0095-1137/10/$12.00 doi:10.1128/JCM.01050-10
Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Complete Nucleotide Sequence Analysis of Plasmids in Strains of
Staphylococcus aureus
Clone USA300 Reveals a High Level
of Identity among Isolates with Closely Related Core
Genome Sequences
䌤
†
Adam D. Kennedy,
1Stephen F. Porcella,
2Craig Martens,
2Adeline R. Whitney,
1Kevin R. Braughton,
1Liang Chen,
3Carly T. Craig,
1Fred C. Tenover,
4Barry N. Kreiswirth,
3James M. Musser,
5and Frank R. DeLeo
1*
Laboratory of Human Bacterial Pathogenesis1and Genomics Unit, Research Technologies Section,2Rocky Mountain Laboratories,
Hamilton, Montana; Public Health Research Institute, University of Medicine and Dentistry of New Jersey, Newark, New Jersey3;
Cepheid, Sunnyvale, California4; and Center for Molecular and Translational Human Infectious Diseases,
Methodist Hospital Research Institute, and Department of Pathology and Laboratory Medicine,
Methodist Hospital, Houston, Texas5
Received 25 May 2010/Returned for modification 3 August 2010/Accepted 4 October 2010
A community-associated methicillin-resistantStaphylococcus aureus(CA-MRSA) strain known as pulsed-field type USA300 (USA300) is epidemic in the United States. Previous comparative whole-genome sequencing studies demonstrated that there has been recent clonal emergence of a subset of USA300 isolates, which comprise the epidemic clone. Although the core genomes of these isolates are closely related, the level of diversity among USA300 plasmids was not resolved. Inasmuch as these plasmids might contribute to significant gene diversity among otherwise closely related USA300 isolates, we performedde novo sequencing of endogenous plasmids from 10 previously characterized USA300 clinical isolates obtained from different geographic locations in the United States. All isolates tested contained small (2-to 3-kb) and/or large (27- (2-to 30-kb) plasmids. The large plasmids encoded heavy metal and/or antimicro-bial resistance elements, including those that confer resistance to cadmium, bacitracin, macrolides, penicillin, kanamycin, and streptothricin, although all isolates were sensitive to minocycline, doxycycline, trimethoprim-sulfamethoxazole, vancomycin, teicoplanin, and linezolid. One of the USA300 isolates con-tained an archaic plasmid that encoded staphylococcal enterotoxins R, J, and P. Notably, the large plasmids (27 to 28 kb) from 8 USA300 isolates—those that comprise the epidemic USA300 clone—were virtually identical (99% identity) and similar to a large plasmid from strain USA300_TCH1516 (a previously sequenced USA300 strain from Houston, TX). These plasmids are largely divergent from the 37-kb plasmid of FPR3757, the first sequenced USA300 strain. The high level of plasmid sequence identity among the majority of closely related USA300 isolates is consistent with the recent clonal emergence hypothesis for USA300.
Community-associated methicillin-resistant Staphylococcus
aureus(CA-MRSA) infections in the United States are largely
caused by a strain known as pulsed-field gel electrophore-sis type USA300 (USA300) (14, 17, 18). Using comparative whole-genome sequencing, we demonstrated previously that there has been clonal emergence of a subset of USA300 iso-lates, which comprise the epidemic USA300 clone (13). On
average, these USA300 isolates differ by⬃50 single-nucleotide
polymorphisms (SNPs) in the core genome (13). Although there is limited nucleotide diversity in the core genome among these USA300 clinical isolates, comparative DNA sequencing failed to resolve plasmid DNA content, presumably because of significant nucleotide diversity.
Plasmids carry antibiotic resistance genes and virulence de-terminants and spread rapidly within and between species. The contribution of plasmids to the emergence of USA300 and clonal subtypes and their potential spread of antibiotic resis-tance has not been thoroughly investigated, and only a few extrachromosomal elements have been characterized. For ex-ample, strain FPR3757, the first USA300 strain for which there was a complete genome sequence, contains a 4-kb plasmid encoding tetracycline resistance that is 99.9% identical to a
plasmid inStaphylococcus epidermidis(7). Strain FPR3757 also
contains a conjugative 37-kb plasmid (pUSA03) that encodes resistance to mupirocin and high-level resistance to clindamy-cin, a front-line antibiotic used to treat CA-MRSA infections (7). A recent study by McDougal et al. identified pUSA03-like plasmids in USA300 isolates recovered from individuals with invasive disease (16). In comparison, strain USA300_TCH1516 has a 27-kb plasmid (pUSA300HOUMR) that is not related to pUSA03 but is a “mosaic” of plasmids from other staphy-lococcal species (11). These observations are consistent with the findings of Tenover et al. (24), who reported that plas-mids among USA300 isolates varied in size and composi-* Corresponding author. Mailing address: Laboratory of Human
Bacterial Pathogenesis, Rocky Mountain Laboratories, NIAID, NIH, 903 South 4th Street, Hamilton, MT 59840. Phone: (406) 363-9448. Fax: (406) 363-9394. E-mail: [email protected].
† Supplemental material for this article may be found at http://jcm .asm.org/.
䌤Published ahead of print on 13 October 2010.
4504
on May 16, 2020 by guest
http://jcm.asm.org/
tion, although the analysis of nucleotide diversity measured was limited to the presence or absence of four selected antibiotic resistance genes.
Since plasmids are readily exchanged among S. aureus
strains, we hypothesized that these mobile genetic elements confer significant diversity among closely related USA300
iso-lates. To test this hypothesis, we performed de novo DNA
sequencing of plasmids from previously characterized USA300 clinical isolates.
MATERIALS AND METHODS
USA300 isolates. The USA300 isolates used in this study (Table 1) were previously characterized by comparative whole-genome sequencing,mectyping, spa(protein A) typing, and multilocus sequence typing (13).
DNA isolation and plasmid sequence analyses.Plasmids were isolated from overnight cultures using the Qiagen Maxi prep kit (Valencia, CA) and quan-tified using the Quant-IT Picogreen double-stranded DNA quantification kit (Invitrogen, Carlsbad, CA). DNA and RNA contamination was visualized by agarose gel electrophoresis and ethidium bromide staining. Protein contam-ination was estimated by measuring theA260/A280ratio with a SpectraMax 384
spectrophotometer. Plasmids were digested with EcoRI according to the manufacturer’s instructions (Roche Diagnostics Corporation, Indianapolis, IN), resolved by agarose gel electrophoresis, and visualized with ethidium bromide.
Plasmid nucleotide sequences were determined with a 454 Life Sciences Ge-nome Sequencer FLX (GS FLX) Titanium (Roche) at Agencourt Biosciences (Danvers, MA) following the manufacturer’s recommended procedures. All plas-mids were sequenced to a minimum of 10⫻coverage, and gap closure was performed by designing PCR primers to flanking regions and sequencing the PCR products by capillary Sanger sequencing. The sequencing reads were then assembled into a single contig using DNA Sequencher 4.7 (Gene Codes Corpo-ration, Ann Arbor, MI). The origin of replication for each plasmid was identified using Oriloc software (http://pbil.univ-lyon1.fr/software/Oriloc/oriloc.html) (9), and complete plasmid sequences were annotated by Integrated Genomics (Chi-cago, IL).
Sequence alignment and phylogenetic analysis.Full-length plasmid sequences were aligned according to a multiple sequence alignment program, MAFFT v6.8.14b (12), and a rooted neighbor-joining method tree with 1,000 bootstraps was generated. For each phylogenetic analysis, the most distantly related strain was used as the outgroup. The numbers at nodes are the percentages of boot-straps that supported the branching pattern shown. The scale bar represents
branch length as it relates to the number of substitutions per base position analyzed.
Determination of antibiotic MICs.Broth microdilution (BMD) assays were performed in 96-well microtiter plates using serial 2-fold dilutions of antibiotic (0.125 to 64g/ml). Susceptibility and resistance levels were defined based on recommendations found in Clinical and Laboratory Standards Institute Supple-ment M100-S18 (4). Antibiotics were purchased from Sigma Aldrich (St. Louis, MO) unless otherwise stated. To confirm MIC results from BMD assays, analyses with Etest strips (AB Biodisk, Solna, Sweden) were performed for several anti-biotics according to the manufacturer’s instructions.
Resistance to trimethoprim-sulfamethoxazole, quinupristin-dalfopristin, and tigecycline was monitored by disk diffusion assays on Mueller-Hinton agar. All antibiotic disks were purchased from Fisher, LLC (Pittsburgh, PA). Erythromy-cin-inducible clindamycin resistance was tested using the double disk diffusion assay (8, 21, 22) and a microbroth assay (23) as previously described. In the disk diffusion assay, inducible clindamycin resistance was monitored as a flattening of the zone of inhibition adjacent to the erythromycin disk. Erythromycin and clindamycin disks were purchased from Remel (Lenexa, KS).
Nucleotide sequence accession numbers.All plasmid sequences were depos-ited in the NCBI database with the following accession numbers: LAC-p01 (CP002150), LAC-p03 (CP002149), 18805-p01 (CP002133), 18805-p03 (CP002132), 18806-p03 (CP002134), 18807-p01 (CP002136), 18807-p03 (CP002135), 18808-p01 (CP002138), 18808-p03 (CP002137), 18809-18808-p01 (CP002140), 18809-p03 (CP002139), 18810-p01 (CP002142), 18810-p03 (CP002141), 18811-p01 (CP002144), 18811-p03 (CP002143), 18813-p03 (CP002145), 18813-p04 (CP002146), 19321-p01 (CP002148), and 19321-p03 (CP002147).
RESULTS
Plasmids in USA300 isolates. All USA300 isolates tested contained endogenous plasmids, including at least one large plasmid (27 to 37 kb) (Table 1; see Fig. S1 in the supplemental material), in accordance with the known plasmid contents of strains FPR3757 and USA300TCH_1516 (7, 11). Based upon agarose gel electrophoresis, there were at least four differ-ent plasmid profiles (see Fig. S1 in the supplemdiffer-ental mate-rial), indicating that nucleotide sequence diversity existed among plasmids from closely related strains (e.g., FPR3757 and LAC).
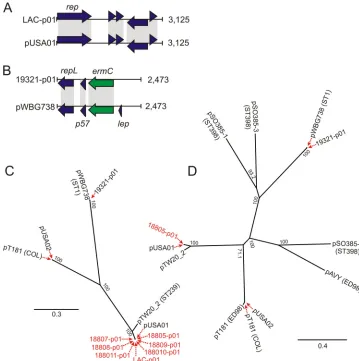
As a step toward understanding the extent of USA300 plasmid diversity, we determined the complete nucleotide sequences of all plasmids from a well-characterized group of USA300 clinical isolates (Table 1). All isolates except 18806 and 18813 had a small 2- to 3-kb plasmid. These small plasmids were nearly identical to pUSA01, a 3.1-kb plasmid of FPR3757 (there were 3 unique plasmid SNPs among all strains) (Fig. 1C and D) (7). Strain 19321 contained a 2.4-kb plasmid (19321-p01) 99.7% identical to pWBG738, a plasmid from a
multilo-cus sequence type (MLST) 1 community-associatedS. aureus
isolate collected from Western Australia (19) (Fig. 1B and C). Plasmid 19321-p01 had three open reading frames (ORFs),
includingermC, which encodes resistance to macrolide,
lin-cosamide, and streptogramin antibiotics (Fig. 1B). As such, strain 19321 was resistant to clindamycin (Table 2), an an-timicrobial agent commonly used to treat CA-MRSA infec-tions (6). Interestingly, plasmid 19321-p01 has 99% se-quence identity to the segment of pUSA03 that contains a
repL homologue (encoding a plasmid replication protein)
andermCand is located between two transposon elements.
[image:2.585.42.284.79.309.2]This plasmid may be an important means for the acquisition of antibiotic resistance elements by USA300. pWBG738 and pUSA01 have no common DNA elements and thus share little sequence similarity (Fig. 1A and B).
TABLE 1. USA300 isolates
Strain Syndrome State No. of plasmids
Size
(kb) Plasmid
FPR3757 Abscess CA 3 37 pUSA03
4 pUSA02 3.1 pUSA01
LAC Abscess CA 2 27 LAC-p03
3.1 LAC-p01
18805 Endocarditis OR 2 27 18805-p03
3.1 18805-p01
18806 Bacteremia GA 1 27 18806-p03
18807 Bacteremia CO 2 27 18807-p03
3.1 18807-p01
18808 Endocarditis NY 2 27 18808-p03
3.1 18808-p01
18809 Bacteremia MN 2 27 18809-p03
3.1 18809-p01 18810 Necrotizing pneumonia PA 2 27 18810-p03 3.1 18810-p01
18811 Osteomyelitis CA 2 27 18811-p03
3.1 18811-p01
18813 Bacteremia TN 2 28 18813-p04
30 18813-p03 19321 Pneumonia/bacteremia OR 2 27 19321-p03 2.4 19321-p01
VOL. 48, 2010 USA300 PLASMIDS 4505
on May 16, 2020 by guest
http://jcm.asm.org/
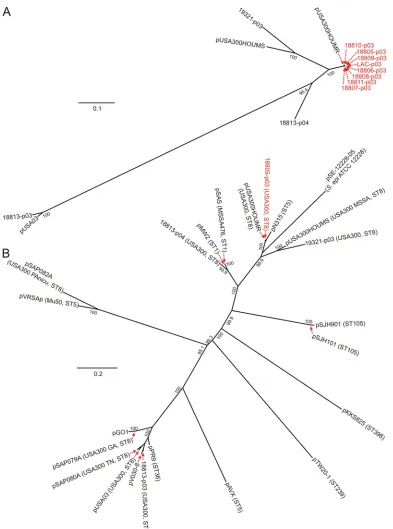
Large (>26-kb) USA300 plasmids.Large plasmids (⬃27 kb) from eight USA300 isolates—those that comprise the epidemic USA300 clone—were essentially identical (99% sequence iden-tity) and also similar to pUSA300HOUMR (11) (Fig. 2 and 3). These plasmids encode several heavy metal and/or antibiotic
resistance elements, including resistance to Cd2⫹,
bacitra-cin, macrolides, penicillin, kanamybacitra-cin, and streptothricin (Fig. 2A).
By comparison, large plasmids from strain FPR3757 (pUSA03) (7), 18813 (18813-p03 and 18813-p04) (13), 19321 (19321-p03) (13), USA300 PAnicu (pSAP082A) (16), USA300 GA (pSAP079A) (16), and USA300 TN (pSAP080A) (16) were divergent from those of the other USA300 isolates (Fig. 2 and 3). Strain 18813 was the only isolate that had two large plasmids (18813-p04 and 18813-p03) (Table 1 and Fig. 2). Plasmid 18813-p03 was 97% identical to pUSA03 but did not
contain anileS(whuch encodes isoleucyl-tRNA synthetase) or
ermCgene (Fig. 2A). As with pUSA03, 18813-p03 encoded the
conjugative transfer elements (tragenes) necessary for
conju-gal transfer of DNA. Plasmid 18813-p04 was a chimera of pN315/pMW2 and plasmids integrated within staphylococcal
cassette chromosome (SCC)mec type III (SCCmecIII) ofS.
aureusstrains V14 and 85/2082 (Fig. 2B). This plasmid
con-tained a cadD-cadX operon (encoding cadmium resistance
protein and regulator) and a-lactamase (bla) resistance
cas-sette also present in pN315 and pMW2 (Fig. 2B) (1, 15). Plasmid 18813-p04 had several elements that confer
resis-tance to heavy metals (e.g., Cd2⫹, Hg2⫹, As2⫹, and Zn2⫹)
(Fig. 2). These elements are found in plasmids integrated
into SCCmecIII of strains V14 and 85/8082 (Fig. 2B). The
meroperon andacr3(which encodes an arsenical-resistance
[image:3.585.109.468.68.429.2]protein) were flanked by a transposon and a gene encoding FIG. 1. Phylogenetic analyses of 3-kb plasmids from USA300 isolates. (A) Comparison of open reading frames in LAC-p01 and pUSA01 (an FPR3757 plasmid). (B) Comparison of open reading frames in 19321-p01 and pWBG738. Arrows and arrowheads indicate ORFs. Green arrows indicateermC, an antibiotic resistance element. Common elements and ORFs are connected by gray shading. (C) Neighbor-joining phylogenetic trees of 3-kb plasmids from USA300 isolates and selectedS. aureusisolates/strains was performed as described in Materials and Methods. Plasmids sequenced in this study are shown in red text. The numbers at each node are the percentages of bootstraps that support the branch. The scale bar represents branch length as it relates to the number of substitutions per base position. (D) Extended phylogenetic analysis (including plasmids fromS. aureusstrains not related to USA300) indicating the relationship between 3-kb plasmids of the closely related USA300 isolates and small plasmids from other sequencedS. aureusisolates/strains. 18805-p01 was used as a representative of the newly sequenced plasmids from the epidemic USA300 clone.
on May 16, 2020 by guest
http://jcm.asm.org/
TABLE 2. Antibiotic susceptibility of USA300 isolates Antibiotic class Antibiotic MIC ( g/ml) a CLSI standard range ( g/ml) b 18805 18806 18807 18808 18809 18810 18811 18813 19321 LAC FPR3757 S I R Aminoglycoside Gentamicin 0.5 1 0.5 0.5 2 0.5 1 1 0.5 2 8 ⬍ 48 ⬎ 16 Kanamycin >64 >64 >64 >64 >64 >64 >64 >64 >64 >64 >64 ⬍ 16 32 ⬎ 64 Tobramycin 212121 1 220 .5 2 ⬍ 48 ⬎ 16 Ansamycin Rifampin ⬍ 0.125 ⬍ 0.125 ⬍ 0.125 ⬍ 0.125 ⬍ 0.125 ⬍ 0.125 ⬍ 0.125 0.25 0.5 0.25 ⬍ 0.125 ⬍ 12 ⬎ 4 Fluoroquinone Ciprofloxacin 32 8 64 0.5 0.125 0.5 >64 0.5 64 >64 64 ⬍ 12 ⬎ 4 Levofloxacin 32 1 32 0.25 0.25 0.25 8 0.5 2 16 16 ⬍ 12 ⬎ 4 Gatifloxacin 2 0.5 4111 2 0 .5 14 2 Glycopeptide Vancomycin 2 2 0.25 2 1 1 2 0.5 2 1 2 ⬍ 2 4–8 ⬎ 16 Teicoplanin 0.25 0.25 0.25 0.25 0.25 0.25 0.25 0.25 ⬍ 0.125 ⬍ 0.125 0.25 ⬍ 81 6 ⬎ 32 Lincosamide Clindamycin 0.5 ⬍ 0.125 0.125 ⬍ 0.125 1 ⬍ 0.125 ⬍ 0.125 ⬍ 0.125 >64 1 >64 ⬍ 0.5 1–2 ⬎ 4 Macrolide Erythromycin 64 64 64 64 >64 64 64 8 64 >64 >64 ⬍ 0.5 1–4 ⬎ 8 Oxazolidinone Linezolid 244444 4 444 4 ⬍ 4 Penicillin Oxacillin 16 16 16 16 32 16 32 32 32 32 32 ⬍ 2 ⬎ 4 Phenicol Chloramphenicol 488888 1 6 32 16 32 16 ⬍ 81 6 ⬎ 32 Pseudomonic acid Mupirocin 212111 1 10 .5 0 .5 >64 Steroid Fusidic acid ⬍ 0.125 0.25 0.25 0.25 ⬍ 0.125 0.25 0.25 0.25 0.25 0.5 0.25 ⬍ 0.5 1 ⬎ 2 c Tetracycline Tetracycline 1 0.5 0.5 0.5 1 4 0.25 ⬍ 0.125 0.5 0.5 64 ⬍ 48 ⬎ 16 Minocycline 0.5 ⬍ 0.125 0.25 0.25 0.25 0.25 0.25 0.25 0.25 0.25 0.5 ⬍ 48 ⬎ 16 Doxycycline 0.5 ⬍ 0.125 0.5 0.25 0.25 0.25 0.25 0.25 0.25 0.25 4 ⬍ 48 ⬎ 16 a All of the USA300 isolates exhibited susceptibility to the antibiotics used in the antibiotic disc assays: the folate pathway inhibitor trimethopri m-sulfamethoxazole, the streptogramins quinupristin-dalfopristin, and the glycylglycine tigecycline. None of the isolates exhibited erythromycin-inducible clindamycin resistance. Values in boldface indicate antib iotic resistance. b S, susceptible; I, intermediate; R, resistant. c Standards in the United Kingdom, Canada, and Australia. Fusidic acid is not approved by the FDA.
VOL. 48, 2010 USA300 PLASMIDS 4507
on May 16, 2020 by guest
http://jcm.asm.org/
[image:4.585.150.439.73.727.2]a recombinase, respectively, suggesting that a recombina-tion event resulted in the mosaic sequence of the plasmid.
Plasmid 19321-p03 was ⬎99% identical to pSK67 (27,420
out of 27,439 bases were identical, with 16 1-base gaps), an orphan family Bla/heavy metal resistance plasmid from an
ar-chaicS. aureusisolate collected from a Melbourne, Australia,
hospital in 1949 (10). These observations are in accordance with comparative whole-genome sequence analysis, which indicates 19321 is divergent from the other isolates compris-ing the epidemic USA300 clone (13). We also discovered that 19321-p03 encoded putative staphylococcal enterotox-ins J, P (or D), and R (Fig. 2A). These enterotoxin genes
have been reported previously in plasmids of otherS. aureus
strains (2, 26).
Antimicrobial agent resistance.To determine whether the antibiotic resistance elements present in plasmids contributed to the resistance patterns of each USA300 isolate, we deter-mined the MIC levels of several clinically relevant antibiotics toward all isolates by using broth microdilution assays (Table
2). As expected, all isolates were resistant to oxacillin (MICⱖ
16g/ml) and erythromycin, although the MIC of
erythromy-cin toward 18813 was considerably lower than that of the other
10 isolates (8g/ml for 18813 versusⱖ64g/ml for all others)
(Table 2). Consistent with these results, plasmids from iso-FIG. 2. Gene contents of USA300 plasmids. (A) Comparison of ORFs carried by the large (⬎26-kb) plasmids from USA300 strains. Antibiotic resistance elements are shown in green, transposase or DNA integration/recombination/inversion elements are in red, conjugal transfer (tra) elements are in yellow, and enterotoxins-encoding elements are in pink. ORFs with no reported function are in white. Selected common DNA elements are connected by light-gray shading. All other ORFs with putative or proven functions are shown in blue. Underlines and accompanying asterisks represent possible homopolymer sequencing errors resulting in frameshifts or truncations of a single ORF. (B) Comparison of ORFs present in plasmid 18813-p04 and those of other sequenced plasmids. TCS, two-component gene regulatory system.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:5.585.42.544.68.505.2]late 18813 lacked macrolide resistance elements (Fig. 2A). FPR3757 and 19321 were constitutively resistant to
clindamy-cin, likely due to ermC present in pUSA03 and 19321-p01,
respectively, and none of the isolates had erythromycin-induc-ible clindamycin resistance (Table 2). Strain FPR3757 was the
only organism resistant to tetracycline, presumably conferred
by the tetracycline resistance element on pUSA02 (MICⱖ64
g/ml for FPR3757 and ⬍ 0.125 to 4 g/ml for all other
strains) (Table 2) (7). However, all isolates were susceptible to minocycline and doxycycline, two antibiotics in the tetracy-FIG. 3. Phylogenetic analyses of large plasmids from USA300 isolates. (A) Neighbor-joining phylogenetic trees of large plasmids from USA300 isolates were generated as described in Materials and Methods. All strains are multilocus sequence type 8 (ST8). Plasmids sequenced in this study are shown in red text. (B) Phylogenetic analysis of large plasmids from diverseS. aureusisolates/strains. The pulsed-field type, common name, and/or multilocus sequence type of each strain (if available) are in parentheses next to the plasmid name. 18805-p03 was used as a representative of the newly sequenced plasmids from the epidemic USA300 clone.
VOL. 48, 2010 USA300 PLASMIDS 4509
on May 16, 2020 by guest
http://jcm.asm.org/
[image:6.585.97.490.64.593.2]cline family used to treat CA-MRSA infections (Table 2). All USA300 isolates were susceptible to trimethoprim-sul-famethoxazole, vancomycin, teicoplanin, tigecycline, and linezolid, antimicrobial agents used as frontline therapy for CA-MRSA infection (Table 2). Strain FPR3757 was the only
organism resistant to mupirocin, as pUSA03 contains theileS
gene, which encodes high-level mupirocin resistance (7). Ad-ditionally, all isolates were susceptible to rifampin and fusidic acid, a current therapeutic compound used in Europe for the treatment of MRSA infections.
DISCUSSION
USA300 infections primarily affect skin and soft tissue, but the pathogen can cause severe invasive syndromes, such as osteomyelitis, sepsis, or pneumonia (reviewed in references 3, 5, and 6). Inasmuch as plasmids provide a relatively efficient means for staphylococci to exchange genetic information, com-prehensive knowledge of plasmid content among USA300 iso-lates is critical for a full understanding of pathogen diversity and virulence and the emergence and spread of antibiotic resistance.
Complete nucleotide sequences of plasmids from at least six USA300 isolates are available for comparative analysis, and these plasmids have varied gene contents (7, 11, 16). Given the relative ease with which plasmids are exchanged among staph-ylococci, there is potential for these mobile elements to con-tribute significantly to genetic diversity among USA300 strains. Indeed, two plasmids reported by McDougal et al. (16), which were recovered from USA300 isolates with unusual antimicro-bial resistance profiles, are similar to pUSA03 (7, 16) and 18813-p03 (Fig. 3B) but relatively divergent from the earlier USA300 isolates that comprise the epidemic clone (except for FPR3757) (Fig. 3B). Such plasmid diversity is likely explained by varied selective pressures imparted to the parent strain. For example, acquisition of plasmid-mediated antibiotic resistance in USA300 is far more common in health care-associated iso-lates than in those from the community (16).
Several USA300 isolates—LAC, USA300_TCH1516, and 18805 to 18811—contained essentially identical plasmids, a finding consistent with the high level of similarity among the core genome sequences of these isolates (11, 13). These data provide further support for the idea that there has been recent clonal emergence of the epidemic USA300 strain. Given these findings, the large plasmid and/or a subset of genes contained within it could be used for molecular epidemiology studies. Whether elements encoded by one or more of the plasmids described here contributed to the emergence or epidemicity of the USA300 clone is unknown. None of the genes encoding proteins with known or putative functions appear uniquely suited to promote the success of USA300. However, there are plasmid open reading frames that encode hypothetical proteins with uncharacterized functions (Fig. 2). Nevertheless, the core genomes of FPR3757 and LAC are virtually identical, but these isolates contain fairly divergent large plasmids (Fig. 2A and 3), an observation that suggests the large plasmid is not key to the emergence and epidemicity of USA300.
In addition to facilitating the acquisition of antibiotic resistance elements, plasmids potentially provide an effi-cient means to transfer virulence factor genes among
staph-ylococci. For example, exfoliative toxin B, a causative agent of staphylococcal scalded skin syndrome, is plasmid encoded (25). Interestingly, plasmid 19321-p03 contained genes en-coding SER, SEJ, and SEP (or D) (Fig. 2A). The
entero-toxin genessed, sej, and ser are carried on plasmids from
Staphylococcus aureusisolates associated with outbreaks of
food poisoning (2, 20, 26). The contribution of the plasmid-encoded enterotoxins to USA300 pathogenesis, if any, re-mains to be determined.
ACKNOWLEDGMENTS
This work was supported by the Intramural Research Program of the National Institute of Allergy and Infectious Diseases, National Insti-tutes of Health.
The authors declare no conflicts of interest.
REFERENCES
1.Baba, T., F. Takeuchi, M. Kuroda, H. Yuzawa, K. Aoki, A. Oguchi, Y. Nagai, N. Iwama, K. Asano, T. Naimi, H. Kuroda, L. Cui, K. Yamamoto, and K. Hiramatsu.2002. Genome and virulence determinants of high virulence community-acquired MRSA. Lancet359:1819–1827.
2.Bayles, K. W., and J. J. Iandolo.1989. Genetic and molecular analyses of the gene encoding staphylococcal enterotoxin D. J. Bacteriol.171:4799– 4806.
3.Chambers, H. F., and F. R. DeLeo.2009. Waves of resistance:Staphylococcus aureusin the antibiotic era. Nat. Rev. Microbiol.7:2464–2474.
4.Clinical and Laboratory Standards Institute.2008. Performance standards for antimicrobial susceptibility testing; 18th informational supplement. Clin-ical and Laboratory Standards Institute, Wayne, PA.
5.DeLeo, F. R., and H. F. Chambers.2009. Reemergence of antibiotic-resistantStaphylococcus aureusin the genomics era. J. Clin. Invest.119: 2464–2474.
6.DeLeo, F. R., M. Otto, B. N. Kreiswirth, and H. F. Chambers.2010. Community-associated meticillin-resistantStaphylococcus aureus. Lancet375:1557–1568. 7.Diep, B. A., S. R. Gill, R. F. Chang, T. H. Phan, J. H. Chen, M. G. Davidson,
F. Lin, J. Lin, H. A. Carleton, E. F. Mongodin, G. F. Sensabaugh, and F. Perdreau-Remington.2006. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistantStaphylococcus aureus. Lancet367:731–739.
8.Fiebelkorn, K. R., S. A. Crawford, M. L. McElmeel, and J. H. Jorgensen. 2003. Practical disk diffusion method for detection of inducible clindamycin resistance in Staphylococcus aureus and coagulase-negative staphylococci. J. Clin. Microbiol.41:4740–4744.
9.Frank, A. C., and J. R. Lobry.2000. Oriloc: prediction of replication bound-aries in unannotated bacterial chromosomes. Bioinformatics16:560–561. 10.Gillespie, M. T., J. W. May, and R. A. Skurray.1985. Antibiotic resistance in
Staphylococcus aureusisolated at an Australian hospital between 1946 and 1981. J. Med. Microbiol.19:137–147.
11.Highlander, S. K., K. G. Hulten, X. Qin, H. Jiang, S. Yerrapragada, E. O. Mason, Y. Shang, T. M. Williams, R. M. Fortunov, Y. Liu, O. Igboeli, J. Petrosino, M. Tirumalai, A. Uzman, G. E. Fox, A. M. Cardenas, D. M. Muzny, L. Hemphill, Y. Ding, S. Dugan, P. R. Blyth, C. J. Buhay, H. H. Dinh, A. C. Hawes, M. Holder, C. L. Kovar, S. L. Lee, W. Liu, L. V. Nazareth, Q. Wang, J. Zhou, S. L. Kaplan, and G. M. Weinstock.2007. Subtle genetic changes enhance virulence of methicillin resistant and sensitive Staphylococ-cus aureus. BMC Microbiol.7:99.
12.Katoh, K., K. Misawa, K. Kuma, and T. Miyata.2002. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier trans-form. Nucleic Acids Res.30:3059–3066.
13.Kennedy, A. D., M. Otto, K. R. Braughton, A. R. Whitney, L. Chen, B. Mathema, J. R. Mediavilla, K. A. Byrne, L. D. Parkins, F. C. Tenover, B. N. Kreiswirth, J. M. Musser, and F. R. DeLeo.2008. Epidemic com-munity-associated methicillin-resistant Staphylococcus aureus: recent clonal expansion and diversification. Proc. Natl. Acad. Sci. U. S. A. 105:1327–1332.
14.King, M. D., B. J. Humphrey, Y. F. Wang, E. V. Kourbatova, S. M. Ray, and H. M. Blumberg.2006. Emergence of community-acquired methicillin-resis-tantStaphylococcus aureusU. S. A. 300 clone as the predominant cause of skin and soft-tissue infections. Ann. Intern. Med.144:309–317.
15.Kuroda, M., T. Ohta, I. Uchiyama, T. Baba, H. Yuzawa, I. Kobayashi, L. Cui, A. Oguchi, K. Aoki, Y. Nagai, J. Lian, T. Ito, M. Kanamori, H. Matsumaru, A. Maruyama, H. Murakami, A. Hosoyama, Y. Mizutani-Ui, N. K. Taka-hashi, T. Sawano, R. Inoue, C. Kaito, K. Sekimizu, H. Hirakawa, S. Kuhara, S. Goto, J. Yabuzaki, M. Kanehisa, A. Yamashita, K. Oshima, K. Furuya, C. Yoshino, T. Shiba, M. Hattori, N. Ogasawara, H. Hayashi, and K. Hira-matsu.2001. Whole genome sequencing of meticillin-resistant Staphylococ-cus aureus. Lancet357:1225–1240.
on May 16, 2020 by guest
http://jcm.asm.org/
16.McDougal, L. K., G. E. Fosheim, A. Nicholson, S. N. Bulens, B. M. Limbago, J. E. Shearer, A. O. Summers, and J. B. Patel.2010. Emergence of resistance among USA300 methicillin-resistant Staphylococcus aureus isolates causing invasive disease in the United States. Antimicrob. Agents Chemother.54: 3804–3811. doi:AAC.00351–10 [pii];10.1128/AAC. 00351–10.
17.McDougal, L. K., C. D. Steward, G. E. Killgore, J. M. Chaitram, S. K. McAllister, and F. C. Tenover. 2003. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: establishing a national database. J. Clin. Microbiol.41: 5113–5120.
18.Moran, G. J., A. Krishnadasan, R. J. Gorwitz, G. E. Fosheim, L. K. McDou-gal, R. B. Carey, and D. A. Talan. 2006. Methicillin-resistantS. aureus infections among patients in the emergency department. N. Engl. J. Med. 355:666–674.
19.O’Brien, F. G., Z. Zaini, G. W. Coombs, J. C. Pearson, K. Christiansen, and W. B. Grubb.2005. Macrolide, lincosamide and streptogramin B resistance in a dominant clone of Australian community methicillin-resistant Staphy-lococcus aureus. J. Antimicrob. Chemother.56:985–986.
20.Omoe, K., D.-L. Hu, H. Takahashi-Omoe, A. Nakane, and K. Sinagawa. 2003. Identification and characterization of a new staphylococcal entertoxin-related putative toxin encoded by two kinds of plasmids. Infect. Immun. 71:6088–6094.
21.O’Sullivan, M. V. N., Y. Cai, F. Kong, X. Zeng, and G. L. Gilbert.2006.
Influence of disk separation distance on accuracy of the disk approximation test for detection of inducible clindamycin resistance inStaphylococcusspp. J. Clin. Microbiol.44:4072–4076.
22.Steward, C. D., P. M. Raney, A. K. Morrell, P. P. Williams, L. K. McDougal, L. Jevitt, J. E. McGowan, Jr., and F. C. Tenover.2005. Testing for induction of clindamycin resistance in erythromycin-resistant isolates of Staphylococ-cus aureus. J. Clin. Microbiol.43:1716–1721.
23.Swenson, J. M., W. B. Brasso, M. J. Ferraro, D. J. Hardy, C. C. Knapp, L. K. McDougal, L. B. Reller, H. S. Sader, D. Shortridge, R. Skov, M. P. Wein-stein, B. L. Zimmer, and J. B. Patel.2007. Detection of inducible clindamy-cin resistance in staphylococci by broth microdilution using erythromyclindamy-cin- erythromycin-clindamycin combination wells. J. Clin. Microbiol.45:3954–3957. 24.Tenover, F. C., L. K. McDougal, R. V. Goering, G. Killgore, S. J. Projan, J. B.
Patel, and P. M. Dunman.2006. Characterization of a strain of community-associated methicillin-resistantStaphylococcus aureuswidely disseminated in the United States. J. Clin. Microbiol.44:108–118.
25.Warren, R., M. Rogolsky, B. B. Wiley, and L. A. Glasgow.1975. Isolation of extrachromosomal deoxyribonucleic acid for exfoliative toxin produc-tion from phage group II Staphylococcus aureus. J. Bacteriol.122:99– 105.
26.Zhang, S., J. J. Iandolo, and G. C. Stewart.1998. The enterotoxin D plasmid ofStaphylococcus aureusencodes a second enterotoxin determinant (sej). FEMS Microbiol. Lett.168:227–233. doi:S0378-1097(98)0042–6.
VOL. 48, 2010 USA300 PLASMIDS 4511