheirus. J. Exp. Biol. 24, 270-303.
TIEGS, O. W. (1955). The flight muscles of insects—their anatomy and histology; with some obser-vations on the structure of striated muscle in general. Phil. Trans. B, 338, 221-347.
TiETZ, A. (1962). Fat transport in the locust. J. Lipid. Res. 3, 421-26.
VOGELL, W., BISHAI, F. R., BOCHER, T., KuNGENBERG, M., PETTE, D. & ZEBE, E. (1959). Ober struk-turelle und enzymatische Muster in Muskeln von Locusta imgratoria. Biochem. Z. 333, 81-117. WEIS-FOGH, T. (1952). Fat combustion and metabolic rate of frying desert locusts (Schistocerca gregaria
Forsk&l.) Phil. Trans. B, 337, 1-36.
WEIS-FOGH, T. (1956). The ventilatory mechanism during flight of insects in relation to the call for oxygen. Proc. XlVth Int. Congr. Zool. Copenhagen, 1953, 283-85.
WEIS-FOGH, T. (1961). Power in flapping flight. In The Cell and the Organism. Ed. Ramsay and Wigglesworth. Cambridge University Presa.
WEIS-FOGH, T. (1964a). Functional design of the tracheal system of flying insects as compared with the avian lung. J. Exp. Biol. 41, 207-27.
WEIS-FOGH, T. (19646). Ventilatory mechanisms in flying insects (in preparation).
WIGGLESWORTH, V. B. (1950). A new method for injecting the tracheae and tracheoles of insects. Quart. J. Micr. Sci. 91, 217-24.
WIGGLESWORTH, V. B. (1963). A further function of the air sacs in some insects. Nature, Land., 198, 106. ZEBB, E. (1954). Ober den Stoffwechsel der Lepidopteren. Z. vergl. Physiol. 36, 290-317.
ZEBE, E. (1958). Untersuchungen zum Fettotofrwechsel der Insekten. Verh. dtsch. Zool. GeseU. 1958,
I3I-37-APPENDIX
Steady-state diffusion in tissue with consumption
dis-tance. The procedure is illustrated in the case of equation (3) and is similar to the treatment offered for instance by Erlang (in Krogh, 1929) and by Crisp (Thorpe & Crisp, 1947; Appendix). Many other examples may easily be developed. When dealing with respiratory gases, the equations are solved with respect to the difference in partial pressure Ap necessary to account for the flow. The same formulae apply to the diffusion of solutes like glucose if the concentration difference Ac and the diffusion coefficient D are inserted instead of Ap and the permeability constant P respectively.
Tissue diffusion means that the transport is assumed to take place in an isotropic liquid
(blood and cytoplasm) while air-tube diffusion means that the transport is confined to the system of air tubes and that transport in the liquid phase is insignificant, as dis-cussed on p. 232.
[image:25.451.51.396.213.440.2]Tinue
Fig. i a. Examples on diffusion systems considered in the Appendix. A-D refer to linear one-dimensional cases and E-J to radial one-dimensional cases. A heavy arrow marked ' tissue' shows the direction of flow in an isotropic tissue (tissue diffusion) and the arrows marked 'air' show the direction when the transport occurs in the tracheal system (air-tube diffusion).
(a) Rectilinear flow
Tissue diffusion. Consider a rectangular block of homogeneous tissue, L cm. long
and with end area Ao from which diffusion takes place in the direction from x = o to
x = L (Fig. 12A). For any value of x, Ax = Ao and we know that all oxygen
con-sumed beyond Ax must pass this surface by diffusion due to the pressure gradient
dpjdx. We then have for the net flow Jx through Ax,
J
x= A
x{L-x)m = A
0{L-x) = -PA
0(dp/dx),
from which —dp = mjP{L — x) dx.
Let/) = p0 for x = o and/) = p^ for x = L. Integration between these limits gives
Air-tube diffusion. Consider a similar rectangular block of tissue pierced in the
direction of x by parallel air tubes whose summed areas <x0 for x = o amount to
ao = aA0, where a is the hole fraction (Fig. 12, B and C). Provided that these tubes are
' tapped' at a rate corresponding to the consumption in the tissue they pass either by diffusion in the liquid phase, as in the terminal tracheoles, or by diffusion in regularly spaced side tubes (see Figs. 4, 5), the transport can be calculated when a = a(x) is known.
Case A. The summed cross-sectional area of the tubes is constant and independent
of x, i.e. ax = aAg, as in the main branches of Cossus larvae (Krogh, 1920),
Aphelo-cheirus (Thorpe & Crisp, 1947) and Rhodnius (Locke, 1958). According to Fig. 12B
and the previous example, we then get
mLi
(4)
Equation (4) also applies to branching systems provided that the air tubes run approxi-mately parallel to the #-axis.
Case B. The tubes taper so that a, i.e. the area, decreases linearly with x (cf.
part of Fig. 7). We have, ax = aA^i—xfL), giving
rf (5)
Case C. The tubes taper so that their diameters decrease linearly with x (cf. Figs. 6,
12C), a n d ax = aA0(i—xjL)2. For 0 ^ x < L,
. ml?, L
...Ap = —-hxr Y . (6)
aP L — x v '
Case D. Diffusion in the central trachea. Consider a cylindrical muscle of length L
and radius R supplied from an axial primary trachea of radius r0 from which secondary tracheae branch off (Fig. 12D). When r0 ^ R/10, the metabolic rate of the muscle is approximately mnR2L. The problem is to what extent diffusion in the primary
trachea can account for transport in the axial direction x. The flow across a cross section of the tube is Jx = mnR\L—x) = —irr^P (dp/dx), which when integrated
from x = o and/) = p0 to x = L and/) = pL and solved with respect to L gives
L-ro/i?(2A/)P/m)i (7)
(b) Radial flow
Only cylindrical cases are treated, the radius of the tissue cylinder being R, its length
L and the distance from the axis r.
Tissue diffusion. The diffusion takes place either from a co-axial cylindrical tube of
radius r0 towards the periphery, or from the peripheral surface towards the cylinder axis.
Case A. Diffusion from inner cylindrical tube towards the periphery. At distance
r, the flow through the tissue surface Ar is (cf. Fig. 12 E),
Integrated from r = roandp = p0tor = Randp = pR, we get
(8)
Case B. When the diffusion occurs from the surface of the cylinder corresponding
to r = R towards an inner cylinder surface of radius r0 (cf. Fig. 12 F), the solution is
^ (9a)
For diffusion to the centre axis where r0 ->• o, we have
A
^^p- (9b)
Air-tube diffusion. The first case is directly related to the preceding one.
Case A. Consider a cylindrical piece of tissue supplied from the outside by tubes
radiating towards the axis and where the summed cross-sectional area of the tubes a is proportional to the distance from the axis (Fig. 12G). We have then aB = aznRL.
If r0 -> o, the solution is
Case B. The tissue cylinder, is supplied from a central co-axial tube of radius r0 which is 'tapped' by means of radially arranged secondary tubes whose summed area at the origin is aTo = azTrrJL. Let us first assume ar = aro = constant (Fig. 12H). Jr = nmL(R2-r2) = -2nr0LdP{dpjdr). Integrated from r = r0 and p = p0 to
r = R and p = pR, we get
L e t / = rojR be inserted in equation (11) and solved for/,
Case C. As above, but with linearly decreasing area (Fig. 121), so that
ar =
. m
For any value of r, we have
Af = jjfeR2^—ro) + i?(r2 — rl)). (^^O
If, again, / = ro/R, we have
D. As the two preceding examples, but with linearly increasing area so that
aT = a27rr0Lr. m I R
Case E. A tissue cylinder is supplied from thin tubes radiating out from the central
axis (Fig. 12 J; cf. outer half cylinder of the lobe in Fig. 5). The hole fraction is here defined as a = ct^lzRL. When ar = a^ = constant, we have
nmR2
Case F. Same as (E) but with linearly decreasing area, ar = 2aL(R — r). The