Copyright 0 1999 by the Genetics Society of America
Direct Cloning of Yeast Genes from an Ordered Set
of Lambda Clones in
Saccharomyces cerevisiae by Recombination
in
Vivo
James
R. Erickson and Mark Johnston
Department of Genetics, Washington University School of Medicine, St. Louis, Missouri 6 3 1 1 0 Manuscript received March 9, 1992
Accepted for publication July 29, 1992
ABSTRACT
We describe a technique that facilitates the isolation of yeast genes that are difficult to clone. This technique utilizes a plasmid vector that rescues lambda clones as yeast centromere plasmids. The source of these lambda clones is a set of clones whose location in the yeast genome has been determined by L. Riles et al. in 1993. The Esherichia coli-yeast shuttle plasmid carries URA3, ARS4
and C E N 6 , and contains DNA fragments from the lambda vector that flank the cloned yeast insert. When yeast is cotransformed with linearized plasmid and lambda clone DNA, Ura+ transformants are obtained by a recombination event between the lambda clone and the plasmid vector that generates an autonomously replicating plasmid containing the cloned yeast DNA sequences. Genes whose genetic map positions are known can easily be identified and recovered in this plasmid by testing only those lambda clones that map to the relevant region of the yeast genome for their ability to complement
the mutant phenotype. This technique facilitates the isolation of yeast genes that resist cloning either because (1) they are underrepresented in yeast genomic libraries amplified in E . coli, (2) they provide phenotypes that are too marginal to allow selection of the gene by genetic complementation or (3)
they provide phenotypes that are laborious to score. We demonstrate the utility of this technique by isolating three genes, GAL83, SSNZ and M A K 7 , each of which presents one of these problems for cloning.
I
N
yeast, cloning a gene by complementation of a recessive mutation is usually a relatively simple matter. However, a number of factors can limit t h e ability to clone a particular gene. First, libraries am-plified in Esherichia coli can be depleted of some genes. For example, RAD4 is lethal to bacteria (SIEDE a n d ECKARDT-SCHUPP 1986); K A R 2 confers a significant growth disadvantage upon the host (ROSE, MISRA a n d VOGEL 1989). Such problems are exacerbated with successive amplifications of a library. In addition, since many circulating libraries were constructed using par- tially digested, size selected yeast genomic
D N A ,
t h e availability of restriction sites greatly influences the probability of cloning any stretch ofDNA.
A
second factor that can contribute to difficulty in cloning a gene is the phenotype caused by the muta- tion used in screening of the library.As
yeast genetics becomes increasingly sophisticated, the phenotypes ofmutations can be increasingly subtle. In some cases the phenotype of the mutant is too subtle to allow rare complementing clones to be identified. In other cases, the phenotypes of the mutant can only be scored after laborious manipulations, which severely reduces the number of transformants that can be screened for
the complementing gene.
All
these factors have made some yeast genes difficult t o isolate by conventional methods.In this paper, we describe how we have solved these
Genetics 1 3 4 151-157 (May, 1993)
problems by adapting the method of MA et al. (1987)
for the purpose of cloning yeast genes. The method is based on the observation that a double-strand break in a plasmid can be repaired by cotransforming with a linear fragment containing
D N A
sequences that flank the double strand break (ORR-WEAVER, SZOS-TAK a n d ROTHSTEIN 1 9 8 1 ; KUNES, BOTSTEIN a n d
Fox
1985). YeastD N A
sequences are rescued fromlambda clones by cotransforming yeast with the lambda clone and linearized plasmid that contains
phage lambda sequences that flank the cloned yeast
DNA.
T h e cloned yeast sequences are thus trans- ferred to a yeast centromere plasmid; the resulting colonies can then be screened for complementation.This method takes advantage of the restriction map of the yeast genome compiled by RILES et al. (1 993), which provides lambda clones that cover 98% of the yeast genome. If the genetic map position of a partic- ular gene is known, the lambda phage carrying it can be identified by testing for complementation of a mutation of only those lambda clones that span t h e region or, in the worst case, the entire chromosome.
We have successfully applied this method to clone three genes: GAL83 (MATSUMOTO, TOH-E a n d OSH-
IMA 1 9 8 1 ; MATSUMOTO, YOSHIMATSU a n d OSHIMA 1983), a gene apparently not present in any of four genomic plasmid libraries screened; SSN2 (CARLSON
152 J. R. Erickson and M. Johnston
be complemented easily in a plate assay; and M A K 7
(WICKNER and LIEBOWITZ 1976), a gene whose phe- notypic effects are laboriously scored.
MATERIALS AND METHODS
Strains and media: E. coli strain DH5a (Bethesda Re- search Laboratories) was the host for all plasmids. Yeast strains used in complementation tests for identifying the various genes were: YM3033 (MATa, ura3-52, hid-200, ade2-101, lys2-801, ga180A538 gal83-2000, LEU2::GALl- lacZ); MCY447 (MATa, ura3-52, ade2-101, his4-539, snfl-
28, ssn2-4) obtained from M. CARLSON; 2778 (MATcu, mak7-1, ura3, met5 L-A-HN, M-a, rho") obtained from R.
WICKNER. Yeast strain 1074 (MATa, leul, karl-1, L-A-HN, M , ) was used to cytoduce killer RNA, and 5x47 (MATa/ MATa, hisl/+, trpl/+, u r d / + M-a) is the killer-sensitive
tester strain.
LB medium containing 100 pg/ml ampicillin was used to grow all bacterial strains harboring plasmids. Standard SD medium and supplements for yeast are described in ROSE, WINSTON and HIETER (1990). Amino acids and nucleotide base supplements were omitted where appropriate. 2-deoxy- D-galactose (2dGal) was added to 0.25%; 4.7 MB plates used to score the killer phenotype were described by WICKNER and LIEBOWITZ (1 976). Plates containing 5-flouro-orotic acid were described by BOEKE, LACROUTE and FINK (1984).
Plasmid construction: T h e CEN-ARS plasmid pBM2240
used to recover lambda clone inserts is based on pRS316 (SIKORSKI and HIETER 1989).?'he lambda DNA fragment flanking the yeast DNA insert on the right was an 858-bp BamHI-Xbal fragment generated by PCR using as template the lambda cloning vector MG3 (derived from the lambda cloning vector 147.1 (LOERAN and BRAMMER 1980)). The PCR was primed with the following oligonucleotides: 5'-
GCTCTAGAGATCTGCCAGATTACGCTCCTG-3' and
T h e PCR product was digested with BamHl and XbaI, gel purified and subcloned into the BamHI-XbaI site of pRS3 16. T h e lambda DNA fragment flanking the yeast DNA insert on the left was a 1.5-kb KpnI fragment from MG3 extending from nucleotide 17053-18556 (DANIELS et al. 1983). This fragment was subcloned in both orientations into the plas- mid containing the BamHI-XbaI fragment. T h e orientations were verified by DNA sequencing. T h e plasmid containing the two lambda fragments i n the same relative orientation as they occur in lambda was designated pBM2240. T h e plasmid with the KpnI fragment in the opposite orientation was designated pBM2241. Plasmid pBM2384, a high-copy plasmid used to recover lambda clone inserts, was con- structed by subcloning the Kpnl fragment and the BamHI- XbaI fragment of lambda from pBM2240 into the 2-pm- based vector pRS426.
DNA manipulations: Lambda clone DNA was prepared
from a 5-ml liquid lysate using Lambdasorb (Promega), according to the manufacturers suggested protocol for liq- uid lysates (Promega technical bulletin 32). Lysates were prepared by pulling an agar plug with the large bore end of
a Pasteur pipet and eluting in 20 mM Tris.CI, pH 8
+
20mM MgCI, at 4" overnight. 100 pl of the phage eluate were combined with 100 pl 10 mM CaClp
+
10 mM MgC1.L and 100 pI of a saturated culture of E . coli strain C600 and incubated for 30 min at 37". Five ml of prewarmed NZC broth (per liter: 10 g NZ amine, 5 g NaCI, 5 g yeast extract, 1 g casamino acids, 2 g M g S 0 4 . 7 H 2 0 ) was added and incubated with vigorous aeration until the culture lysed (8- 10 hr). Cell debris was spun down and the supernatant was5'-CGGGATCCCTGGAAGAAATACTCATAAGCC-3'.
treated with 0.5 PI 10 mg/ml DNase
+
2.5 PI 10 mg/ml RNase for 30 min at 37" prior to adding adsorbent. Recom- binant plasmids were recovered in E. coli from yeast lysed with glass beads (HOFFMAN and WINSTON 1987).Yeast transformation and phenotype scoring: Yeast
transformations were performed by the spheroplast method (BURGERS and PERCIVAL 1987). In a typical experiment to rescue lambda clones, 100-200 ng of pBM2240 digested with EcoRI and XhoI and 1-2 pg of lambda clone DNA were used to transform yeast to Ura+. After digestion with EcoRI and XhoI, the vector DNA was phenol extracted, ethanol precipitated and resuspended in T E pH 8 at 100 ng/pl prior to transformation. Transformants were scored for the pres- ence of the HIS3 or GAL83 genes by replica plating to SD- Ura-His or SD-Ura+ 0.25% SdGal, respectively. T h e clumpy growth of ssn2 mutants was scored after growth of transfornlants in liquid SD-Ura media. T h e phenotype of mak7 mutants was scored as described by WICKNER and
LEIBOWITZ (1976). To score for mak7 complementing activ- ity, Ura+ transformants from each transformation plate were picked to SD-Ura. These isolates were grown overnight at 30" and mated with the killer and mitochondrial donor, 1074, to cytoduce killer plasmid. After 6 hr at 30" the mating mixture was streaked for single colonies onto SD- Ura-Leu to select for the inheritanced of the plasmid (Ura') and against the nucleus from the killer+ strain (Leu+). Suc- cessful cytoduction was scored by the ability to grow on YP
+
glycerol. Rho+ haploids were then printed to a lawn of the killer sensitive strain 5 x 4 7 . Halos indicating mainte- nance of the killer plasmid on 4.7 MB plates were scored after 2 days at 20". Five independent transformants were used to score the phenotype.Enzyme assays: Beta-galactosidase (lacz) expression from the GALl-lad fusion, PRY 18 1 integrated at the LEU2 locus was assayed as described previously (YOCUM et al. 1984; FLICK and JOHNSTON 1990). Yeast were grown in SD-Ura medium containing either 2% glucose or 2% galactose. Enzyme activity for the recombinant p5083 is the average of four independent recombinants. Invertase was assayed as described previously (GOLDSTEIN and LAMPEN 1975; NEI- GEBORN and CARLSON 1984). Repressed cultures were grown to mid-log phase in 2% glucose, and cells were derepressed for 3 hr in SD-Ura+ 0.05% glucose. T h e results reported are the average of two assays.
RESULTS
Recovery of the HIS3 gene using pBM2240: To
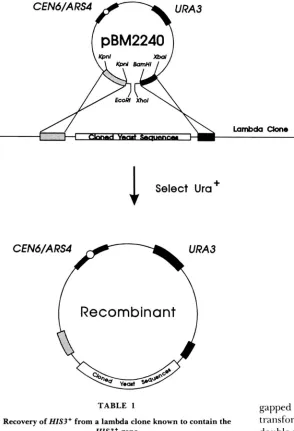
establish that the yeast DNA in lambda clones can be recovered in a yeast plasmid, a lambda phage that was previously shown to contain the HZS3+ gene was used. A his3 mutant (YM3033) was cotransformed with DNA from the HZS3+-containing lambda clone (5380) and either with either of t w o different plasmid vectors containing homology to the arms of the lambda clon- ing vector flanking the inserted yeast DNA sequences. Plasmid pBM2240 (Figure 1) contains the lambda arm DNA fragments in the same relative orientation as they exist in the lambda clone; pBM2241 contains the lambda left arm fragment in the opposite orientation. Only pBM2240 is predicted to yield a closed circular plasmid through a double recombination event be- tween the vector and the lambda clone.
Cloning by Recombination in Yeast 153
FIGURE I.-Diagram of the recombination be-
a u ~
tween pBM2240 and a lambda clone, and the recom- binant product. pBM2240 is based on the E. coli- yeast shuttle vector pRS3 16 (SIKORSKI and HIETER 1989). The shaded regions represent fragments of the lambda-cloning vector that were subcloned into pRS316 using the indicated restriction sites. The filled regions represent sequences required to select and maintain the plasmid in yeast. pBM2240 was digested with EcoRl and XhoI prior to cotransfor- mation with lambda clone DNA. High-efficiency transformation is the result of recombination be- tween the plasmid and the lambda clone. An analo- gous 2-pm-based high copy plasmid was also con- structed (pBM2384, see MATERIALS AND METHODS).[
Recombinant
)
TABLE 1
Recovery of HZS3+ from a lambda clone known to contain the
HZS3+ gene
gapped pBM2241 did not increase the frequency of transformation. These are the expected results if the double strand break in the vector is being repaired by recombination with the lambda clone, as diagramed in Figure 1. As expected, most of the Ura+ transform- ants are His+. Loss of the recombinant plasmid by selection on 5-FOA (BOEKE, LACROUTE and FINK 1984) resulted in the concomitant loss of the Ura+ and His+ phenotypes. This is in contrast to the two
His+ isolates obtained with pBM2241, in which the two phenotypes did not cosegregate. T h e identity of four recombinant plasmids was confirmed by restric- tion enzyme digestion (data not shown).
Recovery of GAL83: GAL83 is a gene required for glucose repression of GAL gene expression (MATSU-
MOTO, YOSHIMATSU and OSHIMA 1983; FLICK and JOHNSTON 1990). We believe that this gene is under- represented or absent in yeast plasmid libraries am- plified in E. coli because we have been unsuccessful in our repeated attempts to isolate it from several differ- ent libraries by genetic complementation. Because the map position of GAL83 is known (MATSUMOTO, YOSH-
Transforming DNA
Plasmid Lambda transformants Ura' rested
No. Ura' No. His' clones/no.
pBM2240 uncut None 2000 0154
pBM2240 cut None 45 0134
pBM2240 cut 5380 (100 ng) 200 20122 pBM2240 cut 5380 (2 pg) 1000 44/49
pBM2241 uncut None 1300 0152
pBM2241 cut None 85 0150
pBM2241 cut 5380 (2 p g ) 115 2/55
154 J. R. Erickson and M . Johnston
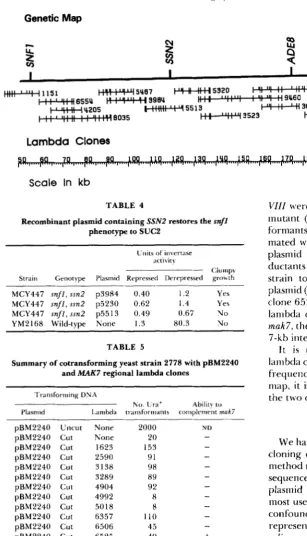
Genetic Map
h
Y
z
I I
IF
6977 5083
H+tHtHi56OU -7150 ki4Hi 5313 k"Wi U678
M 1581
65UO 6224
5961
6793 P P W I k l 6 5 9 2
6697
Lambda
Clones PI-w" 4019FIGURE 2.-Map of chromosome V sur- rounding GAL83. T h e location of CEN5 and H I S 1 were determined by hybridization (RILES et al. 1993). T h e location of GAL83
was deduced by determining which lambda clone complemented the ga183-2000 muta- tion (see MATERIALS AND METHODS). T h e lambda clones used spanned the interval from
6540-6793. T h e physical scale reflects the position of this interval in relation to the left end of Chromosome V. T h e physical scale ( i n kb) begins at the left telomere.
Scale
In
kbTABLE 2
Cotransformation of YM3033 with pBM2240 and GAL83 regional lambda clones
Tral>sforming DNA
No. Ura' No. 2dGal' cIones/no. Plasrnid Lambda transforniants Urn+ tested
pBM2240 uncut None 2000 0/54
pBM2240 cut None 45 Of34
pBM2240 cut 4049 400 Of54
pBM2240 cut 5083 550 49 /56
pBM2240 cut 531 3 400 0 /54
pBM2240 cut 6224 400 0/52
pBM2240 cut 6540 400 O f 5 1
pBM2240 cut 6793 250 Of54
pBM2240 cut 6977 500 O f 5 1
pBM2240 cut 7 150 150 0 /24
TABLE 3
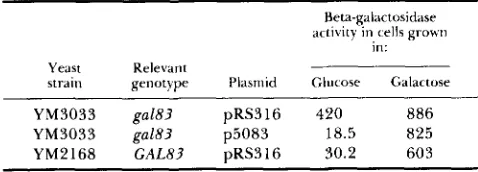
Recombinant plasmid containing sequences from the lambda clone 5083 restores wild-type regulation to a CALI-lac2 fusion
Beta-galactosidase activity in cells grown
in:
straln
Yeast
genotype Plasmid Glucose Galactose RdeVdnt
YM3033 gal83 pRS316 420 886
YM3033 gal83 p5083 18.5 825
YM2168 GAL83 pRS316 30.2 603
IMATSU and OSHIMA 1983), it is amenable to cloning in pBM2240 by cotransforming with lambda clones carrying the relevant region from the yeast genome.
We employed the galactose analog 2-deoxy-D-galac- tose (2dGal) to test transformants for the presence of
GAL83. This compound is converted to a form that is toxic to yeast (2-deoxygalactose-1 -phosphate) by the action of galactokinase, the product of the GALl gene (PLATT 1984). Wild-type yeast grown in the presence of glucose are resistant to 2dCal due to glucose repres- sion of GALl expression; gal83 mutants express GALl
under these conditions and are therefore sensitive to the analog.
To clone GAL83, eight lambda clones that encom-
pass the region to which GAL83 maps (between C E N 5
and H I S l , see Figure 2) were cotransformed with pBM2240 cut with EcoRI and XhoI. As expected, the inclusion of lambda DNA resulted in an increase in transformation efficiency (Table 2). T o screen for ga/8?-complementing activity, Ura+ transformants
were tested for sensitivity to 2dGal, which revealed that a single lambda clone, 5083, carries GAL83. This was verified by assays of GALl expression in the transformants that contain GAL83 (Table 3). These results delimit the location of GAL83 to the approxi- mately 6 kb of DNA unique to this clone.
Cloning of SSN2: As a second demonstration of the utility of the lambda clone rescue technique, we em- ployed it to isolate a gene whose mutant phenotype is too marginal to allow it to be isolated readily from a complex library. Such a gene is S S N 2 , whose muta- tions suppress the inability of s n f l mutants to grow on raffinose as a sole carbon source. Mutants of s n f l are unable to grow on raffinose because they are unable to derepress expression of invertase (encoded by
S U C 2 ) , the enzyme responsible for cleaving raffinose;
ssn2 mutants partially restore SUC2 expression in snfl mutants. Because mutations in ssn2 only modestly suppress the s n f l defect, they do not confer a readily selectable phenotype. T h e most easily scored pheno- type of ssn2 mutants is their "clumpy" growth in liquid culture. DNA was prepared from individual lambda clones containing the region of chromosome IV to which S S N 2 maps and cotransformed with pBM2240 cut with EcoRI and XhoI (data not shown). Ura+ trans- formants were tested for clumpy growth and the ability to grow on YP Raffinose. A single lambda clone, 55 13, complemented the ssn2 mutant pheno- types (see Figure 3). This was confirmed by assaying
SUC2 expression in the transformants (Table 4). Cloning of MAK7: A third example of a gene that would be difficult to clone by screening libraries is
Cloning by Recombination i n Yeast 1.55
Genetic Map
h
Ir
5
oc
2
x
Lambda
ClonesFIGURE 3.-Map of chromosome IV near S N F 1 . S S N 2 was shown to be tightly linked to S N F l (CARLSON et al. 1984). ‘The location of S N F l and
ADEB were determined by hybridiza- tion (RILES et al. 1993). The seven lanlbda clones between S N F l and
ADER were used with pBM2240 (clones numbered 8000 and higher are costnid clones). A single lambda clone, 551 3 , complemented the ssn2
phenotype. T h e physical scale ( i n kb) begins a t the left telomere.
Scale in kb
TABLE 4
Recombinant plasmid containing SSN2 restores the snfl
phenotype to SUC2
Units of invertase activity
Clun1py Strain Genotype Plasmid Repressed Ikrepressed growth
MCY447 snfl, ssn2 p3984 0.40 1.2 Yes MCY447 s n f l , ssn2 p5230 0.62 1.4 Yes MCY447 snfl, ssn2 p5513 0.49 0.67 No YM2168 Wild-type None 1.3 80.3 N o
TABLE 5
Summary of cotransforming yeast strain 2778 with pBM2240 and M A K 7 regional lambda clones
Transforming D N A
N o . Ura’ Ability t u
Plasnlld Lanlbda tr-msforl11ants complement mak7
pBM2240 Uncut None 2000 ND
pBM2240 Cut None 20
pBM2240 Cut 1623 153 pBM2240 Cut 2590 91
pBM2240 Cut 3138 98 pBM2240 Cut 3289 89
pBM2240 Cut 4904 92
pBM2240 Cut 4992 8
pBM2240 Cut 5018 8
pBM2240 Cut 6357 110
pBM2240 Cut 6506 45
pBM2240 Cut 6525 49
- - - - - -
-
-
-
-
+
for replication of the double-stranded kilfer RNA plasmid (WICKNER and LIEBOWITZ 1976). Since the killer plasmid is not maintained in a mak mutant, potential transformant clones must have the killer plasmid reintroduced by cytoduction to score for com- plementing activity. This severely limits the number of transformants that can be tested for the presence of the gene.
M A K 7 had been previously mapped near the end of the left arm of chromosome VZZZ (WICKNER 1979). T h e 1 0 left-most lambda clones from chromosome
VZZZ were cotransforrned with pBM2240 into a mak7
mutant (see M A ~ r E R I A L s A N D METHODS). Ura+ trans-
formants produced from each lambda clone were
mated with a Karl nlutant to cytoduce the MI RNA plasmid into the transformants. T h e resultant cyto- ductants were printed to a lawn of a killer-sensitive strain to score their ability to maintain the killer plasmid (Makt phenotype). As seen in Table 5, lambda clone 6525 complemented the mak7 defect. Since the lambda clones 2590 and 6505 did not complement
mak7, the gene has been localized to an approximately 7-kb interval (see Figure 4).
I t is noteworthy that cotransforming with two
lambda clones (4992 and 50 18) actually decreased the frequency of transformation. Based on the genetic map, it is likely that the overlapping region between the t w o clones contains CEN8.
DISCUSSION
156 J. R. Erickson and M. Johnston
Genetic Map
B
9
h
h rn
s
g
cr,
Lambda Clones
Scale
In
k bFIGURE 4.-Map of chromosome VIIInear the left telomere. T h e location of S P O I l was determined by hybridiza- tion; the subtelomeric repeats X and Y hybridize to lambda clone 4904 (RILES et al. 1993). To clone M A K 7 , the 10 left- most lambda clones were used with pBM2240 to cotransform a mak7 mu- tant. A single lambda clone, 6525. com- plemented the mak7 defect. T h e posi- tion of CENR is based on transformation frequencies (see RESULTS). T h e physical scale (in kb) begins at the left telomere.
screened. This allows one to reduce greatly the com- plexity of the library being screened because each lambda clone is tested individually for complementing activity. I t would not be unreasonable to attempt to clone a gene that is only mapped to a chromosome by screening all of the lambda clones that span that chromosome for complementation by cotransforma- tion. Such an approach is attractive because of the ease of mapping genes to chromosomes (WAKAM and SHERMAN 1990). For genes that lie on larger chro- mosomes, such as chromosome IV, which is repre- sented by 114 lambda clones, it would be desirable to gather more genetic map information so that fewer lambda clones are needed. Once a gene has been localized to a lambda clone or overlapping clones, the actual location of the gene is often delimited to a fairly small interval, which simplifies subcloning the gene of interest.
This approach can also be used to convert lambda clones into high-copy yeast plasmids. We have con- structed a 2-pm-based high copy plasmid similar to pBM2240. This plasmid (pBM2384, see MATERIALS
AND METHODS), when cut with EcoRI and Xhol and cotransformed with the GAL83-containing lambda clone 5083, yielded GAL83+ transformants at high frequency, similar to the results reported in Table 2.
There are three limitations that may prevent a particular gene from being cloned by this method. First, a few regions of the yeast genome are currently inaccessible to the technique because of gaps in the map for which no clones exist. Second, not all of the restriction map is based on lambda clones; in many cases, small gaps in the lambda clone map were spanned by cosmid clones. Such intervals cannot be recovered in pBM2240. We are in the process of developing a similar vector designed to rescue yeast sequences cloned in cosmid vectors. Third, it is im- portant to note that two different lambda vectors, MG3 and MG14, were used in constructing the library
used to generate the yeast restriction map. Most of the lambda clones are based on the MG3 vector, whose inserts pBM2240 was designed to recover. However, if one needed to recover a lambda clone from the MG14 vector, a new plasmid analogous to pBM2240 would have to be constructed. Neverthe- less, since most regions of the yeast genome can be recovered by this technique, it provides a useful alter- native to the use of plasmid libraries amplified in E. coli for cloning yeast genes. Finally, it should be noted that pBM2240 is not suited for recovering genes that are tightly CEN-linked because the recombinant plas- mid would be dicentric. T h e 2-pm-based plasmid, pBM2384, would permit the recovery of lambda clones which contain centromeres.
We thank L. RILES for assistance with the lambda clones and map information and M. CARLSON and R. WICKNER for providing yeast strains. This work was funded by a grant from the NIH GM32540. J.R.E. was supported by by NIH institutional training grant T 3 2 GM 07076 to the Division of Biology and Biomedical Sciences.
L I T E R A T U R E C I T E D
BOEKE, J. D., F. LACROUTE and G . R. FINK, 1984 A positive selection for mutants lacking orotidine-5'-phosphate decarbox- ylase activity in yeast: 5-flouro-orotic acid resistance. Mol. Gen. Genet. 197: 345-346.
BURGERS, P. M. J., and K . J. PERCIVAL, 1987 Transformation of yeast spheroplasts without cell fusion. Anal. Biochem. 163:
391-397.
CARLSON, M., B. C. OSMOND, L. NEIGEBORN and D. BOTSTEIN, 1984 A suppressor of snff mutations causes constitutive high- level invertase synthesis in yeast. Genetics 107: 19-32. DANIELS, D., J. SCHROEDER, W. SZYBALSKI, F. SANCER and F.
BLATTNER, 1983 Complete annotated lambda sequence, pp. 519-676 in Lambda fI, edited by R. W. HENDRIX, J. W. ROBERTS, F. W. STAHL and R. A . WE~SBERG. Cold Spring Harbor Press, Cold Spring Harbor, N.Y.
FLICK, J . S., and M . JOHNSTON, 1990 Two systems of glucose repression of the GAL1 promoter in Saccharomyces cerevisiae.
Mol. Cell. Biol. 10: 4757-4769.
Cloning by Recombination in Yeast 157
HOFFMAN, C. S., and F. WINSTON, 1987 A ten-minute DNA preparation from yeast efficiently releases autonomous plas- mids for transformation of Escherichia coli. Gene 57: 276-272.
KUNFS, S., D. BOTSTEIN and M. S. Fox, 1985 Transformation of yeast with linearized plasmid DNA. J. Mol. Biol. 1 8 4 375- 387.
LOERAN, W. A. M., and W. J. BRAMMER, 1980 A bacteriophage lambda vector for cloning large DNA fragments with several restriction enzymes. Gene 20: 249-259.
MA, H., S. KUNFS, P. J. SCHATZ and D. BOTSTEIN, 1987 Plasmid construction by recombination in yeast. Gene 58: 201-216.
MATSUMOTO, K., A. TOH-E and Y. OSHIMA, 1981 Isolation and characterization of dominant mutations resistant to carbon catabolite repression of galactokinase synthesis in Saccharomyces cereuisiae. Mol. Cell. Biol. 1: 19-32.
MATSUMOTO, K., T. YOSHIMATSU and Y. OSHIMA, 1983 Recessive mutations conferring resistance to carbon catabolite repression of galactokinase synthesis in Saccharomyces cerevisiae. J. Bacte-
rial. 153: 1405-1414.
NEIGEBORN, L., and M. CARLSON, 1984 Genes affecting the reg- ulation of SUC2 gene expression by glucose repression in Sac- charomyces cereuisiae. Genetics 108: 845-858.
ORR-WEAVER, T. L., J. W. SZOTACK and R. J. ROTHSTEIN,
1981 Yeast transformation: a model system for the study of recombination. Proc. Natl. Acad. Sci. USA 78: 6354- 6358.
PLATT, T., 1984 Toxicity of 2-deoxygalactose to Saccharomyces cereuisiae cells constitutively synthesizing galactose-metaboliz- ing enzymes. Mol. Cell. Biol. 4: 994-996.
RILES, L., J. E. DUTCHIK, A. BAKTHA, B. K. M c CAULEY, E. C. THAYER, M. P. LECKIE, V. V. BRADEN, J. E. DEPKE and M. V.
OLSON, 1993 Physical maps of the six smallest chromosomes of Saccharomyces cerevisiae at a resolution of 2.6-kilobase pairs. Genetics, 134: 81-0000.
ROSE, M. D., L. M. MIRSA and J. P. VOGEL, 1989 KARZ: a karyogomy gene, is the yeast homolog of the mammalian BiP/ GRP78 gene. Cell 57: 1211-1221.
ROSE, M. D., F. WINSTON and P. HIETER, 1990 Methods in Yeast Genetics: A Laboratory Course Manual. Cold Spring Harbor Press. Cold Spring Harbor, N.Y.
SIEDE, W., and F. ECKARD-SCHUPP, 1986 DNA repair genes of Saccharomyces cerevisiae: complementing rad4 and rev2 muta- tions by plasmids which cannot be propagated in Esherichia coli. Curr. Genet. 11: 205-210.
SIKORSKI, R. S., and P. HIETER, 1989 A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cereuisiae. Genetics 122: 19-27.
WAKAM, L. P. and F. SHERMAN, 1990 Chromosomal assignment of mutations by specific chromosome loss in the yeast Saccha- romyces cereuisiae. Genetics 125: 333-340.
WICKNER, R. B., 1979 Mapping chromosomal genes of Saccharo- myces cereuisiae using an improved genetic method. Genetics 92: 803-821.
WICKNER, R. B. and M. J. LEIBOWITZ, 1976 Chromosomal genes essential for replication of a double-stranded RNA plasmid of Saccharomyces cerevisiae: the killer character of yeast. J. Mol. Biol. 105: 427-443.
YOCUM, R. R., S. HANLEY, R. WEST JR. and M. PTASHNE,
1984 Use of lacZ fusions to delimit regulatory elements of the inducible divergent GALI-GAL10 promoter in Saccharomy- ces cereuisiae. Mol. Cell. Biol. 4: 198551998,