ABSTRACT
The neurobiological effects of binge-like alcohol consumption during adolescence
include reduced frontolimbic connectivity later in adulthood, which may result in decreased inhibitory control, impulsive behavior, and increased likelihood of alcoholism or alcohol abuse
later in life. Animal behavior in Pavlovian Conditioned Approach (PCA) can be classified as sign-tracking (approach to the cue) or goal-tracking (approach to the reward delivery site). Sign-tracking behavior demonstrated during PCA shares many behavioral characteristics with
vulnerability to drug or alcohol addiction and abuse and is, thus, an appropriate model to investigate the neurobiological effects of binge alcohol consumption in adolescents. In the
present study, we modeled binge-like alcohol consumption in adolescent rats to determine how adolescent intermittent ethanol (AIE) exposure alters behavior in PCA. AIE-exposed rats exhibited decreased goal-tracking behavior compared to controls, indicating that adolescent
alcohol consumption alters reward-associated behavior. We applied optogenetic techniques to determine the effects of stimulating orbitofrontal cortex (OFC) neurons projecting to the nucleus
accumbens (NAcc) on conditioned approach behavior. Data showed that stimulation of the accumbens pathway does not reliably alter behavior in AIE-exposed rats. Thus, the orbito-accumbens pathway is likely not a desirable target to reverse the long-term effects of binge
alcohol consumption during adolescence that are demonstrated by conditioned-approach behavior in PCA. However, further research is needed to thoroughly investigate the role of this
pathway in addiction-like behavior in additional subjects.
INTRODUCTION
interference with brain development. Notably, 33% of teenagers report having at least one alcoholic drink by age 15, and over 5 million young people aged 12 to 20 report at least one
binge drinking episode (five or more drinks for males and four or more drinks for females within a few hours) in the past month1. In addition to the negative short-term effects of binge drinking during adolescence, there are consequences related to brain development that may affect
addiction patterns and alcohol use throughout adulthood. Binge-like alcohol consumption can inhibit the structural and functional connectivity changes that occur in the prefrontal cortex and
frontolimbic pathways in rats during adolescence (Broadwater et al., 2017). Decreased frontolimbic connectivity may result in reduced inhibitory control, impulsive behavior, and increased likelihood of alcoholism or alcohol abuse later in adulthood (Broadwater et al., 2017).
In the present study, we model binge-like alcohol consumption in adolescent rats to determine how alcohol exposure alters behavior in Pavlovian conditioned approach (PCA). In
PCA training, a conditioned stimulus, such as a lever and light, is presented prior to an unconditioned stimulus, such as a sucrose reward. Though the reward is not contingent on the rats’ behavior during cue presentation, some rats will display a preference for interaction either
with the conditioned stimulus or with the reward site. Specifically, rats that display sign-tracking behavior tend to interact more with the cue (conditioned stimulus), while rats that display
goal-tracking behavior tend to interact more with the reward site. Adolescent intermittent ethanol (AIE) exposed rats exhibit decreased goal-tracking behavior (Madayag et. al, 2017) or increased sign-tracking behavior (McClory and Spear, 2014) compared to control rats, indicating that
adolescent alcohol consumption alters reward-associated behavior. Sign-tracking behavior demonstrated during PCA shares behavioral characteristics with drug or alcohol addiction and
abuse, such as long-term retention, spontaneous recovery, and impulsivity regarding the conditioned response (Tomie et al., 2007). Thus, PCA is an appropriate model to investigate the
neurobiological effects of binge alcohol consumption in adolescents and subsequent patterns of addiction-like behavior.
We are interested in the brain circuits that drive behavior and how those circuits may be
disrupted by AIE exposure. One way to manipulate neural circuits is through optogenetics, an experimental technique developed over the past decade to activate or silence specific neurons in
vivo with precise timing. Optogenetics allows for precise control of activity in specific types of neurons. Targeted neurons are genetically modified to express light-sensitive proteins called opsins, typically in the form of ion channels. When opsins are exposed to light of a specific
wavelength, they undergo a conformational change that results in activation or silencing of the targeted neuron (Guru et al., 2015). Optogenetics may be used to determine the contributions of
specific neuronal circuits and pathways and to develop an understanding of various neurological processes associated with reward-seeking and compulsive behaviors. For example, a study by Burguiere and colleagues (2013) found that optogenetic stimulation of the pathway from the
lateral orbitofrontal cortex to the medial striatum may suppress compulsive behaviors. Researchers deleted a gene known to prevent repetitive self-grooming in mice, thereby inducing
a compulsive behavior in these animals. Optogenetic stimulation of the orbitofrontal-striatal pathway restored down-regulation of striatal projections and effectively inhibited repetitive behavior (Burguiere et al., 2013).
In the present study, we applied optogenetic techniques to determine the effects of stimulating orbitofrontal cortex (OFC) neurons projecting to the nucleus accumbens (NAcc) on
ethanol exposure in adolescents, and there is evidence that AIE exposure affects the orbito-accumbens pathway. For example, compared to controls, AIE-exposed rats exhibited decreased
activation of the OFC and increased activation in the NAcc after alcohol challenge, as indicated by expression of the immediate early gene c-Fos (Liu and Crews 2015). Though the effects of AIE exposure may permanently alter subjects’ neurobiology, we are interested in whether
optogenetic manipulation may reverse some of the behavioral effects. It is known that OFC neurons are more active during goal-tracking behaviors such as approaching the reward
receptacle during cue delivery than during sign-tracking behaviors (Stringfield et al., 2017). Therefore, we hypothesized that optogenetic stimulation of neuronal projections from the OFC to the NAcc would promote goal-tracking behavior and reverse some of the behavioral effects of
AIE exposure. In this study, we found that AIE exposure reduced goal-tracking behavior in PCA, but in vivo optogenetic activation of the orbito-accumbens pathway did not reliably promote
goal-tracking behavior or reverse the effects of AIE exposure.
METHODS
Animals
The University of North Carolina Institutional Animal Care and Use Committee approved all experimental procedures.
Female Sprague-Dawley rats acquired from in-house breeders were used in this
experiment. A previous study by Madayag and colleagues found that female rats showed enhanced sign-tracking behavior following AIE exposure compared to males (Madayag et al.,
temperature and humidity. Female littermates were weaned to two per cage on postnatal day (P) 21. From P25-P54, AIE rats received 5 g/kg intragastric ethanol once per day following a
2-days-on, 2-days-off schedule. Control (CON) rats received water in place of ethanol following the same regimen. Rats received a total of 16 AIE or water exposures. Rats continued to be housed two per cage for the first 3 weeks of behavioral training and were then separated into
individual cages.
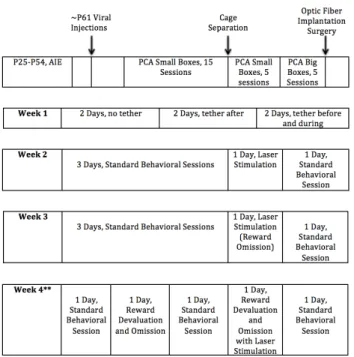
Figure 1: Timeline of Surgery and Behavioral Training
Rats followed a standard schedule of AIE or water exposure, surgery, behavioral training, and laser stimulation. Note: Only a subset of rats (n=7) underwent the reward devaluation protocol shown for Week 4.
Viral Injections and Optic Fiber Implantation Surgery
On ~P61, rats were anesthetized with 5% isoflurane and were maintained on 2-3%
isoflurane throughout the surgery. Viral injections were administered to the lateral OFC, 3.8 mm anterior and 2.0 mm lateral from bregma, and 4.2 mm ventral from the adjacent skull surface. The control (inactive) virus, AAV5: CaMKIIa-> EYFP, resulted in expression of only the
stored at -80 degrees Celsius were thawed on ice prior to the surgery and added to PBS to achieve a final concentration of 1 x 1012 pfu/mL. Injections were administered in the lOFC with a 27ga cannula connected to a microsyringe. One microliter was infused into each hemisphere at a rate of 0.1 microliter per minute. Twenty minutes after the injection start, the cannula was removed and rats were sutured. Rats recovered for 7-10 days and were given subcutaneous
meloxicam injections (50 mg/kg) for analgesia every day for three days. Sutures were removed and rats received a 100 mL bottle of 20% sucrose overnight to familiarize them with the reward
solution. PCA training sessions began the following day.
For the optic fiber implantation surgery, rats were again anesthetized with isoflurane. The NAcc was targeted with coordinates of 1.7 mm anterior and 2.0 mm lateral from bregma, and 4.2
mm ventral from the adjacent skull surface. Optic fibers were implanted bilaterally. A connector was placed for later use with the tether and patch cables. The optic fiber and connector
placement was secured with a headcap of dental cement and four screws into the skull. Rats followed the same post-operative recovery procedure as described for the viral injection surgeries.
Pavlovian Conditioned Approach
Rats began behavioral training ~P65 in small operant boxes (12” L x 12.5” W x 11.5”H)
with a house light and white noise present consistently throughout the behavioral session (Med Associates, St. Albans, VT). To familiarize rats with the reward solution, a bottle of 20% sucrose was administered in their home cage the day before training began. On the first day of behavioral
training, rats received receptacle training in the small operant boxes; 0.1 mL of 20% sucrose was administered at time intervals varying from 120 to 230 seconds. Upon completion of the session,
At ~P70, rats began PCA training. Rats were placed in the operant boxes with the house light off for 5 minutes prior to the start of the session to allow for habituation. When the session
began, a conditioned stimulus consisting of an extended lever and cue light was presented for 30 seconds prior to the delivery of a 0.1 mL 20% sucrose reward in the receptacle. After a variable interval of 90 to 210 seconds, the cue was presented again, followed by reward delivery. Each
behavioral session consisted of 15 trials. Receptacle entries were measured with a photobeam in the reward receptacle and the number of lever presses was recorded during the presentation of
the cue.
Experimental Procedure and Laser Stimulation
We used a laser controlled by an Arduino board and MedPC software to directly
stimulate the orbito-accumbens pathway in rats expressing channelrhodopsins. The laser administered light at the wavelength 473 nm in pulses with a frequency of 20 Hz for 500 ms
every 2s throughout the 30s duration of the cue. Rats received laser stimulation for three behavioral sessions: standard PCA, reward omission, and reward devaluation. In the reward omission session, rats received no reward upon termination of the cue. For reward devaluation,
Figure 2. Laser Stimulation during Pavlovian Conditioned Approach
(A) Between trials, no cue was presented. (B) At the beginning of each trial, cue onset consisted of illumination of a light and the extension of a lever. Throughout the 30s duration of the cue, the rat received stimulation from laser pulses every 2 seconds with a wavelength of 473 nm, frequency of 20 Hz, and duration of 500 ms.
Histological Analysis
Upon completion of the experiment, rats were anesthetized with urethane and perfused
using 4% PBS followed by 4% paraformaldehyde in 0.05M PB. Brains were removed from the rats and incubated in 4% paraformaldehyde in 0.05M PB overnight at 4 degrees Celsius. Brains
were then moved to 30% sucrose in PBS for 2-3 days and then rapidly frozen. OFC slices were mounted on a cover slip with fluorescent mounting medium. The edges of the coverslip were sealed, allowed to dry, and stored in the freezer. NAcc slices were stained and mounted with a
cover slip. The viral injection sites in OFC slices were viewed with an epifluorescence microscope to determine the viral spread. NAcc slices, where the optic fiber was located, were
Behavioral Analysis
The operant box software digitally recorded behavioral data, including lever presses,
receptacle elevation score, latency to lever press, latency to receptacle entry, probability of lever press in a trial, and probability of receptacle entry in a trial. Lever presses were counted during cue presentation, and the receptacle elevation score was calculated as the number of receptacle
entries during cue delivery minus the number of receptacle entries in the 30 seconds prior to the cue. Latency to lever press and receptacle entry measured the time it took for a ratl to press the
lever or enter the receptacle after the onset of the cue.
Ex Vivo Slice Electrophysiology
Ex vivo slice electrophysiology was used to confirm that laser stimulation could
successfully induce action potentials in ChR2-positive OFC neurons that project to the NAcc. The active virus containing a vector for ChR2 and EYFP protein was infused into the lateral
OFC of adult, male rats. Whole cell recordings were performed from ChR2-positive lOFC neurons in current-clamp mode using a K-gluconate based internal in order to induce action potentials with laser stimulation. ChR2-positive neurons were stimulated with LED light at 473
nm for 10 pulses of 5 ms duration. Functional connectivity of the orbito-accumbens pathway was demonstrated with whole cell recordings in the NAcc. Recordings were taken in current clamp
RESULTS
AIE Exposure Decreases Goal-Tracking Behavior in a Standard PCA Session
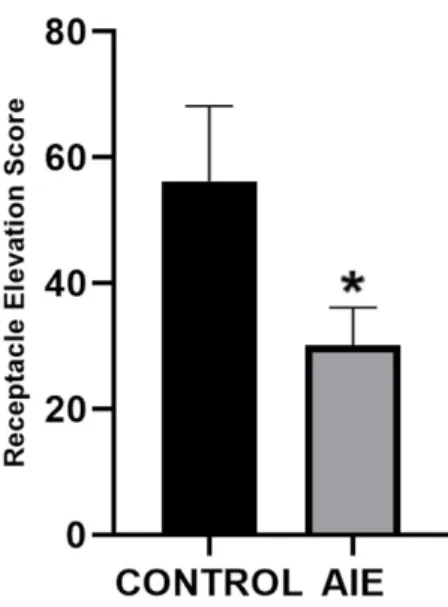
Our hypothesis was centered on the prediction that AIE-exposure results in decreased goal-tracking behavior in PCA, as previously reported. One way to measure goal-goal-tracking behavior is by analyzing the receptacle elevation score, which calculates how the presentation of the cue
affects the rat’s interaction with the reward delivery receptacle. A higher receptacle elevation score indicates that the rat entered the reward delivery receptacle significantly more during the
presentation of the cue than when the cue was not presented, indicating conditioned approach and goal-tracking behavior. In a standard PCA session, rats in the AIE group had significantly lower elevation scores (D=0.4361, p=0.045) than control-exposed rats (Figure 3), suggesting that
AIE exposure results in decreased goal-tracking behavior. No significant differences between groups were observed for any other behavioral metrics.
Figure 3. AIE-exposed rats show decreased receptacle elevation score compared to controls
Reward Devaluation Decreases Goal Tracking Behavior Similarly in AIE and Control Rats As described above, we observed that AIE exposure results in decreased goal-tracking
behavior in PCA. To further explore the effects of AIE exposure in these subjects, we performed a reward devaluation test in which rats had ad libitum access to the sucrose reward solution for one hour prior to the behavioral session. During the behavioral session, no reward was given
following termination of the cue. Previous studies showed that AIE-exposed female rats demonstrated habitual behavior following reward devaluation (Barker et al., 2017), and that
reward devaluation increases sign-tracking behavior in PCA (Morrison et al., 2015). Based on these findings, we predicted that AIE-exposed rats would show behavioral differences from controls in response to reward devaluation. Specifically, the literature suggested that
AIE-exposed rats would be less likely to alter their behavior as a result of reward devaluation, or would show increased sign-tracking behavior.
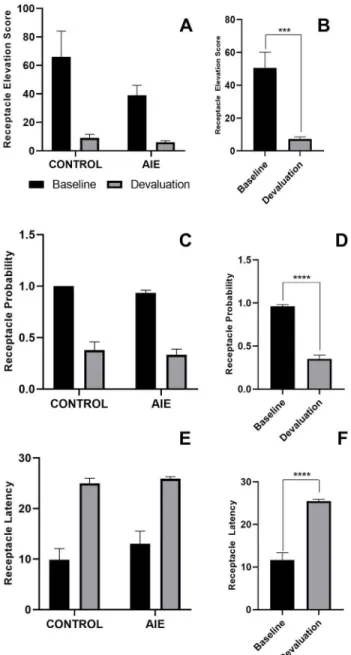
Decreased goal-tracking behavior can be demonstrated by a decrease in receptacle elevation score, a decrease in receptacle probability, or an increase in receptacle latency. Our results showed a significant change in all behavioral metrics consistent with decreased
goal-tracking behavior, but no differences were observed between AIE (n=4) and control (n=3) rats (Figure 4). Two-way ANOVA revealed that receptacle elevation score decreased with reward
devaluation for both AIE and control rats, with a main effect of day (F(1,5)=26.38, p=0.0037) but no main effect of exposure or interaction between day and exposure. Similarly, the
probability of receptacle entry decreased with reward devaluation. A two-way ANOVA revealed
a main effect of day (F(1,5)=196.4, p<0.0001), but no main effect of exposure or interaction between variables. Finally, a decrease in goal-tracking behavior was demonstrated by 12 an
day (F(1,5)=87.11, p=0.0002), but no main effect of exposure or interaction between variables. The main effect of day for each behavioral metric is demonstrated by figures that collapsed data
from all rats to baseline vs. devaluation behavioral sessions (Figure 4BDF). Data analysis did not reveal any trends regarding the effects of AIE exposure or reward devaluation on sign-tracking behavior.
Figure 4. AIE and control rats do not differ in their responses to reward devaluation in a PCA behavioral session
Note: Exposure groups are combined in panels B, D, and F.
Receptacle Elevation Score (A,B): A two-way ANOVA revealed a significant decrease in receptacle elevation score for both groups between the baseline and devaluation days with a main effect of day (B) (F(1,5)=26.38, p=0.0037), but no main effect of exposure or interaction between variables (A).
Receptacle Probability (C,D): A two-way ANOVA revealed a significant decrease in receptacle probability for both groups between the baseline and devaluation days with a main effect of day (E) (F(1,5)=196.4, p<0.0001), but no main effect of exposure or interaction between variables (D).
Confirmation of Orbito-accumbens Pathway Activation
Before investigating the behavioral effects of in vivo optogenetic activation of the
orbito-accumbens pathway during a PCA behavioral session, we confirmed that laser stimulation successfully induced these neurons to fire. We verified the function of channelrhodopsin protein channels in OFC neurons with ex vivo slice electrophysiology data collected by Dr. Aric
Madayag and Dr. Dipa Pati). Electrophysiological recordings were taken from five lateral OFC neurons. Of these five, four fired with 100% spike fidelity for 10 pulses of laser stimulation at 20
Hz and 473 nm (Figure 4A). In the Nacc, all five recorded neurons showed excitatory postsynaptic potentials (EPSPs) with two pulses of laser stimulation. Each pulse was 5ms in duration and 50 ms apart. The EPSP elicited by the first pulse was larger than that of the second
pulse, with average amplitudes of 9.754 mV and 3.546 mV respectively (Figure 4B). These results indicate that laser stimulation at a wavelength of 473 nm successfully activates OFC
neurons projecting to the NAcc.
(A) Representative trace of a ChR2-positive lateral OFC neuron firing in response to 10 pulses of 20 Hz of 473 nm LED light. Scale: X- axis: 1000ms; Y-axis: 25 mV. (B) Representative trace of a nucleus accumbens neuron firing in response to stimulation of ChR2-positive fibers. Scale: X-axis: 250ms; Y-X-axis: 5 mV (Madayag, A., and Pati, D.).
Histology
To ensure that the orbito-accumbens pathway was selectively targeted in all subjects, we
used epifluorescent microscopy to view viral spread throughout orbitofrontal cortex projection
neurons and the precision of the optic fiber implantation in the core of the nucleus accumbens.
After the last behavioral session, we performed histology on all rats’ brains to confirm viral expression and optic fiber placement. This verified that all rats included in data analysis had expressed channelrhodopsin proteins in OFC projection neurons and that the orbito-accumbens
pathway had been successfully activated. An example histology from an AIE-ChR2 rat is provided below (Figure 6). After excluding rats with insufficient viral spread or inaccurate optic
fiber placement, only seven rets met the criteria for inclusion in data analysis. Three of these rats were control-exposed, and four were AIE-exposed.
Figure 6. Histology to verify viral expression and optic fiber placement
rats with sufficient viral expression and correct optic fiber placement were included in data analysis.
Orbito-accumbens Activation Does Not Reliably Reverse Effects of AIE Exposure, but May Affect Behavioral Response to Laser Stimulation
As demonstrated by Figure 3, AIE-exposed rats had a lower receptacle elevation score in a standard PCA session compared to controls, which is indicative of reduced goal-tracking behavior compared to controls. Based on previous evidence that the orbitofrontal cortex and the
nucleus accumbens play a role in goal-tracking behavior (Liu & Crews, 2015 and Stringfield et al., 2017), we hypothesized that in vivo optogenetic activation of the orbito-accumbens pathway
could potentially reduce the effects of AIE exposure. However, as shown in Figure 7, laser stimulation of the orbito-accumbens pathway failed to significantly increase receptacle elevation scores for rats that received the active channelrhodopsin (ChR2) virus. An initial repeated
measures ANOVA revealed that there was no main effect of exposure or interactions between exposure, virus, and day. Considering this lack of exposure effect, we grouped subjects based on
whether they received the active or inactive virus. We then performed a repeated measures ANOVA to observe interactions between laser stimulation and virus type (Figure 7, inset panel). The repeated measures ANOVA yielded an interaction of day by virus (F(1,21)=5.618,
p=0.0274), wherein the laser stimulation day differed from baseline day only in the inactive virus group. For this group, the receptacle elevation score decreased with laser stimulation compared
Figure 7. Effect of laser stimulation on receptacle elevation score
No significant differences were observed between groups when comparing the effects of laser stimulation on receptacle elevation score, so results were collapsed in order to compare the effects of laser stimulation based only on type of virus: active (n=7) or inactive (n=16). After collapsing experimental groups by active or inactive virus (inset panel), a repeated measures ANOVA revealed no main effect of day or virus, but revealed a significant interaction between day and virus in the inactive group. Animals that received the inactive virus had significantly lower receptacle elevation scores with laser stimulation compared to a baseline PCA session (F(1,21)=5.62, p=0.0274).
Orbito-accumbens Activation Does Not Increase Behavioral Flexibility During Reward Omission Task
AIE exposure has been shown to increase sign-tracking behavior or reduce goal-tracking
behavior as a result of neurobiological changes in functional connectivity. A 2014 study by Gass 17 and colleagues found that adolescent alcohol exposure reduces behavioral flexibility,
observed as resistance to extinction (Gass et al., 2014) or reward omission. Based on these
findings, we predicted that AIE-exposed rats would demonstrate similar behavior during a reward omission session compared to a baseline behavioral session. Alternatively, we predicted
that control rats would be more likely to alter their behavior in response to reward omission, thereby demonstrating increased behavioral flexibility. In addition, we predicted that activation of the orbito-accumbens pathway would reverse the effects of AIE exposure, so we expected
and reward omission sessions as a result of increased behavioral flexibility caused by activation of the orbito-accumbens pathway.
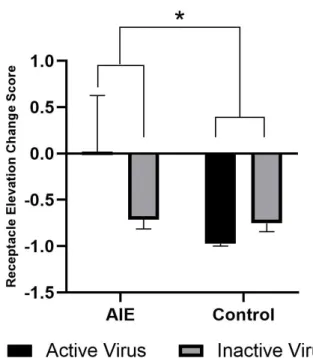
Behavioral changes between baseline and reward omission sessions are shown by change scores, or the percent change in receptacle elevation score between sessions. A more negative change score shows a lower tendency to approach the reward receptacle following cue
termination when reward is not present compared to a baseline session. This metric represents behavioral flexibility. A two-way ANOVA revealed a main effect of exposure (F(1,19)=4.488,
p=0.0475), where AIE-exposed rats had less negative receptacle elevation change scores on average compared to controls, regardless of whether they received the active or inactive virus. There was no main effect of virus, meaning that activation of the orbito-accumbens pathway
during the reward omission task did not affect receptacle elevation change scores. Two-way ANOVA did not reveal any interaction between virus and exposure.
Figure 8. Laser stimulation does not alter sensitivity to reward omission in PCA.
revealed a main effect of exposure, with AIE-exposed rats exhibiting a significantly smaller overall decrease in receptacle elevation score (F(1,19)=4.488, p=0.0475). There was no main effect of virus and no significant interaction between virus and exposure.
DISCUSSION
In this study, we first examined the behavioral differences between AIE-exposed rats and controls. Compared to controls, AIE-exposed rats exhibited significantly lower receptacle
elevation scores in a standard PCA session. Our findings are consistent with a previous study (Madayag et. al, 2017) that discovered that AIE exposure reduced goal-tracking behavior in
PCA. However, data analysis did not reveal any significant differences in sign-tracking
behavioral metrics for AIE and control animals, which contrasts with a previous study that found that AIE exposure resulted in increased sign-tracking behavior (McClory and Spear, 2014).
Behavioral effects of AIE exposure were not observed in a PCA session with reward devaluation. Results showed a significant decrease in all behavioral metrics associated with
goal-tracking on the reward devaluation day compared to a baseline day. However, there was no significant difference in the responses of AIE-exposed rats and control-exposed rats. This finding is not consistent with a previous study, which found that female AIE-exposed rats showed
decreased sensitivity to reward devaluation compared to controls (Barker et al., 2017). This inconsistency could be explained by various differences in the methodology of the experiments.
Both experiments allowed ad libitum access to the reward solution for one hour prior to the behavioral session. However, in the study by Barker and colleagues, the behavioral session following reward devaluation took place in a novel cage while ours took place in the same
Finally, the subset of rats included in the devaluation task for our experiment was fairly small, which contributed to a high degree of variability. As a result, it was difficult to observe any
significant differences between exposure groups.
After establishing behavioral differences between AIE-exposed rats and controls, we applied in vivo optogenetics to evaluate whether activation of the orbito-accumbens pathway
could successfully reverse the effects of AIE exposure. Though results from an ex vivo
electrophysiology experiment confirmed that laser stimulation evoked neuronal firing of
ChR2-positive neurons in the OFC and NAcc, in vivo activation of this pathway failed to reverse the effects of AIE exposure or increase goal-tracking behavior. One possible explanation for this result is that the neuronal firing pattern evoked by laser stimulation was not physiologically
relevant. We selected our laser stimulation pattern and frequency to match previous findings that OFC neurons fire at 20 Hz (Stringfield et al., 2017). However, it is difficult to perfectly mimic
neuronal firing in vivo. Finally, one reason for lack of results in an in vivo optogenetics
experiment could be insufficient laser power at the tip of the optic fiber. We carefully measured the power output at the tip of the optic fiber prior to each laser stimulation session to ensure that
ChR2-positive neurons were effectively activated. Therefore, it is unlikely that insufficient laser power contributed to our lack of significant results.
Interestingly, laser stimulation only caused a significant behavioral change in rats that received the inactive virus. The inactive virus group exhibited a significant decrease in receptacle elevation score with laser stimulation, while rats that received the active virus did not show any
significant change in receptacle elevation score as a result of laser stimulation. The behavioral difference observed in rats with the inactive virus cannot be explained by stimulation of the
that the laser stimulation itself caused behavioral differences in these rats. For instance, a small amount of light is visible to the rat during the laser stimulation session, which could act as a
source of distraction. Because active virus rats were subject to this same potential distraction, it is possible that activation of the orbito-accumbens pathway reversed some of these unintended behavioral effects in the active virus group. This finding suggests that activation of the
orbito-accumbens pathway may counteract the effects of laser stimulation that led to reduced goal-tracking behavior in the inactive virus group. However, this sample size is fairly small, and more
data is needed to confirm this theory.
In the reward omission task, AIE-exposed rats had a less negative change score than control-exposed animals, regardless of whether they received the active or inactive virus. These
results were consistent with a previous study’s findings that AIE-exposed rats are less sensitive to reward omission than controls as a result of decreased behavioral flexibility (Gass et al.,
2014). There was no main effect of virus on receptacle elevation change score, which indicates that optogenetic activation of the orbito-accumbens pathway did not increase behavioral flexibility in exposed or control-exposed rats that received the active virus. In fact,
AIE-exposed rats that received the active virus appear to show decreased behavioral flexibility as a result of laser stimulation. However, there is a high degree of variability within this group, and
more data is needed to establish this conclusion. Overall, findings from the reward omission task indicate that activation of the orbito-accumbens pathway does not increase behavioral flexibility. This finding further supports the conclusion that optogenetic activation of the orbito-accumbens
pathway does not reverse the effects of AIE exposure.
In conclusion, in vivo optogenetic activation of the orbito-accumbens pathway is likely
Activation of this pathway did not reduce behavioral differences between AIE-exposed and control rats neither in a standard behavioral session nor in a reward omission session. However,
the behavior observed in this experiment is subject to a high degree of variability, and the sample size of rats with usable behavioral data from optogenetic activation is fairly small. We used rigorous standards in determining which rats could be included based on histology results
showing viral expression and optic fiber placement. Many rats were excluded from the optogenetics portion of the study due to insufficient viral expression or incorrect optic fiber
placement, as we were not confident that the orbito-accumbens pathway was selectively activated in these rats. Despite this small sample size, statistical analysis of data did not reveal any trends that suggested the predicted behavioral effect of increased goal-tracking or decreased
sign-tracking behavior as a result of activation of the orbito-accumbens pathway. Though results were not consistent with our hypothesis, this data is useful in exploring other potential neural
pathways that are affected by AIE exposure. For example, future studies can target the pathway between the orbitofrontal cortex and the dorsomedial striatum. Similar to the nucleus accumbens, this structure is involved in the brain’s reward system and has been found to have a role in
goal-directed reward learning and behavioral flexibility (Cole and Stone, 2017).
Optogenetics is a powerful, precise tool for manipulation of neuronal pathways. The
ability to selectively activate or inhibit neuronal networks in vivo has the potential to provide valuable information about neuronal networks that may play a role in addiction. In future studies, there is potential for the use of optogenetics to identify a pathway that, when manipulated, can
reverse the effects of binge alcohol exposure that are involved in vulnerability to addiction. By using optogenetic techniques to elucidate the role of specific neuronal circuits, more effective
addiction, as well as a wide range of other neurological disorders. Considering the high prevalence of alcohol consumption among adolescents and the damaging effects of
substance-use disorder and addiction, further research is necessary to explore additional methods of reversing the effects of binge-level alcohol exposure during adolescence.
ACKNOWLEDGEMENTS
This project was based on an initial hypothesis formulated by Dr. Aric Madayag and Dr.
Donita Robinson. Thank you to Dr. Alex Gomez-A, Dr. Carol Dannenhoffer, and Jose Pochapski for their valuable contributions to animal surgeries, histology, and behavioral training. In
addition, I would like to thank Nancy Sey and Hannah Jaggers for their help in collecting
behavioral data. Finally, thank you to all other members of the Robinson Lab for their support throughout the duration of this project!
REFERENCES
Barker, JM., Bryant, KG., et al (2017). Age and Sex Interact to Mediate the Effects of
Intermittent, High-Dose Ethanol Exposure on Behavioral Flexibility. Frontiers in Pharmacology, (8), pp. 450.
Broadwater, M., Lee, S., et al. (2017). Adolescent alcohol exposure decreases frontostriatal resting-state functional connectivity in adulthood. Addiction Biology (10.111).
Burguiere, E., Monteiro, P., et al. (2013). Optogenetic stimulation of lateral orbitofronto-striatal pathway suppresses compulsive behaviors. Science, 340 (6137), pp. 1243-6.
Gass, JT., Glen, WB., et al. (2014). Adolescent alcohol exposure reduces behavioral flexibility, promotes disinhibition, and increases resistance to extinction of ethanol self-administration in adulthood. Neuropsychopharmacology, 39 (11), pp. 2570-83.
Guru, A., Post, R., et al. (2015). Making Sense of Optogenetics. International Journal of Neuropsychopharmacology. 18(11).
Liu, W., Crews, FT. (2015). Adolescent intermittent ethanol exposure enhances ethanol
activation of the nucleus accumbens while blunting the prefrontal cortex responses in adult rat. Neuroscience, 293, pp. 92-108.
Madayag, A., Stringfield, S., et al. (2017). Sex and Adolescent Ethanol Exposure Influence Pavlovian Conditioned Approach. Alcoholism: Clinical and Experimental Research, 41(4), pp. 846-856.
McClory, A., Spear, L. (2014). Effects of ethanol exposure during adolescence or in adulthood on Pavlovian conditioned approach in Sprague-Dawley rats. Alcohol, 48(8), pp. 755-63. Morrison, SE., Bamkole, MA., et al (2015). Sign Tracking, but Not Goal Tracking, is Resistant to Outcome Devaluation. Frontiers in Neuroscience (9), pp. 468.
Stringfield, S., Palmatier, M., et al. (2017). Orbitofrontal participation in sign- and goal-tracking conditioned responses: effects of nicotine. Neuropharmacology, 116, pp. 208-223.