A model for the compressible, viscoelastic behavior of human amnion addressing tissue variability through a single parameter
Full text
Figure
![Fig. 1 Comparison of lateral in-plane (λ2) and out-of-plane(λ3) stretches vs. stretch in loading direction in uniaxial ten-sion for the incompressible model with planar fibers [B¨urzleand Mazza, 2013] and the refined, compressible model withslightly inclined](https://thumb-us.123doks.com/thumbv2/123dok_us/8652354.373790/6.595.56.255.103.328/comparison-stretches-direction-incompressible-rened-compressible-withslightly-inclined.webp)

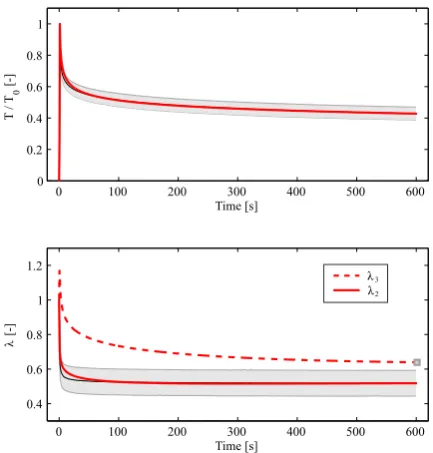
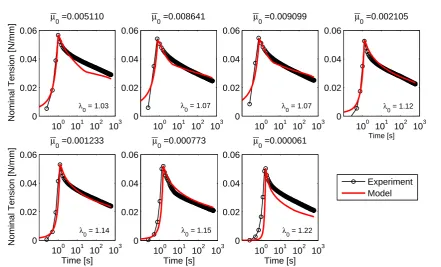
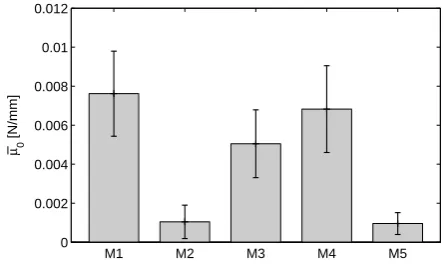
![Fig. 7 Prediction of creep in planar equibiaxial tension basedon the parameters in Table 1 and comparison with the exper-imentally observed ratio of apex displacement in creep infla-tion experiments [Mauri et al, 2015c].](https://thumb-us.123doks.com/thumbv2/123dok_us/8652354.373790/11.595.49.269.325.438/prediction-equibiaxial-parameters-comparison-imentally-observed-displacement-experiments.webp)
Related documents
Method: This study investigates patients with type 2 diabetes and hypertension for which concentrations of total high-density lipoprotein (HDL) and low-density lipoprotein
In order to estimate whether starting in different positions of model state space has an impact on predictability, we calculated the anomaly correlation and NRMSE metrics again but
It examined the determinants of female's involvement and commitment in the higher education institutions in Saudi Arabia and the relationship between females'
Experiments were designed with different ecological conditions like prey density, volume of water, container shape, presence of vegetation, predator density and time of
Race condition is an error in clinical programming software which results when two processes try to access the same resource and the result depends on the transaction of
Passed time until complete analysis result was obtained with regard to 4 separate isolation and identification methods which are discussed under this study is as
Therefore, no agreement exists among the groups as to the legitimate methods for resolving conflict; this leads to military interference in politics that most commonly takes the
