DOI: 10.1534/genetics.107.076885
Expression of 10 S-Class
SLF-like
Genes in
Nicotiana alata
Pollen and Its
Implications for Understanding the Pollen Factor of the S Locus
David Wheeler and Ed Newbigin
1Plant Cell Biology Research Centre, School of Botany, University of Melbourne, Parkville, Victoria 3010, Australia Manuscript received May 31, 2007
Accepted for publication October 3, 2007
ABSTRACT
The S locus ofNicotiana alataencodes a polymorphic series of ribonucleases (S-RNases) that determine the self-incompatibility (SI) phenotype of the style. The pollen product of the S locus (pollen S) inN. alatais unknown, but in species from the related genus Petunia and in self-incompatible members of the Plantaginaceae and Rosaceae, this function has been assigned to an F-box protein known as SLF or SFB. Here we describe the identification of 10 genes (designatedDD1–10) encoding SLF-related proteins that are expressed inN. alatapollen. Because our approach to cloning theDDgenes was based on sequences of SLFs from other species, we presume that one of theDDgenes encodes theN. alata SLFortholog. Seven of the DDgenes were exclusively expressed in pollen and a low level of sequence variation was found in alleles of eachDDgene. Mapping studies confirmed that all 10DDgenes were linked to the S locus and that at least three were located in the same chromosomal segment as pollen S. Finally, the different topologies of the phylogenetic trees produced using available SLF-related sequences and those produced usingS-RNasesequences suggests thatpollen Sand theS-RNasehave different evolutionary histories.
S
ELF-INCOMPATIBILITY (SI) is a genetic mecha-nism found in many flowering plants that acts to reduce the negative fitness effects associated with inbreed-ing (deNettancourt2001). In the best-understood ex-amples, SI is controlled by a single genetic S locus with many S alleles: styles reject pollen grains when both express the same S specificity. Although SI systems have evolved independently several times during the diver-sification of flowering plants (Mattonet al. 1994), in three families—the Solanaceae, Plantaginaceae, and Rosaceae—the SI phenotype of the style (but not of pollen) is controlled by an extracellular ribonuclease known as the S-RNase (Andersonet al. 1986; McClureet al. 1989; Sassaet al. 1993; Xueet al. 1996), a feature that suggests a single evolutionary origin for RNase-based SI systems (Igicand Kohn2001). Mechanistically, S-RNases are thought to act as cytotoxins that can enter both compatible and incompatible pollen tubes (Luu
et al. 2000; Goldraijet al. 2006), retarding the growth of incompatible pollen tubes possibly by degrading the limited amount of ribosomal RNA that they contain (McClureet al. 1990).
The S-locus pollen factor, known as pollen S, until recently was unknown. The first indication of its identity came from Lai et al. (2002), who sequenced a 63-kb
region ofAntirrhinum hispanicum (Plantaginaceae) ge-nomic DNA that contained theS-RNasegene and who found within this sequence an F-box gene specifically expressed in pollen.F-boxgenes were soon identified at the S loci of Petunia (Solanaceae) (Qiaoet al. 2004a; Y. Wanget al. 2004; Tsukamotoet al. 2005), Prunus, Pyrus, and Malus (Rosaceae) (Entani et al. 2003; Ushijima
et al. 2003; Sassaet al. 2007), and collectively these genes are now called theS locus F-boxgenes (abbreviated as SLF for the Solanaceae and Plantaginaceae and as SFB for the Rosaceae). Functionally, F-box proteins are a com-ponent of one type of E3 ubiquitin (Ub)-protein ligase, the enzyme responsible for transferring Ub from the E2 Ub-conjugating enzyme to free Lys residues on a selected protein target (Vierstra 2003). This associa-tion with protein ubiquitinaassocia-tion has led to a model in which compatible pollen tubes are protected from S-RNase cytotoxicity by a SLF-mediated mechanism in-volving Ub attachment and degradation (Laiet al. 2002; Qiaoet al. 2004b).
In each family, evidence exists to show thatSLF/SFB
encodes pollen S. In the Rosaceae, for example, where producing transgenic plants is difficult, statistical tests of amino acid variation between different Prunus SFB
alleles have identified two short regions of sequence that are under balancing selection (Ikeda et al. 2004; Nuneset al. 2006). SI is one of the best-documented examples of balancing selection and the way in which it affects nucleotide sequence variation has previously been used to identify genes involved in SI (Takebayashi
et al. 2003). Also consistent with a role for SFB in SI are Sequence data from this article have been deposited with the EMBL/
GenBank Data Libraries under accession nos. EF420251–EF420260. 1Corresponding author:Plant Cell Biology Research Centre, School of Botany, University of Melbourne, Parkville, VIC 3010, Australia. E-mail: [email protected]
truncations or deletions of this gene in self-compatible cultivars of fruit trees such as Prunus avium (sweet cherry) andPrunus mume( Japanese apricot) that carry mutations specifically affecting the SI phenotype of pollen (Ushijimaet al. 2004; Sonneveldet al. 2005). For the Solanaceae and Plantaginaceae, transgenic experi-ments have been used to show that SLF encodes a protein that behaves in the manner expected ofpollen S
(Qiaoet al. 2004a; Sijacicet al. 2004). ASLFtransgene does not cause pollen from a transgenic plant to express a new S-allele identity, but instead causes a breakdown of SI in pollen if theSLFtransgene is derived from an S allele that differs from the plant’s own S alleles (Qiao
et al. 2004a; Sijacicet al. 2004). Thus, theSLFtransgene derived from thePetunia inflataS2allele (PiSLF2) caused
loss of SI expression in pollen carrying either the S1or
S3allele but not in pollen carrying the S2allele (Sijacic
et al. 2004). The ability of two different pollen Salleles present in the same pollen grain to ‘‘competitively interact’’ with each other, and thus allow the pollen tube to grow through an otherwise incompatible style, is well known, having previously been observed in tetra-ploids derived from self-incompatible ditetra-ploids (e.g., Chawlaet al. 1997). SI plants with pollen-part muta-tions (pollen-part mutants, PPMs) are another example of competitive interaction between pollen S alleles. PPMs are diploid plants with an extra copy of the S locus that is present either as a small additional chro-mosome called a centric fragment, as a translocated fragment attached to a nonhomologous (i.e., non-S-bearing) chromosome, or as a fragment attached to a homologous (S-bearing) chromosome via unequal ex-change (Golzet al. 1999, 2001).
Genes related toSLF/SFBbut with no known role in SI also occur in plant genomes, including those of self-compatible plants lacking an RNase-based SI mecha-nism (L. Wang et al. 2004). These genes, referred to here asSLF-likegenes, are members of a large family of
F-box proteingenes found only in plants (L. Wanget al. 2004). Except forSLF/SFB, functions for most members of this family are unknown, even though it includes
13% of theF-box proteingenes (92/694) inArabidopsis thaliana (L. Wang et al. 2004). Phylogenetic analysis divides theSLF-likegene family into five different classes (A, B, C, M, and S), with allSLF/SFBgenes and some SLF-likegenes belonging to the S class (L. Wanget al. 2004). The presence of SLF-like genes can complicate the process of identifyingSLF/SFB genes, especially when
SLF-likegenes are pollen expressed and linked to the S locus (Entaniet al. 2003; Ushijimaet al. 2003; Zhou
et al. 2003; Y. Wanget al. 2004; Sassaet al. 2007). In this article we describe an analysis of the pollen-expressed S-class SLF-like genes from the solanaceous self-incompatible plantNicotiana alata. A motivation for this study was to broaden the range of solanaceous plants for which information onSLFgenes is available, as all existing sequences are from Petunia (Qiao et al.
2004a; Sijacicet al. 2004; Tsukamotoet al. 2005). Using degenerate primers and a reverse transcriptase PCR (RT–PCR) approach with N. alata pollen RNA as the template, full-length sequences for 10 S-class genes were obtained. Because potentially 1 of these 10 genes is the
N. alata SLF ortholog, we gave them the temporary namesDD1–10. Of the 10DDgenes, 7 were exclusively expressed in pollen. Polymorphisms in 7DDgenes were used to show linkage to the S locus: placement of individual DD genes with regard to the S-RNase gene and surrounding S-locus markers was done using the PPM plants described in Golz et al. (2001). Finally, phylogenetic analysis using DD and other S-class se-quences suggests that the SLFs in the Solanaceae and Plantaginaceae do not have a long history of co-evolving with theS-RNasegene. We discuss the implications that this has for identifyingpollen Sin these families and for understanding the evolutionary history of the pollen and stylar genes of the S locus.
MATERIALS AND METHODS
Plant material:SI lines ofN. alatahomozygous for S1, S2, S3, S6, or S7 were maintained as described in Anderson et al. (1986). The collection ofN. alataPPMs homozygous for the S6 allele and with mapped duplications containing the S3allele have been described previously (Golz et al. 1999, 2001). N. alatafamilies segregating for four S alleles were produced by germinating S1S3 3 S6S7 seed and S2S3 3 S6S7 seed. S genotypes were determined by PCR amplification of S-RNase genes as described by Liet al. (2000).
Primer design and PCR: The primers D2 59-YTIATIGGIC CITGYRAYGG-39(forward) and D4 59-CICCRTAYTSIWTCAT NAYCC-39 (reverse) were designed on the basis of regions conserved inSLFgenes from the Solanaceae and Plantagina-ceae. PCR was performed with50 ng of genomic DNA or 1ml of cDNA template. Reactions were carried out in a final volume of 20 ml of 13 PCR buffer (Invitrogen, San Diego) containing template DNA, 0.2–0.5mmof each primer, 0.2 mm dNTPs, 1.5 mm MgCl2, and 2 units of Taq polymerase (Invitrogen) on a GeneAmp 2700 thermal cycler (Applied Biosystems, Foster City, CA). Cycling conditions were as follows: 94° for 2 min and then 32 cycles of 94°at 30 sec, 45°–65°at 30 sec, and 72°at 60 sec. Rapid amplification of cDNA ends (RACE) PCR was performed using the Smart RACE kit (CLONTECH), as described in the manufacturer’s instructions.
generated using TRANALIGN (http://bioweb.pasteur.fr/docs/ EMBOSS/tranalign), based on ClustalW (v1.8) (Higgins and Sharp1988) protein alignments of the corresponding sequences. Maximum-parsimony (MP) trees were estimated from the DNA alignment using DNAPARS with default settings. Neighbor-joining (Saitouand Nei1987) distance trees were generated using NEIGHBOR, with a DNA dis-tance matrix computed using DNADIST. Bootstrap analyses (Felsenstein1985) were carried out using 1000 bootstrap pseudoreplicates.
DNA blot analysis:Genomic DNA was extracted from leaf material using the plant DNAeasy kit (QIAGEN). Leaf DNA (10mg) was digested to completion withEcoRI (Promega), fractionated on an agarose gel, and transferred to Hybond N1 (Amersham, Piscataway, NJ) membrane using the alkaline blot procedure described by the manufacturer. DNA fragments were radiolabeled with ½32PdCTP using the Primagene kit
(Promega). Hybridizations were performed overnight at 65°
in 53SSPE, 53Denhardt’s solution, 0.5% SDS, and 100mg/ ml denatured herring sperm. After hybridization membranes were washed twice in 23SSC and 0.1% SDS and exposed to film.
RT–PCR analysis: Total RNA was prepared using Trizol reagent (Invitrogen). DNA contamination was removed by treating isolated RNA (1 mg) with 2 units of DNase I (Invitrogen). First-strand cDNA synthesis was carried out using an oligo(dT17) primer and 200 units of Superscript III (Invitrogen). Sequences of the gene-specific primers used in this experiment are shown in supplemental Table 1 at http:// www.genetics.org/supplemental/.
RESULTS
Cloning and sequence analysis of candidate S-class SLF-like protein genes from N. alata pollen: Partial cDNAs for S-classSLF-likegenes were isolated from N. alatapollen by a degenerate PCR approach. Primers D2 and D4 were designed to bind to sequences conserved in Antirrhinum and PetuniaSLFs. Pollen RNAs isolated fromN. alataplants homozygous for the S1, S2, S3, S6, or
S7allele were reverse transcribed and used as templates.
PCR products were cloned and sequenced, and concep-tual translations of all six frames were compared to GenBank using BlastP. After several rounds of screen-ing, 10 partial cDNAs with high amino acid sequence similarity to Petunia SLF and SLF-like sequences were identified and given the provisional namesDD1–DD10. Sequence differences between the 10 partial cDNAs were 30–40% and there was #5% difference in the sequence of the sameDDcDNA amplified from differ-ent pollen RNA templates (data not shown). As theDD
cDNAs represented candidates forN. alata SLF, RACE was used to obtain additional 59 and 39 sequence information. Full-length sequences forDD1and DD5–
8were obtained by RACE using S2S2pollen cDNA as the
template. Full-length sequences forDD2 (S6S6), DD3/
DD4 (S1S1), and DD9 (S3S3) were obtained from the
indicated pollen cDNAs, and a near full-length se-quence forDD10(missing the initiator Met codon and 59-UTR) was obtained from S6S6pollen cDNA.
Figure 1 shows conceptual translations of the 10DD
cDNAs aligned with representative SLF and SLF-like
proteins from P. inflata (Solanaceae), A. hispanicum
(Plantaginaceae) and P. mume (Rosaceae). DD5 and DD8 were the most closely related of the N. alataDD proteins with 88% amino acid similarity (78% identity), and DD3 and DD7 were the least related with 50% amino acid similarity (35% identity). The protein similarity between the DD sequences and P. inflata
SLF-S3ranged from 53% (DD7) to 80% (DD3). SMART
and PFAM analysis identified an N-terminal F-box do-main in allN. alataDD proteins, consistent with them being members of the F-box protein superfamily. Other F-box proteins have diverse regions outside the F-box domain that bind to specific substrate proteins (Gagne
et al. 2002). Potential substrate-binding motifs present in the DD proteins include the related F-box-associated domains type 1 and type 3 (FBA_1, PF07734; FBA_3, PF08268) (Figure 1). L. Wanget al. (2004) incorporated these domains into the four conserved C-terminal motifs (C1–C4) used to define the S-class SLF-like proteins. C1–C4 are present in the SLF/SFB proteins and in all 10 DD proteins (Figure 1).
Nucleotide sequences of the N. alata DDgenes were aligned with sequences of the SLF and S-class SLF-like
genes fromP. inflata,Petunia axillaris, andPetunia hybrida
(Solanaceae) and A. hispanicum (Plantaginaceae). Se-quences ofSFBs from species in the Rosaceae were also included. Figure 2 shows the MP tree produced from the DNA alignment. Bootstrap analysis provides strong support for almost all nodes, and distance trees gen-erated using the same alignment had an identical to-pology (results not shown). The Solanaceae SLF and
SLF-like sequences are in a single cluster, as are the Antirrhinum sequences, with nodes at the base of the Solanaceae and Plantaginaceae clusters (nodes 1 and 2) having strong bootstrap support (100%).DD7andDD6
are at the base of the Solanaceae cluster and sisters to three well-supported smaller clusters (nodes 3–5). The
P. inflata SLFsequences,PiSLF-S1,PiSLF-S2, andPiSLF-S3,
group together in a cluster (node 6) withPaF1, an SLF-likesequence fromP. axillaris(Tsukamotoet al. 2005). These sequences are sisters to the Nicotiana sequences
DD1,DD3, andDD4(node 5). The second small cluster containsP. hybridaandP. inflata SLF-likesequences and
DD2 (node 4), and the third cluster comprises only
DDsequences (node 3).
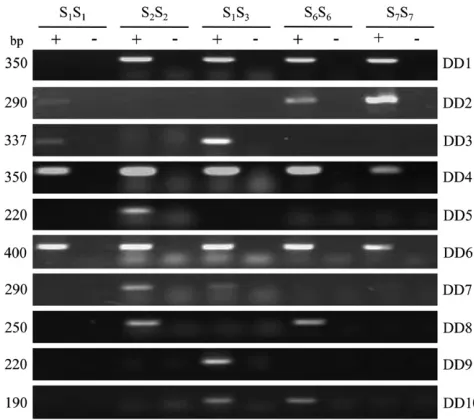
Expression of the DD genes in various N. alata S genotypes and tissues: To determine whether expres-sion of aDDgene was restricted to pollen of a particular
S genotype, PCR experiments were done with gene-specific primers and S1S1, S2S2, S1S3, S6S6, and S7S7
five pollen templates tested (DD4 andDD6) and tran-scripts for another 6DD genes (DD1–DD3, DD7,DD8, andDD10) were amplified from between two and four templates. For example,DD1transcripts were present in all pollen cDNAs except S1S1, andDD2transcripts were
detectable in S1S1, S6S6, and S7S7cDNAs, but not in S2S2
and S1S3 cDNAs. DD3 was detected in S1S1 and S1S3
pollen cDNAs but could not be amplified from S3S3
cDNA (data not shown). The remaining twoDDgenes were more restricted in their expression and transcripts were detectable in only a single cDNA type: the S2S2
template forDD5and the S1S3template forDD9.
The tissue-specific expression patterns of the DD
genes were investigated by RT–PCR using cDNA synthe-sized from N. alata pollen, style, leaf, and petal RNA. Figure 4 shows that DD2, DD3, and DD5–9 were ex-pressed only in pollen, that DD1 and DD4 were ex-pressed in pollen and style, and thatDD10was expressed in pollen and petal. TheDDgenes were not expressed at detectable levels in nonfloral tissues such as leaf or root (Figure 4 and data not shown).
Determining linkage between theDDgenes and theS locus: Evidence of linkage between the S locus and individualDDgenes was obtained using two families of
N. alataplants in which four S alleles were segregating.S
genotypes of progeny plants resulting from a S1S3 3
S6S7cross or a S2S33S6S7cross were determined using
allele-specific PCR of the S-RNase gene (results not shown). Cleavable amplified polymorphic sequence markers were designed for eight of the N. alata DD
genes to test for cosegregation with an individual S-RNaseallele (e.g.,S3). Analysis of 18–20 plants detected no
recombination between the DD marker and S3-RNase
(supplemental Table 2 at http://www.genetics.org/ supplemental/), indicating that theseDDgenes are,5 cM from theSlocus.
Mapping of the DD genes was also done using a collection of PPMN. alataplants that are homozygous for the S6 allele and that carry chromosomal
duplica-Figure 2.—Maximum-parsimony tree (5087 steps) derived from aligned SLF/SFB and SLF-like DNA sequences. The three Petunia sequences identified with pollen Sare indicated by boldface type. Bootstrap values (1000 pseudoreplicates) for numbered nodes are 1 ¼ 100%, 2 ¼ 100%, 3¼96%, 4¼100%, 5¼99%, 6¼100%. Sequen-ces in the tree and their acSequen-cession numbers are PaSLFS17 (AY766153), PaSLFS19 (AY766154), PhSLFS3A (AY639403), PhSLFS3B (AY639402), PaF1 (AY766155), PiS3A113 (AY363972), S1A134 (AY363973), PiS2A134 (AY363974), Pi-S2A113 (AY363971), AhSLFS5 (AJ515536), AhSLF-S1E (AJ515535), AhSLF-S4D (AJ515534), AhSLFS4A (AJ515534), PmSFBS7 (AB092622), PdSFBa (AB092966), MdSFBB9b (AB270794), MdSFBB9a (AB270793), PpFBB4a (AB270797), PpSFBB4b (AB270798), PavSFB1 (PavSFB1), and PbSFBb (AB081648).
tions of varying lengths containing anS3 allele (dS3).
The duplications are present in the genomes of these plants as small extra chromosomes (centric fragments), segments that have been added to other chromosomes (translocations), or segments that are linked to anS6
allele via an unequal crossover (Figure 5B). This ap-proach to mapping genes to theN. alata Slocus is quicker than the standard linkage-based approach (supplemental Table 2 at http://www.genetics.org/supplemental/) and provides information about relative gene order, but is suitable only for genes withS3-andS6-associated
poly-morphisms. Suitable polymorphisms were detected for
DD1–3, DD6, DD7,DD9, andDD10. DNA blot analysis indicated that the DD genes were all single-copy se-quences in theN. alatagenome (Figure 5A and data not shown).
Figure 5 illustrates how the PPMs were used to determine map locations for theDDgenes. Four PPM lines were selected for this analysis (M1-1, M1-2, M1-5, and M1-7) with Figure 5B showing the previously de-scribed order of markers (CP100, 48A, S-RNase, and
167A) on dS3in each line.CP100,48A, andS-RNaseall
map without recombination to theSlocus and167Ais 0.9 cM away (Liet al. 2000). On blots of genomic DNA fromS3S3,S6S6, M1-1, M1-2, M1-5, and M1-7 (Figure 5A),
the DD1 probe detected a restriction fragment length polymorphism betweenS3S3(DD1–3) andS6S6(DD1–6).
As expected, DD1–6 was present in all M1 plants (S6S6dS3) butDD1–3was present only in M1-7, placing
DD1to the right of marker167A(Figure 5C). Similarly, theDD2probe hybridized toDD2–3andDD2–6inS3S3
andS6S6, respectively (Figure 5A).DD2–3was present in
all M1 plants, placing this gene within theSlocus in a chromosomal segment delimited by the left border of the translocation in M1-7 and the right border of the unequal exchange in M1-5 (Figure 5C). The other genes known to lie in this segment are48Aandpollen S. Figure 5C shows DD2 grouped with 48A on the centromeric side of the S locus, although the placement of these markers with respect to each other andpollen S
is arbitrary. Figure 5C summarizes the PPM mapping data. DD7 and DD10 are in the same chromosomal segment asDD2,DD9lies between theS-RNaseand167A, andDD1,DD3, andDD6are farther from the S locus than
167A (supplemental Figure 6 at http://www.genetics. org/supplemental/).
DISCUSSION
This article describes the identification and initial characterization of 10 genes expressed inN. alatapollen (DD1–10) that code for S-class SLF-like proteins, a class that includes all the known SLFs of Antirrhinum and Petunia, the SFBs of Prunus, and other clearly related proteins in these and other species with no obvious role in SI (L. Wang et al. 2004). The DD genes were all present as single-copy sequences and the 10DDgenes appear to represent the full complement of S-class S
SLF-likegenes expressed inN. alatapollen that can be recovered using the RT–PCR-based approach described here. Because none of theDDgenes could unambigu-ously be identified as encoding theN. alataortholog of SLF (see below), we propose to continue using this provisional naming scheme until such time as functions are identified and individual DD genes can be given more descriptive names.
A surprising aspect of this work was the linkage detected in small-scale experiments between 8 of the 10DDgenes (DD3–DD10) and theSlocus (supplemen-tal Table 2 at http://www.genetics.org/supplemen(supplemen-tal/). This potentially indicated the existence of a single cluster ofDDgenes. Clusters ofF-box proteingenes have been described in Arabidopsis by Gagneet al. (2002), where 35.9% of the 694F-box proteingenes are in arrays of two to seven members. Instead of a single cluster of
DDgenes, it could equally be true that there are many separate SLF-like genes, all subject to the suppressed recombination affecting the S locus and precluding the
production of well-resolved genetic maps (Liet al. 2000; Wanget al. 2003). The relative order of seven of theDD genes with respect to each other and the surrounding S locus markers, however, could be determined using a small number of PPMs with duplicated S3 allele
seg-ments of differing sizes (Golzet al. 2001). This analysis showed that fourDDgenes (DD2,DD7,DD9, andDD10) exist as single genes (or possibly as a cluster of a few genes) on either side of theS-RNasegene and within the S locus. Three of the DDgenes (DD1, DD3, and DD6) were farther from the S-RNase gene than the marker
167Aand accordingly must be at least 0.9 cM from the S locus. In total, all 10DDgenes were linked to the S locus, although a lack of suitable polymorphisms for DD4,
DD5, and DD8 meant that the position of these genes relative to other S-locus markers and each other could not be determined.
Even though all the class SSLF-likegenes identified in this study were expressed inN. alatapollen, they clearly cannot all be involved in SI. Some, for example, were expressed in tissues other than pollen—specificallyDD1
andDD3, which were also expressed in styles, andDD10, which had some expression in petals. Since all SLFs
characterized to date are expressed only in pollen, these genes can be excluded from consideration as potential
SLForthologs. The remaining sevenDDgenes, however, appear to be expressed only in pollen and thus warrant further study. Although all theDDgenes are at or near the N. alataS locus, fine-scale mapping indicates that only three—DD2, DD7, and DD10—are in the same region of the chromosome aspollen S. Since expression in petal means thatDD10can be excluded, onlyDD2and
DD7can still be considered possibleSLForthologs, with the status ofDD4,DD5, andDD8being uncertain.
Interestingly, of these five ‘‘candidate’’ genes, DD5
transcripts were amplified only from S2S2pollen cDNA,
andDD2,DD7, andDD8transcripts were amplified only from two of the five pollen cDNA templates tested (Figure 3 and data not shown). DD4 transcripts were amplified from all five pollen templates. While this could potentially reflect a level of S-allele specificity, amplification of transcripts from some cDNA templates but not others was also seen with DD genes that are clearly not pollen S (e.g., DD9 was amplified only from S1S3pollen cDNA).
Both the number of S-classSLF-likegenes at or near the S locus and their pollen expression are worthy of comment. No such clustering of S-classSLF-likegenes is apparent in Arabidopsis, where the 7 genes of this type are distributed across three of the five chromosomes (L. Wanget al. 2004). Predominant or exclusive expression of 10 S-class SLF-like genes in N. alata pollen also contrasts with the situation in Arabidopsis, where six of the seven genes are constitutively expressed and only one,AtSFL79, is strictly pollen expressed (L. Wanget al. 2004). SLF-likegenes, however, have been found at or near the S locus in the Rosaceae (e.g., see Entaniet al. 2003; Ushijima et al. 2003; Sassa et al. 2007), the Plantaginaceae (Zhou et al. 2003), and in the solana-ceous speciesP. inflata(Wanget al. 2003). In each case, these S-locus-linked F-box protein genes are predomi-nantly or exclusively expressed in pollen, just like theDD
genes. We speculate that the number of S-classSLF-like
genes at the S locus and their pollen expression is not due to chance but is an evolutionary outcome arising from the state of permanent heterozygosity that SI imposes.
In a process analogous to that leading to a loss of active genes on the male-determining Y chromosome of
Figure5.—Linkage of theDDgenes to the S locus. (A) Representative Southern blots using probes specific for the indicated DDgene. The source of genomic DNA is shown at the top of each lane and the different alleles are indicated at the right. (B) Maps ofdS3in each of the
indi-cated PPMs. A solid dot indicates the centromere. All PPMs used in this experiment have the geno-typeS6S6dS3. (C) Map of theN. alataS locus
animals (see Skaletskyet al. 2003), permanent hetero-zygosity at the S locus will reduce the effectiveness of selection against deleterious mutations occurring in genes with unrelated functions that are embedded within it (Uyenoyama1997, 2005). As each individual S allele experiences a small effective population size, deleterious mutations in genes tightly linked to the S locus are highly likely to replace functional wild-type alleles, a process that should, over time, lead to the de-generation of most genes within the S-locus region except those with SI-specific functions. Any gene ex-pressed in a haploid cell like a pollen grain will escape this effect because there are no genes associated with a second S allele to provide intransthe functions lost by mutation. Hence these genes will still be subject to selec-tion against deleterious mutaselec-tions, especially if their products contribute to the reproductive fitness of pol-len. Selection may also favor expanding the number of these genes at the S locus through gene duplication in cases where higher expression levels enhance male fit-ness. Thus, this evolutionary scenario suggests that the cluster ofDDgenes at theN. alataS locus arises because enforced heterozygosity favors genes that contribute to male fitness and that through pollen expression avoid gene erosion processes.
The phylogenetic tree in Figure 2 highlights several puzzling aspects of the S-class SLF-like proteins. Most notably, the SLF/SFB proteins of the Plantaginaceae, Solanaceae, and Rosaceae do not form a monophyletic clade to the exclusion of other S-class SLF-like proteins with no role in SI, such as many of the DD proteins. Contrast can be made here to trees of S-RNases and the related S-like RNases, which are not involved in SI. Phylogenetic analysis places S-RNases and S-like RNases into one of three classes: a single monophyletic clade that contains all S-RNases from the Solanaceae, Plantaginaceae, and Rosaceae and two separate clades of S-like RNases (Igicand Kohn2001; Steinbachsand Holsinger 2002). The apparent homology of the S-RNases leads to the conclusion that the three known RNase-based SI systems share a single evolutionary ori-gin. In Figure 2, however, S-class SLF-like proteins are grouped not according to function but on the basis of the taxonomic relationships of the organisms from which they were derived. Thus the conclusion that the S-RNases of the Solanaceae, Rosaceae, and Plantagina-ceae are homologous is not borne out by this analysis of the SLFs and SFBs. Assuming that the SLFs and SFBs are involved in SI, then the most parsimonious interpreta-tion of Figure 2 is that their encoding genes have been independently recruited to roles in RNase-based SI in the Rosaceae, Plantaginaceae, and Solanaceae. Lack of homology implies that different mechanisms may be used to achieve the rejection of incompatible pollen in each family.
A second notable aspect of the phylogenetic recon-struction in Figure 2 is the well-resolved clade arising at
node 5 that contains all the Petunia SLFs, proteins such as PaF1 (P. axillarisF-box protein 1) that are presumed not to be at the Petunia S locus (Tsukamotoet al. 2005), and two DD proteins (DD1 and DD3) that are not considered candidates for the N. alata SLF ortholog because their encoding genes are at least 0.9 cM from the S locus (Figure 5). Numerous phylogenetic analyses of the solanaceous S-RNases place these sequences in a well-resolved monophyletic clade that excludes S-like RNases (e.g., Igic and Kohn 2001; Steinbachs and Holsinger2002). Any RNase sequence falling into the S-RNase clade not involved in self-incompatibility (e.g., one obtained from a self-compatible plant) is presumed to be derived from a nonfunctional S-RNase (relic S-RNases; Golzet al. 1998). For the clade in Figure 2 to be a clade of solanaceous SLFs resembling the clade of solana-ceous S-RNases, DD1, DD3, and PaF1 must all be relic SLFs. Since these proteins are from self-incompatible species, it therefore becomes problematic to assign the name SLF to any protein sequence within this clade unless its function in SI has been verified experimen-tally. It is also notable that four of the remaining five SLF candidates—DD2,DD5,DD7, andDD8—are not in this SLF-containing clade.
The final puzzling feature of the tree is the shortness of terminal branches for SLFs in the Plantaginaceae and Solanaceae, reflecting the low levels of sequence poly-morphism among SLF alleles in these two families. Notably, this feature is not seen with SFBs from the Rosaceae. The four Antirrhinum SLF alleles are identi-cal at 97% or more of their amino acid positions and the two most divergent Petunia SLF alleles, PiSLF-S3 and
PaSLF-S17, are 87% identical. These percentages of
identity values are closer to those of genes with functions unrelated to self-incompatibility that also lie within the S locus (such as48A; Takebayashiet al. 2003) than they are to the S-RNases, where pairwise amino acid identities of 40–50% are common (Clarkand Kao 1994). The pairwise amino acid identity in partial cDNAs of DD1, DD2, DD4, DD6, DD7, DD8, and DD10 from different S-allele backgrounds ranges from 1 to 5%.
SI imposes a very intense form of balancing selection on the S locus, as a consequence of which high numbers of S-allele lineages are maintained over long periods of time (reviewed by Clark and Kao 1994; Richman 2000). The effect that balancing selection has had on the phylogenetic history of the S-RNase gene, particu-larly on extending the phylogenies of allelic lineages over time spans that are vastly longer than those ex-pected under neutrality, has been demonstrated many times (see Ioergeret al. 1990; Richmanet al. 1996). In the Rosaceae, identifying SFBs as the pollen S factor is based in part on agreement between the expected phylogeny of a gene under balancing selection and the observed SFB phylogenies (Ikedaet al. 2004; Nunes
phylogeny did not show the expected features (e.g., Takebayashiet al. 2003).
Little evidence of this predicted evolutionary history is seen in the Petunia and Antirrhinum SLFs, however: the clades containing these sequences are marked by short terminal branches that quickly converged on the common ancestor of all sampled lineages. Because in the Rosaceae SFBs, sites identified as positively selected largely overlap with regions of highest sequence vari-ability (Ikedaet al. 2004; Nuneset al. 2006), we visually inspected an alignment of SLF proteins from the Solanaceae for highly variable sites (i.e., sites occupied by many different amino acids; see supplemental Figure 7 at http://www.genetics.org/supplemental/). As only one such site was found, it is probable that the sequences also lack positively selected sites likely to be responsible for defining pollen specificities.
The marked lack of concordance between S-RNase trees, which are largely consistent with well-established theories as to how genes evolve under balancing selection (Clarkand Kao1994), and the S-class SLF-like tree for the Solanaceae leads to the unsettling conclusion that the SLFs and S-RNases are not stably co-evolving partners, which previously had been assumed would be true of the pollen and stylar factors of the S locus. Two possible explanations of this situation exist: either there is no obligate relationship between the S-RNase and a particular S-class F-box protein or the SLFs do not fulfill the role of pollen S in the Solanaceae and Plantaginaceae.
Although phylogenetic reconstruction of the S-class SLF-like proteins provides little indication that the SLFs have a long history of evolving under balancing selec-tion, other evidence is consistent with the expected behavior of pollen S. Apart from their genetic location and expression pattern, this evidence includes the binding of recombinant SLF proteins to S-RNases and the pollination behavior of transgenic plants that express SLFs in their pollen (e.g., see Qiao et al. 2004b; Sijacicet al. 2004). It should, however, be noted that so far it has not been possible to demonstrate allele specificity for the SLFs (McClure2004). Recombinant SLF proteins bind S-RNases nonspecifically, and SLF expression in transgenic plants results in a loss of SI rather than in the gain of a new S-allele specificity. Since other proteins can bind S-RNases (Simsand Ordanic 2001), and since mutations in other genes can lead to self-compatible phenotypes (e.g., Ai et al. 1991), it re-mains formally possible that the properties currently used to define pollen S in the Solanaceae and Planta-ginaceae are not exclusive to this factor.
The alternative explanation is that the functions of
pollen Sare transient and able to be performed by any one of the many paralagous S-classSLF-likegenes within the S locus. Sassaet al. (2007) recently made a similar suggestion on the basis of their observation that the S locus in Malus and Pyrus (Rosaceae) contained two or
more copies of theSFBgene (‘‘SFB brothers’’ orSFBBs). If paralagous SLF genes act as proposed, then pollen S
lineages might be expected to turn over far more rapidly than S-RNase lineages. Although this suggestion cur-rently lacks experimental support, it is obvious that identifying which, if any, of the DD genes codes for pollen S inN. alatawill help refine ideas as to why the
SLFandS-RNasegenes have such remarkably divergent evolutionary histories.
We thank Bruce McGinness for assistance in the glass house and Poh Ling Koh for experimental help. This work is supported by a grant from the Australian Research Council.
LITERATURE CITED
Ai, Y. J., E. Kronand T. H. Kao, 1991 S-alleles are retained and
ex-pressed in a self-compatible cultivar ofPetunia hybrida. Mol. Gen. Genet.230:353–358.
Anderson, M. A., E. C. Cornish, S.-L. Mau, E. G. Williams, R. Hoggart
et al., 1986 Cloning of cDNA for a stylar glycoprotein associated with expression of self-incompatibility inNicotiana alata. Nature 321:38–44.
Chawla, B., R. Bernatzky, W. Liang and M. Marcotrigiano,
1997 Breakdown of self-incompatibility in tetraploid Lycopersi-con peruvianum: inheritance and expression of S-related proteins. Theor. Appl. Genet.95:992–996.
Clark, A. G., and T. Kao, 1994 Self-incompatibility: theoretical concepts
and evolution, pp. 220–242 inGenetic Control of Self-Incompatibility and Reproductive Development in Flowering Plants, edited by E. G. Williams, R. B. Knoxand A. E. Clarke. Kluwer, Dordrecht,
The Netherlands.
deNettancourt, D., 2001 Incompatibility and Incongruity in Wild and
Cultivated Plants. Springer, New York.
Entani, T., M. Iwano, H. Shiba, F.-S. Che, A. Isogai et al.,
2003 Comparative analysis of the self-incompatibility (S-) locus region ofPrunus mume: identification of a pollen-expressed F-box gene with allelic diversity. Genes Cells8:203–213.
Felsenstein, J., 1985 Confidence limits on phylogenies: an
ap-proach using the bootstrap. Evolution39:783–791.
Felsenstein, J., 2004 PHYLIP (Phylogeny Inference Package), Version
3.6. University of Washington, Seattle.
Gagne, J. M., B. P. Downes, S. H. Shiu, A. M. Durskiand R. D. Vierstra,
2002 The F-box subunit of the SCF E3 complex is encoded by a diverse superfamily of genes in Arabidopsis. Proc. Natl. Acad. Sci. USA99:11519–11524.
Goldraij, A., K. Kondo, C. B. Lee, C. N. Hancock, M. Sivaguru
et al., 2006 Compartmentalization of S-RNase and HT-B degra-dation in self-incompatibleNicotiana. Nature439:805–810. Golz, J. F., A. E. Clarke, E. Newbiginand M. Anderson, 1998 A
relic S-RNase is expressed in the styles of self-compatible Nicoti-ana sylvestris. Plant J.16:591–599.
Golz, J. F., V. Su, A. E. Clarkeand E. Newbigin, 1999 A molecular
description of mutations affecting the pollen component of the
Nicotiana alataS locus. Genetics152:1123–1135.
Golz, J. F., H. Y. Oh, V. Su, M. Kusaba and E. Newbigin,
2001 Genetic analysis ofNicotianapollen-part mutants is consis-tent with the presence of an S-ribonuclease inhibitor at the S lo-cus. Proc. Natl. Acad. Sci. USA98:15372–15376.
Higgins, D. G., and P. M. Sharp, 1988 CLUSTAL: a package for
per-forming multiple sequence alignment on a microcomputer. Gene73:237–244.
Igic, B., and J. R. Kohn, 2001 Evolutionary relationships among
self-incompatibility RNases. Proc. Natl. Acad. Sci. USA 98: 13167–13171.
Ikeda, K., B. Igic, K. Ushijima, H. Yamane, N. R. Hauck et al.,
2004 Primary structural features of the S haplotype-specific F-box protein, SFB, inPrunus. Sex. Plant Reprod.16:235–243. Ioerger, T. R., A. G. Clarkand T. H. Kao, 1990 Polymorphism at
Lai, Z., W. Ma, B. Han, L. Liang, Y. Zhanget al., 2002 An F-box
gene linked to the self-incompatibility (S) locus ofAntirrhinum
is expressed specifically in pollen and tapetum. Plant Mol. Biol. 50:29–42.
Letunic, I., R. R. Copley, B. Pils, S. Pinkert, J. Schultz et al.,
2006 SMART 5: domains in the context of genomes and net-works. Nucleic Acids Res.34:D257–D260.
Li, J.-H., N. Nass, M. Kusaba, P. N. Dodds, N. Treloaret al., 2000 A
genetic map of theNicotiana alataS locus that includes three pol-len-expressed genes. Theor. Appl. Genet.100:956–964. Luu, D. T., X. Qin, D. Morseand M. Cappadocia, 2000 S-RNase
uptake by compatible pollen tubes in gametophytic self-incom-patibility. Nature407:649–651.
Matton, D. P., N. Nass, A. E. Clarkeand E. Newbigin, 1994
Self-incompatibility: how plants avoid illegitimate offspring. Proc. Natl. Acad. Sci. USA91:1992–1997.
McClure, B., 2004 S-RNase and SLF determine S-haplotype-specific
pollen recognition and rejection. Plant Cell16:2840–2847. McClure, B. A., V. Haring, P. R. Ebert, M. A. Anderson, R. J. Simpson
et al., 1989 Style self-incompatibility gene products ofNicotiana alataare ribonucleases. Nature342:955–957.
McClure, B. A., J. E. Gray, M. A. Anderson and A. E. Clarke,
1990 Self-incompatibility in Nicotiana alata involves degrada-tion of pollen rRNA. Nature347:757–760.
Nunes, M. D. S., R. A. M. Santos, S. M. Ferreira, J. Vieiraand C. P.
Vieira, 2006 Variability patterns and positively selected sites at
the gametophytic self-incompatibility pollen SFB gene in a wild self-incompatible Prunus spinosa (Rosaceae) population. New Phytol.172:577–587.
Qiao, H., F. Wang, L. Zhao, J. Zhou, Z. Laiet al., 2004a The F-box
protein AhSLF-S2controls the pollen function of S-RNase-based
self-incompatibility. Plant Cell16:2307–2322.
Qiao, H., H. Wang, L. Zhao, J. Zhou, J. Huanget al., 2004b The
F-box protein AhSLF-S2physically interacts with S-RNases that may
be inhibited by the ubiquitin/26S proteasome pathway of pro-tein degradation during compatible pollination inAntirrhinum. Plant Cell16:582–595.
Richman, A., 2000 Evolution of balanced genetic polymorphism.
Mol. Ecol.9:1953–1963.
Richman, A. D., M. K. Uyenoyamaand J. R. Kohn, 1996 Allelic
di-versity and gene genealogy at the self-incompatibility locus in the Solanaceae. Science273:1212–1216.
Saitou, N., and M. Nei, 1987 The neighbor-joining method: a new
method for reconstructing phylogenetic trees. Mol. Biol. Evol.4: 406–425.
Sassa, H., H. Hiranoand H. Ikehashi, 1993 Identification and
charac-terization of stylar glycoproteins associated with self-incompatibility genes of Japanese pear,Pyrus serotinaRehd. Mol. Gen. Genet.241: 17–25.
Sassa, H., H. Kakui, M. Miyamoto, Y. Suzuki, T. Hanada et al.,
2007 S-locus box brothers: multiple and pollen-specific F-box genes with S haplotype-specific polymorphisms in apple and Japanese pear. Genetics175:1869–1881.
Sijacic, P., X. Wang, A. L. Skirpan, Y. Wang, P. E. Dowd et al.,
2004 Identification of the pollen determinant of S-RNase-mediated self-incompatibility. Nature429:302–305.
Sims, T. L., and M. Ordanic, 2001 Identification of a
S-ribonuclease-binding protein inPetunia hybrida. Plant Mol. Biol.47:771–783. Skaletsky, H., T. Kuroda-Kawaguchi, P. J. Minx, H. S. Cordum, L.
Hillieret al., 2003 The male-specific region of the human Y
chromosome is a mosaic of discrete sequence classes. Nature 423:825–837.
Sonneveld, T., K. R. Tobutt, S. P. Vaughanand T. P. Robbins,
2005 Loss of pollen-S function in two self-compatible selections ofPrunus aviumis associated with deletion/mutation of an S hap-lotype-specific F-box gene. Plant Cell17:37–51.
Steinbachs, J. E., and K. E. Holsinger, 2002 S-RNase-mediated
ga-metophytic self-incompatibility is ancestral in eudicots. Mol. Biol. Evol.19:825–829.
Takebayashi, N., P. B. Brewer, E. Newbiginand M. K. Uyenoyama,
2003 Patterns of variation within self-incompatibility loci. Mol. Biol. Evol.20:1778–1794.
Tsukamoto, T., T. Ando, H. Watanabe, E. Marchesiand T. H. Kao,
2005 Duplication of the S-locus F-box gene is associated with breakdown of pollen function in an S-haplotype identified in a natural population of self-incompatible Petunia axillaris. Plant Mol. Biol.57:141–153.
Ushijima, K., H. Sassa, A. M. Dandekar, T. M. Gradziel, R. Tao
et al., 2003 Structural and transcriptional analysis of the self-incompatibility locus of almond: identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. Plant Cell15:771–781.
Ushijima, K., H. Yamane, A. Watari, E. Kakehi, K. Ikedaet al.,
2004 The S haplotype-specific F-box protein gene, SFB, is de-fective in self-compatible haplotypes of Prunus avium and P. mume. Plant J.39:573–586.
Uyenoyama, M. K., 1997 Genealogical structure among alleles
reg-ulating self-incompatibility in natural populations of flowering plants. Genetics147:1389–1400.
Uyenoyama, M. K., 2005 Evolution under tight linkage to mating
type. New Phytol.165:63–70.
Vierstra, R. D., 2003 The ubiquitin/26S proteasome pathway, the
complex last chapter in the life of many plant proteins. Trends Plant Sci.8:135–142.
Wang, L., L. Dong, Y. Zhang, W. Wu, X. Denget al., 2004
Genome-wide analysis of S locus F-box-like genes inArabidopsis thaliana. Plant Mol. Biol.56:929–945.
Wang, Y., X. Wang, A. G. McCubbinand T. H. Kao, 2003 Genetic
mapping and molecular characterization of the self-incompatibility (S) locus inPetunia inflata. Plant Mol. Biol.53:565–580. Wang, Y., T. Tsukamoto, K. W. Yi, X. Wang, S. Huanget al., 2004
Chro-mosome walking in thePetunia inflataself-incompatibility (S-) locus and gene identification in an 881-kb contig containing S2-RNase.
Plant Mol. Biol.54:727–742.
Xue, Y., R. Carpenter, H. G. Dickinson and E. S. Coen,
1996 Origin of allelic diversity inAntirrhinumS locus RNases. Plant Cell8:805–814.
Zhou, J., F. Wang, W. Ma, Y. Zhang, B. Hanet al., 2003 Structural
and transcriptional analysis of S-locus F-box genes inAntirrhinum. Sex. Plant Reprod.16:165–177.