Fish red blood cells (RBCs) possess volume-sensitive passive permeation mechanisms for the inorganic ions and amino acids (AA) that are involved in cell volume regulation. Thus, swelling of RBCs in a hypotonic medium is succeeded by a regulatory volume decrease (RVD) response, in which stimulation of dissipative K+and AA pathways leads to a net osmolyte efflux from the cells. This is accompanied by an osmotically obligated water loss, which restores cell volume towards the original value (Lauf, 1982; Fincham et al. 1987;
Garcia-Romeu et al. 1991). In addition to the volume-activated K+ efflux, recent studies have demonstrated an oxygenation-sensitive K+/Cl2cotransport mechanism in the membrane of fish RBCs (Jensen, 1990, 1992; Borgese et al. 1991; Nielsen and Lykkeboe, 1992; Nielsen et al. 1992). This K+ efflux
mechanism is activated by oxygenation but also by methaemoglobin formation or CO binding to the haemoglobin and it is inactivated by deoxygenation and by low pH, indicating that it is influenced by haemoglobin conformation rather than oxygenation conditions per se (see Jensen, 1992). The oxygenation-activated K+/Cl2 cotransport elicits cell shrinkage in the absence of a prior volume perturbance and may be closely related to the volume-reducing KCl extrusion seen after cell swelling. It is accordingly of interest to study the interactions between oxygenation-activated and volume-activated K+pathways in fish RBCs and to examine whether the K+ and AA permeation mechanisms used in volume regulation are dependent on the degree of RBC oxygenation. On the basis of the pH sensitivity of the oxygenation-activated
155
The Journal of Experimental Biology198, 155–165 (1995)
Printed in Great Britain © The Company of Biologists Limited 1995
Hypo-osmotic swelling of carp red blood cells (RBCs) induced a regulatory volume decrease (RVD), which restored the original cell volume within 140 min in oxygenated RBCs, whereas volume recovery was incomplete in deoxygenated RBCs. The complete RVD in oxygenated RBCs resulted from a sustained volume-activated release of K+, Cl2and amino acids (AAs). In the
absence of ouabain, the cells also lost Na+ as released K+
was partially regained via the Na+/K+ pump. Inorganic
osmolytes contributed approximately 70 %, and organic osmolytes approximately 30 %, to the RVD of oxygenated RBCs. Oxygenation in isotonic medium per se activated a K+ efflux from the RBCs. Hypo-osmotic cell swelling
stimulated an additional K+ release. The
oxygenation-activated and the volume-oxygenation-activated K+ efflux were both
inhibited by DIDS and by the replacement of Cl2 with NO32, showing that both types of K+ efflux were Cl2
-dependent and probably occurred via the same K+/Cl2
cotransport mechanism. Once activated by oxygenation, the K+/Cl2 cotransport was further stimulated by cell
swelling. Deoxygenation inactivated the oxygenation-induced Cl2-dependent K+release and cell swelling was not
a sufficient stimulus to reactivate it significantly. In
deoxygenated RBCs, the volume-induced K+ release was
transient and primarily Cl2-independent and, in the absence of ouabain, the cell K+content recovered towards
control values via the Na+/K+pump. The Cl2-independent
K+ efflux seemed to involve K+/H+ exchange, but other
transport routes also participated. Swelling-activated AA release differed in kinetics between oxygenated and deoxygenated RBCs but was important for RVD at both oxygenation degrees. Approximately 70 % of the AA release was inhibited by DIDS and substitution of NO32for
Cl2 produced a 50 % inhibition, suggesting that the AA permeation was partly Cl2-dependent. In oxygenated RBCs, a reduction in pH lowered the volume-activated Cl2-dependent K+ efflux but not the AA efflux. In
deoxygenated RBCs, the acute volume-stimulated K+and
AA release were both increased by acidification. The data are discussed in relation to possible transducer mechanisms and physiological implications.
Key words: K+ transport, amino acid transport,
oxygenation-dependent osmolyte transport, membrane permeation in red blood cells, volume regulation.
Introduction
Summary
REGULATORY VOLUME DECREASE IN CARP RED BLOOD CELLS: MECHANISMS
AND OXYGENATION-DEPENDENCY OF VOLUME-ACTIVATED POTASSIUM AND
AMINO ACID TRANSPORT
FRANK B. JENSEN
Institute of Biology, Odense University, Campusvej 55, DK-5230 Odense M, Denmark
156
K+/Cl2 cotransport (Jensen, 1992; Nielsen et al. 1992), it is also pertinent to investigate the influence of pH on the RVD response in fish RBCs.
Previous studies on volume regulation in fish RBCs have been carried out on euryhaline species, such as flounder (Fugelli, 1967; Cala, 1977; Fincham et al. 1987), eel (Fincham
et al. 1987) and rainbow trout (Garcia-Romeu et al. 1991).
These species tolerate ambient salinities ranging from fresh water to full-strength sea water, and they benefit from effective volume regulatory mechanisms to minimize the effects of a changing extracellular osmolality on cell volume. In stenohaline freshwater species, such as carp, little is known about the extent of cell volume regulation in response to osmotic stress, nor whether the mechanisms of volume regulation are similar to those observed in euryhaline teleosts. The present paper analyzes the K+ and AA permeation mechanisms involved in RVD in carp RBCs and reports that both the degree of oxygenation and pH influence these mechanisms.
Materials and methods
Experimental animals
Carp (Cyprinus carpio L., mass 1–2 kg, N=42) were obtained from lakes at Gråsten Castle, Jutland, Denmark, and maintained at 15 ˚C under normoxic conditions (water
PO∑>18 kPa=135 mmHg) in 500 l holding tanks supplied with a constant inflow of tap water. The animals were subjected to a 12 h:12 h light:dark photoperiod and fed regularly on commercial fish food pellets.
Experimental protocol
Approximately 15 ml of blood was withdrawn from the caudal vessels of individual carp. After centrifugation and removal of plasma, the red blood cells (RBCs) were washed twice in a physiological carp saline consisting of: 130 mmol l21 NaCl, 12 mmol l21 NaHCO
3, 2.4 mmol l21 KH2PO4, 1 mmol l21 MgSO
4.7H2O, 2 mmol l21 CaCl2.2H2O, 3.9 mmol l21glucose. The cells were suspended at a fractional haematocrit of 0.20–0.25 and incubated overnight at 5 ˚C to ensure that the cells were at steady state and not in a catecholamine-stimulated condition (Bourne and Cossins, 1982). The cells were then centrifuged and resuspended in fresh saline at a fractional haematocrit of 0.226±0.016 (mean ± S.D., N=82). Two 5.3 ml samples were transferred to two Eschweiler tonometers and equilibrated at 15 ˚C with a humidified gas mixture supplied from cascaded Wösthoff (Bochum, Germany) gas-mixing pumps. A gas mixture of 0.2 % CO2/30 % O2/69.8 % N2 was used to oxygenate the RBCs, and a gas mixture of 0.2 % CO2/99.8 % N2was used to deoxygenate the RBCs. Anoxic incubation of blood from cyprinids may cause a slow decline in RBC nucleoside triphosphate content (Jensen and Weber, 1985). It was, however, verified that the nucleoside triphosphate content of deoxygenated RBCs stayed high (above 7 mmol l21RBC) for 220 min in the isotonic physiological saline. The RBCs were
pre-equilibrated in the tonometers for 35 min in order to guarantee equilibrium with respect to degree of oxygenation and PCO∑/pH. Following the pre-equilibration period, a control sample (time zero) was drawn from the tonometers.
The red cells in one tonometer were kept equilibrated at a constant level of oxygenation in the physiological saline throughout the experiment. The red cells from the other tonometer were subjected to hypo-osmotic swelling in order to study the mechanisms of regulatory volume decrease at the same level of oxygenation. The cell suspension from the latter tonometer was quickly moved to a tube containing an atmosphere of the equilibration gas. The tube was closed and centrifuged for 2 min. After removal of the supernatant, the cells were resuspended in a hypotonic saline equilibrated to the same gas mixture and temperature as the cells. This saline had the same composition as the physiological saline except that [NaCl] was reduced to 65 mmol l21. Following hypo-osmotic swelling, the cell suspension was immediately transferred back to the tonometer. Samples were drawn from the tonometer 5, 30, 60 and 140 min following the hypo-osmotic swelling. Parallel samples were drawn from the tonometer in which the cells were maintained in the physiological saline.
Experiments were performed on oxygenated and deoxygenated red cells in the absence and presence of various pharmacological drugs. Chemicals were added as microlitre samples from freshly prepared stock solutions 5 min before time zero (except quinine, which was added 20 min before time zero). Ouabain (Sigma Chemical Co.) was added to a final concentration of 2 mmol l21. DIDS (4,49-diisothiocyanostilbene-2,29-disulphonic acid) (Aldrich Chemical Co.) was added to a final concentration of 0.1 mmol l21. Furosemide (Aldrich Chemical Co.) was added to a final concentration of 1 mmol l21. Quinine (Sigma Chemical Co.) was added to a final concentration of 0.2 mmol l21. In experiments on the effects of substitution of nitrate for chloride, the whole experimental procedure (washing, incubation/equilibration, swelling) was performed using salines in which the chloride salts of the saline had been substituted with the corresponding nitrate salts at the same concentration.
The experiments were conducted at a high (physiological) pH (pH≈8.0) and at low pH (pH≈7.6). pH was lowered by elevating the gaseous CO2content to 1.0 %.
Measurements
157
K
+and amino acid transport in carp RBCs
volume of 12 % trichloroacetic acid (TCA, Sigma Chemical Co.) corresponding to twice the wet mass of red cells was added to the other sample of packed red cells. The deproteinized sample was then centrifuged and the supernatant used for determination of RBC ion and amino acid contents.
Extra- and intracellular concentrations of amino acids were determined spectrophotometrically as Ninhydrin-positive substances (e.g. Moore and Stein, 1948). Potassium was measured by atomic absorption spectrophotometry (Perkin-Elmer 2380). Sodium was measured with a flame photometer (Instrumentation Laboratory 243) and chloride was measured by coulometric titration (Radiometer CMT 10).
Extracellular pH was measured with a Radiometer (Copenhagen, Denmark) BMS3 electrode system thermostatted at 15 ˚C and connected to a PHM 73 monitor. The total haemoglobin (Hb) concentration was assessed spectrophotometrically after conversion of the Hb to cyanmethaemoglobin, using a millimolar extinction coefficient of 11 at 540 nm. Haematocrit (Hct) was determined by centrifugation. Red blood cell counts (NRBC) were performed using a Bürger-Türk counting chamber and a microscope (Nikon Alphaphot/YS). Mean cellular volume (MCV) was calculated from Hct/NRBC, and the red cell [Hb] was assessed from total [Hb]/Hct.
The data are presented as means ±S.E.M. and were analyzed by analysis of variance (ANOVA) for repeated measurements to determine within-treatment changes over time. Differences between treatments were evaluated by a two-way ANOVA,
where time was treated as a repeated factor, followed by the Tukey test.
Results
Changes in extracellular [K+]
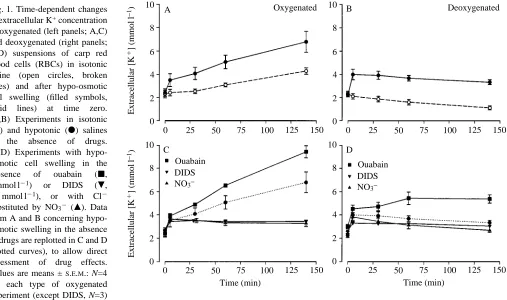
The extracellular K+ concentration increased as a function of time (P<0.0001) when carp RBCs were equilibrated in the oxygenated state in an isotonic saline at physiological pH (Fig. 1A). This net K+ efflux from the RBCs, which was inhibited by DIDS, by substitution of nitrate for chloride and by furosemide (data not shown), has been ascribed to an oxygenation-activated K+/Cl2cotransport mechanism (Jensen, 1992).
Hypo-osmotic swelling of oxygenated RBCs stimulated an additional release of K+. The swelling-induced K+efflux rate was particularly high during the first 5 min following cell swelling, but it remained elevated compared with that in non-swollen cells from 5 to 140 min (Fig. 1A), so that the extracellular potassium concentration increased to significantly (P<0.01) higher levels in swollen than in non-swollen oxygenated RBC suspensions. Treatment of the RBCs with ouabain significantly (P<0.05) increased the swelling-induced K+ efflux (Fig. 1C), showing that in the absence of ouabain some of the K+that was released from the cells was regained
via the Na+/K+pump.
[image:3.612.59.566.431.731.2]Deoxygenation of the RBCs in the isotonic saline eliminated the oxygenation-induced K+/Cl2 cotransport, and the
Fig. 1. Time-dependent changes
in extracellular K+concentration
of oxygenated (left panels; A,C) and deoxygenated (right panels; B,D) suspensions of carp red blood cells (RBCs) in isotonic saline (open circles, broken lines) and after hypo-osmotic cell swelling (filled symbols, solid lines) at time zero. (A,B) Experiments in isotonic
(s) and hypotonic (d) salines
in the absence of drugs. (C,D) Experiments with hypo-osmotic cell swelling in the
presence of ouabain (j,
2 mmol l21) or DIDS (.,
0.1 mmol l21), or with Cl2
substituted by NO32 (m). Data
from A and B concerning hypo-osmotic swelling in the absence of drugs are replotted in C and D (dotted curves), to allow direct assessment of drug effects.
Values are means ±S.E.M.: N=4
for each type of oxygenated experiment (except DIDS, N=3)
and N=5 for each type of deoxygenated experiment (except DIDS, N=3; ouabain, N=3). Temperature, 15 ˚C. 6
8 10
4
2
0
Extracellular [K
1] (mmol
l
−
1)
Extracellular [K
1] (mmol
l
−
1)
0 25 50 75 100 125 150
Oxygenated
6 8 10
4
2
0
0 25 50 75 100 125 150
Deoxygenated
6 8 10
4
2
0
0 25 50 75 100 125 150
Time (min) Ouabain
DIDS
NO3−
Ouabain DIDS
NO3−
6 8 10
4
2
0
0 25 50 75 100 125 150
Time (min)
B
C D
158
extracellular potassium concentration decreased with time as a result of a net RBC K+uptake via the Na+/K+pump (Fig. 1B; Jensen, 1990, 1992). Hypo-osmotic swelling of the deoxygenated cells caused a rapid elevation of extracellular [K+] at 5 min (P<0.01), following which the extracellular [K+] slowly decreased at a rate similar to that in iso-osmotic deoxygenated RBC suspensions (Fig. 1B). Thus, the swelling-induced K+ efflux was transient in deoxygenated RBCs and was succeeded by a net K+ uptake via the Na+/K+ pump. Administration of ouabain supported this interpretation. In ouabain-treated deoxygenated RBCs there was no net K+ uptake and the extracellular [K+] changed only slightly following the initial swelling-induced RBC K+ efflux (Fig. 1D).
The long-lasting K+ efflux mechanism induced in hypo-osmotically swollen oxygenated RBCs was significantly (P<0.01) inhibited by DIDS and by substitution of nitrate for chloride (Fig. 1C), suggesting major involvement of a Cl2 -dependent K+ transport mechanism, as in the case of oxygenation-induced K+ release. In contrast, the rapid K+ release observed in both oxygenated and deoxygenated cells during the first 5 min following hypo-osmotic swelling was more modestly affected by DIDS and by substitution of NO32 for Cl2 (Fig. 1C,D), suggesting a contribution from a Cl2 -independent mechanism. Treatment of NO32-substituted RBCs with quinine (which inhibits Ca2+-activated K+ channels) did not significantly change K+ movements compared with those found in NO32-substituted cells not treated with quinine (data not shown).
Changes in the extracellular amino acid concentration
In isotonic RBC suspensions, the extracellular AA concentration was low (approximately 0.3 mmol l21) and it stayed constant with time (Fig. 2A,B). Thus, the oxygenation-induced RBC K+release was not accompanied by AA release. Hypotonic stress activated a large AA efflux from the red cells. In oxygenated RBC suspensions, the mean extracellular AA concentration increased to 2.3 mmol l21 within 5 min (P<0.01) and to 5.2 mmol l21 between 5 and 140 min (Fig. 2A). In deoxygenated RBCs, the net AA efflux was larger (P<0.05) than that from oxygenated RBCs during the first 5 min following swelling (extracellular [AA] increasing to 3.7 mmol l21). The subsequent AA efflux from deoxygenated cells was, however, low (Fig. 2B). Thus, as with K+release (Fig. 1), the acute RBC AA release was larger in deoxygenated than in oxygenated cells, but a significant sustained release of cellular osmolytes was only seen from oxygenated cells.
The AA release from oxygenated and deoxygenated RBCs was not affected by ouabain but it was significantly (P<0.01) inhibited by DIDS and by substitution of NO32 for Cl2 (Fig. 2C,D). DIDS caused an inhibition of the AA release by some 70 %, whereas substitution of nitrate for chloride reduced the AA release to approximately 50 % of that observed in the physiological Cl2 saline, suggesting that the AA efflux was partly via a Cl2-dependent pathway. Quinine, at 0.2 mmol l21, had only a minor influence on the AA release in RBC suspensions with Cl2 substituted by NO32 (i.e. on the Cl2 -independent component; data not shown).
F. B. J
ENSEN6 8 10
4
2
0
Extracellular [AA] (mmol
l
−
1)
Extracellular [AA] (mmol
l
−
1)
0 25 50 75 100 125 150
Oxygenated
6 8 10
4
2
0
0 25 50 75 100 125 150
Deoxygenated
6 8 10
4
2
0
0 25 50 75 100 125 150
Time (min) Ouabain
DIDS
NO3−
Ouabain DIDS
NO3−
6 8 10
4
2
0
0 25 50 75 100 125 150
Time (min)
B
C D
[image:4.612.181.561.454.738.2]A
159
K
+and amino acid transport in carp RBCs
Volume-induced changes in red cell K+, amino acid, Cl2
and Na+contents
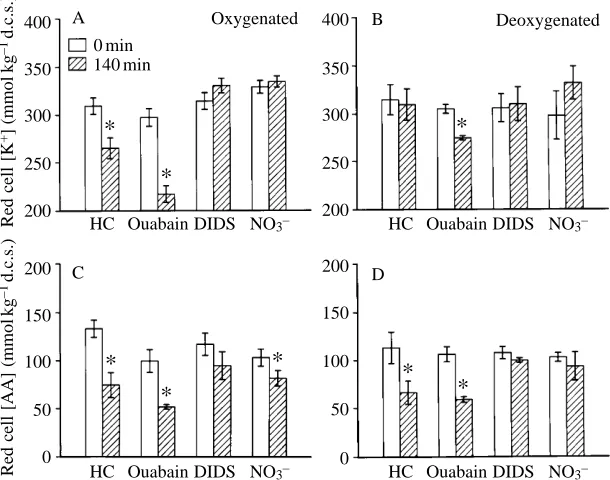
Extracellular changes in potassium and amino acid concentrations were paralleled by reciprocal changes in the intracellular K+and AA content. The changes in red cell [K+] and [AA] resulting from 140 min of exposure to the hypo-osmotic medium are illustrated in Fig. 3 for control cells as well as for ouabain-treated, DIDS-treated and nitrate-substituted cells.
In oxygenated control cells, hypo-osmotic swelling caused a significant decrease in both red cell [K+] and [AA] (HC in Fig. 3A,C). The expected elevation of extracellular concentrations (DXe, mmol l21) can be calculated from the decreases in intracellular concentrations (DXi, mmol kg21d.c.s.), the fractional haematocrit (F
Hct) and the RBC fractional water content (Fw) according to:
DXe = DXi 3 FHct(1 2Fw)/(1 2FHct) .
Thus, in oxygenated control RBCs at the prevailing Hct, the 44.5 and 58.8 mmol kg21d.c.s. decreases in red cell [K+] and [AA], respectively, should elevate extracellular [K+] and [AA] by 4.1 and 5.4 mmol l21, respectively, during the 140 min. These values are similar to the changes actually observed (Figs 1, 2), illustrating that the released cellular osmolytes could be recovered in the extracellular compartment.
In deoxygenated RBCs, the red cell AA content was significantly reduced after 140 min of hypo-osmotic exposure (Fig. 3D, HC) whereas the K+ content was not significantly different between 0 and 140 min (Fig. 3B, HC), which agrees with the elevation of extracellular [AA] (Fig. 2B) and re-uptake of released K+(Fig. 1B).
Ouabain treatment augmented the decrease in red cell [K+] of swollen oxygenated and deoxygenated cells (Fig. 3A,B),
whereas D[AA] was the same in treated and untreated cells (Fig. 3C,D). These results were also in general agreement with the measurements from the extracellular fluid. Similarly, the influence of DIDS and NO32substitution on red cell [K+] and [AA] confirmed their inhibitory effect on the volume-induced K+and AA release (Fig. 3).
The gradual release of K+ from oxygenated cells was accompanied by a gradual decrease in the red cell Cl2content, which was significantly (P<0.01) larger under hypo-osmotic than under isotonic conditions (Fig. 4A). This shows that the anion accompanying K+was Cl2. The decrease in Cl 2content of oxygenated RBCs during 140 min of hypotonic exposure (Fig. 4A) was somewhat larger than the decrease in K+content (Fig. 3A, HC). This was explained by a parallel decrease in red cell Na+ content of approximately 25 mmol kg21d.c.s. (not illustrated). In the presence of ouabain, the RBC Na+content was not significantly changed by hypo-osmotic swelling.
In deoxygenated red cells, the acute swelling-induced release of K+(Fig. 1B) was accompanied by a decrease in RBC Cl2content within the first 5 min (P<0.05), after which the Cl2 content increased slightly with time (Fig. 4B). Thus, as was the case with the RBC K+content (Fig. 3B, HC), the Cl2content of deoxygenated RBCs 140 min after hypo-osmotic swelling was not significantly different from initial Cl2levels.
Changes in red cell volume
Information on red cell volume changes was obtained by three independent measurements: changes in water content (Fig. 5A,B), actual volume (Fig. 5C,D) and mean cellular [Hb] (not illustrated), all of which gave the same general results.
In the isotonic physiological saline, the volume of oxygenated control RBCs and ouabain-treated RBCs decreased with time as a result of the oxygenation-induced KCl efflux;
400
350
300
250
200
400
350
300
250
200
Red cell [K
+] (mmol
kg
–1
d.c.s.)
200
150
100
50
0
200
150
100
50
0
Red cell [AA] (mmol
kg
–1
d.c.s.)
HC OuabainDIDS NO3–
HC OuabainDIDS NO3– HC OuabainDIDS NO3–
HC OuabainDIDS NO3–
Oxygenated 0 min
140 min
Deoxygenated A
C
B
D
*
*
*
*
*
*
*
[image:5.612.260.565.76.316.2]*
Fig. 3. Intracellular K+(A,B) and amino acid (C,D)
contents in oxygenated (A,C) and deoxygenated (B,D) carp RBCs before (open bars, time zero) and 140 min after hypo-osmotic shock (hatched bars) in the absence (HC, hypo-osmotic control) and
presence of ouabain or DIDS, or with Cl2substituted
by NO32. Asterisks indicate a significant difference
(P<0.05) between time zero and 140 min values.
Values are means ±S.E.M. N values are as described
160
the inhibition of this efflux by DIDS and by substitution of NO32 for Cl2 abolished the cell shrinkage (Fig. 5; Jensen, 1992).
Hypo-osmotic shock caused a large increase in cell volume of both oxygenated and deoxygenated RBCs. Swelling of oxygenated RBCs was followed by a regulatory volume decrease (RVD) that returned the volume of both control and ouabain-treated cells to values not significantly different from the original values within 140 min (Fig. 5A,C). Volume readjustment was very limited in DIDS-treated and NO32 -substituted RBCs.
In deoxygenated RBCs, the RVD response was incomplete under all experimental conditions (Fig. 5B,D).
pH changes during regulatory volume decrease
[image:6.612.203.560.75.214.2]Oxygenation-induced K+ efflux did not significantly affect pH (Fig. 6A), as shown earlier (Jensen, 1990). Osmolyte fluxes during RVD at physiological pH were, however, associated with a significant (P<0.0001) alkalization of the extracellular compartment (equivalent to an acidification of the intracellular compartment) (Fig. 6). The extracellular alkalization developed gradually. 5 min after hypo-osmotic swelling, extracellular pH was only moderately changed, whereas a significant pH elevation was present from 30 min onwards (Fig. 6). DIDS treatment accentuated the pH elevation at 5 min (increasing pH by 0.15 with oxygenated cells and 0.10 with deoxygenated cells) but did not significantly change pH at
F. B. J
ENSENFig. 4. Time-dependent changes in cellular chloride content of oxygenated (A) and deoxygenated (B) carp RBCs in an isotonic saline
(s) and following exposure to a
hypotonic saline (d). Asterisks
indicate a significant difference (P<0.01) between values in hypotonic and isotonic salines. Values are
means ±S.E.M., N=4 for each type of
oxygenated experiment and N=5 for each type of deoxygenated experiment. d.c.s., dry cell solids.
0 250
200
150
100
50
0 250
200
150
100
50
Red cell [Cl
–] (mmol
kg
–1
d.c.s.)
0 25 50 75 100 125 150
Time (min)
0 25 50 75 100 125 150
Time (min)
Oxygenated Deoxygenated
A B
*
*
*
1 2 3 4
1 2 3 4
Water content (g
H2
O
g
–1
d.c.s.)
150 200 250 300
150 200 250 300
MCV (fl
)
0 25 50 75 100 125 150 0 25 50 75 100 125 150
0 25 50 75 100 125 150 0 25 50 75 100 125 150
Oxygenated
A B
C D
Time (min) Time (min)
[image:6.612.155.560.429.740.2]Deoxygenated
Fig. 5. Water content (A,B) and mean cellular volume (MCV) (C,D) of oxygenated and deoxygenated carp RBCs in isotonic saline (open symbols, broken lines) and after hypo-osmotic shock (filled symbols, solid lines) in the absence
(d) and presence of
ouabain (j) or DIDS (.)
or with Cl2substituted by
NO32 (m). Values are
means ±S.E.M. N values
161
K
+and amino acid transport in carp RBCs
subsequent times (not illustrated). The pH changes during RVD were diminished by a reduction in pH (Fig. 6A,B).
Influence of pH on osmolyte fluxes and the regulatory volume decrease
A lowering of extracellular pH from a representative in vivo level (pH≈8) to values around pH 7.6 significantly influenced osmolyte fluxes during RVD. In deoxygenated RBCs, a
lowering of pH increased the acute releases of both K+ (Fig. 6D) and AAs (Fig. 6F), thus reducing the changes in cell volume following hypo-osmotic shock (Fig. 6H). In oxygenated RBCs, the K+release was slightly decreased by a reduction in pH (Fig. 6C), notably between 5 and 140 min, whereas the AA release appeared to be unaffected (Fig. 6E), leaving volume adjustments only marginally affected (Fig. 6G).
7.4 7.6 7.8 8.0 8.2 8.4
7.4 7.6 7.8 8.0 8.2 8.4
Extracellular pH
0 25 50 75 100 125 150
Oxygenated Deoxygenated
A B
C D
E F
G H
Time (min)
0 25 50 75 100 125 150
Time (min)
0 25 50 75 100 125 150 0 25 50 75 100 125 150
0 25 50 75 100 125 150 0 25 50 75 100 125 150
0 25 50 75 100 125 150 0 25 50 75 100 125 150
0 2 4 6 8 10
0 2 4 6 8 10
Extracellular [K
1] (mmol
l
−
1)
0 2 4 6 8 10
0 2 4 6 8 10
Extracellular [AA] (mmol
l
−
1)
1 2 3 4
1 2 3 4
Water content (g
H2
O
g
–1
[image:7.612.203.568.71.623.2]d.c.s.)
Fig. 6. Extracellular pH, potassium concentration, and amino acid concentration and RBC water content in oxygenated and deoxygenated suspensions of carp RBCs in isotonic saline (open symbols, broken lines) and following hypotonic cell swelling (filled symbols, solid lines) at
physiological pH (circles, Noxy=4;
Ndeoxy=5) and at low pH (triangles,
Noxy=3; Ndeoxy=3). Values are means
162
Discussion
Carp RBCs responded to hypo-osmotic swelling by a coordinated release of both inorganic (KCl) and organic (AAs) osmolytes. The overall RVD response of carp RBCs accordingly resembles that seen in other fish RBCs. It is, however, evident from the present study, that the RVD response and its underlying mechanisms are strongly dependent on both the degree of oxygenation of the cells and on pH.
Oxygenation-dependence of regulatory volume decrease
In oxygenated RBCs, hypo-osmotic swelling triggered a sustained release of K+(Fig. 1) and AAs (Fig. 2) that restored the original cell volume within 140 min (Fig. 5). The resulting decreases in intracellular concentrations (Fig. 3) and increases in extracellular concentrations (Figs 1, 2) were slightly larger for AAs than for K+. However, since the net K+ efflux is followed by a net efflux of Cl2(and of Na+in the absence of ouabain, see below), the contribution of inorganic osmolytes to RVD in oxygenated control RBCs is larger (approximately 70 %) than that of AAs (approximately 30 %).
Deoxygenated RBCs showed an incomplete volume recovery following osmotic swelling (Fig. 5). A large AA release was observed (Fig. 2), but K+ was only transiently released and was regained via the Na+/K+pump (Fig. 1). Thus, although the AA content of RBCs was reduced 140 min after the hypo-osmotic shock, the K+content had returned to control values (Fig. 3). In the presence of ouabain, a lasting K+release limited cell swelling and improved volume recovery but the RVD response remained incomplete (Figs 1, 3, 5). Ouabain-treated RBCs from rainbow trout show a similar incomplete RVD response when deoxygenated (Garcia-Romeu et al. 1991) and complete volume recovery when oxygenated (Nielsen et
al. 1992), as found here for carp both in the absence and in the
presence of ouabain. The RVD response may accordingly be oxygenation-dependent in general in fish RBCs. In order to understand this oxygenation-dependency it is necessary to analyze the mechanisms of osmolyte permeation.
Mechanisms of potassium permeation
As previously described in detail in carp (Jensen, 1990, 1992) and rainbow trout (Borgese et al. 1991; Nielsen and Lykkeboe, 1992; Nielsen et al. 1992), oxygenation per se activates a ouabain-resistant K+ efflux that is inhibited by DIDS, by furosemide and by substitution of NO32 for Cl2. This Cl2-dependent K+ permeation can be ascribed to a K+/Cl2 cotransport mechanism and is inactivated by deoxygenation.
Following hypo-osmotic swelling, potassium permeation of the carp RBC membrane apparently involved both Cl2 -dependent and Cl2-independent pathways of differing kinetics and importance for RVD in oxygenated and deoxygenated RBCs. The Cl2-independent mechanism (only marginally affected by NO32 substitution) dominated the K+ efflux immediately after cell swelling in both oxygenated and deoxygenated RBCs, but then became less important. The Cl2
-dependent mechanism (inhibited by NO32 substitution) provided a long-lasting K+efflux in oxygenated cells but was absent or only marginally active in deoxygenated cells. Thus, activation of the Cl2-dependent K+pathway is needed for the complete RVD response in oxygenated RBCs and its inactivation contributes to the incomplete RVD in deoxygenated RBCs.
The Cl2-dependent K+permeation
The swelling-induced Cl2-dependent K+ efflux in oxygenated RBCs closely resembled the oxygenation-induced Cl2-dependent K+ efflux by being effectively inhibited by NO32 substitution, by DIDS and by deoxygenation (Fig. 1), and both types of K+ efflux may occur via the same K+/Cl2 cotransport mechanism. Activation of the K+/Cl2 cotransporter by oxygenation in an isotonic medium causes cell shrinkage in the absence of a prior perturbing cell volume increase. However, once activated by oxygenation, the Cl2 -dependent K+efflux can be further stimulated by cell swelling (Fig. 1), leading to an appropriate RVD response. This conclusion is supported by recent data on rainbow trout RBCs, in which the oxygenation-activated K+transport was shown to be volume-sensitive: cell swelling increased the K+ flux and cell shrinkage decreased it (Nielsen et al. 1992). When, however, the Cl2-dependent K+flux has been deactivated by deoxygenation, a substantial hypo-osmotically induced cell swelling (Fig. 5) was not a sufficient stimulus to reactivate it significantly (Fig. 1). Involvement of the Cl2-dependent K+ efflux mechanism in the RVD response following hypotonic swelling thus appears to be crucially dependent on the cells being oxygenated. This observation challenges the suggestion that hypotonic swelling is associated with a Cl2-independent K+ permeation by means of decreasing intracellular ionic strength (Motais et al. 1991).
Sustained swelling-induced Cl2-dependent K+release from oxygenated RBCs was reduced by a reduction in pH (Fig. 6). This pH-sensitivity is unlike that of the K+/Cl2 cotransporter in young human RBCs, which is stimulated by acidification (Lauf et al. 1992). The inhibition of the swelling-induced K+/Cl2cotransport by acidification is, however, similar to that observed for oxygenation-induced K+/Cl2cotransport (Jensen, 1992; Nielsen et al. 1992), corroborating the similarity between the permeation mechanisms and supporting the idea that the activity of the cotransporter can be reduced by shifting the allosteric equilibrium between R and T structure haemoglobin towards the T-state (see Jensen, 1992).
The Cl2-independent K+permeation
163
K
+and amino acid transport in carp RBCs
nature of osmolyte release. Intracellular sodium content was not significantly changed in carp RBCs in the presence of ouabain, and in its absence the cellular [Na+] actually decreased.
The Cl2-independent K+permeation may have occurred via conductive transport but it was not significantly affected by quinine, suggesting that Ca2+-activated K+channels, known to mediate conductive K+ transport during RVD in some other cell types (Hoffmann et al. 1988), are not involved. Similarly, in rainbow trout, the Cl2-independent K+flux is not via a Ca2+ -activated pathway (Garcia-Romeu et al. 1991), and recent data have shown that the Cl2-independent K+loss during RVD is electrically silent and not via conductive transport (Guizouarn
et al. 1993). Electrically silent movements of K+ can be mediated either by K+/anion cotransport or by K+/H+ exchange. In trout, K+/H+exchange was ruled out, and it was suggested that both the Cl2-independent and the Cl2 -dependent K+ losses occur via coupled K+/anion cotransport (Guizouarn et al. 1993).
In carp, K+/H+ exchange may be involved. Under normal conditions without inhibitors, both K+/H+ exchange (coupled to Cl2/HCO32exchange via the band 3 anion exchanger) and K+/Cl2 cotransport (which secondarily activates the anion exchanger) could cause a net transfer of acid equivalents into the cells (Nikinmaa, 1990), contributing to the extracellular alkalization during RVD (Fig. 6). Both the anion exchanger and the K+/Cl2 cotransporter in fish RBCs are inhibited by DIDS, so fluxes of both K+ and acid equivalents should be strongly inhibited in the presence of DIDS if the Cl2 -independent K+transfer is via K+/Cl2cotransport. If coupled K+/H+and Cl2/HCO
32exchange is involved, however, then a net K+ release would be expected together with an enhancement of the extracellular pH increase. The observed acute K+release (Fig. 1C,D) and accentuated pH elevation (see Results) 5 min after swelling in DIDS-treated cells suggests that K+/H+ exchange contributes to the Cl2-independent K+ flux during RVD. However, it is also evident that part of the acute K+ release is inhibited by DIDS (notably in deoxygenated cells, see Fig. 1), suggesting that additional mechanisms must be involved. These could either be conductive transport via a mechanism that is not Ca2+ -activated or K+/anion cotransport, as proposed for deoxygenated trout RBCs (Guizouarn et al. 1993). Further experiments are needed to identify fully the Cl2-independent K+flux in carp and explain its apparent transient nature.
Involvement of the Na+/K+pump
The RVD response of oxygenated RBCs was complete both in the absence and in the presence of ouabain (Fig. 5), even though the K+release was larger in its presence (Fig. 1). Since AA release is unaffected by ouabain (Fig. 2), the RVD response in the absence of ouabain must be aided by release of cations in addition to K+. Indeed, in the absence of ouabain, the red cell Na+ content decreased. The Na+/K+ pump was stimulated by the elevated external [K+], and the cells regained part of the released K+and lost Na+. Thus, the Na+/K+pump
is involved in RBC volume decrease following hypotonic swelling, as is also the case following b-adrenergic swelling (O. B. Nielsen, personal communication).
Involvement of amino acids
Amino acids make a substantial contribution to the intracellular osmotic pressure in carp RBCs and play an important role in cellular volume regulation, as in more euryhaline teleosts such as flounder, eel and trout (Fugelli, 1967; Fugelli and Zachariassen, 1976; Fincham et al. 1987; Garcia-Romeu et al. 1991) and in seawater elasmobranchs (Goldstein et al. 1990). The high AA concentration in fish RBCs is dependent on active accumulation via a Na+ -dependent amino acid transport system (Fincham et al. 1987). This uptake may be inhibited by cell swelling (Fincham et al. 1987), but the major contribution to RVD comes from regulation of AA leak pathways.
Under iso-osmotic conditions, the extracellular AA concentration was low and stayed constant with time in both oxygenated and deoxygenated RBC suspensions (Fig. 2), showing the AA permeability of volume-static RBCs is low and unaffected by a change in the level of oxygenation. The AA permeability is, however, not zero (since the cells were initially washed and equilibrated with an AA-free saline). A balance is established between passive leak and active uptake of AAs.
Hypo-osmotic swelling of the RBCs triggered an increase in membrane permeability to AAs, which diffused out from the cells. Acute AA release (at 5 min) was larger in deoxygenated than in oxygenated cells, whereas subsequent sustained release was more prominent in the oxygenated cells (Fig. 2). Thus, the volume-activated AA efflux was dependent on the oxygenation status of the cells (though to a lesser degree than with K+ fluxes). This finding adds AA transport to the list of oxygenation-sensitive transport routes in fish RBCs (which includes K+/Cl2cotransport, adrenergic Na+/H+exchange and conductive transport). Furthermore, the acute AA release from deoxygenated RBCs was accentuated by a pH decrease (Fig. 6), whereas no clear pH-dependence was found with oxygenated cells.
The AA efflux mechanism is not fully understood. The volume-activated AA release is strongly inhibited by DIDS (Fig. 2) and is known to be Na+-independent (Fincham et al. 1987; Garcia-Romeu et al. 1991; Kirk et al. 1992). In trout RBCs, the swelling-induced AA loss is little affected by substitution of NO32 for Cl2 (Garcia-Romeu et al. 1991), whereas in carp RBCs it was approximately halved (Fig. 2), indicating that the AA loss is partly Cl2-dependent.
On the basis of the DIDS inhibition, an involvement of the anion exchanger (band 3) has been suggested (Goldstein et al. 1990). This idea is supported by a low-volume-activated AA efflux in cyclostome RBCs deficient in band 3 protein (Brill et
al. 1992). Alternative proposals are that the volume-activated
164
Transducer mechanisms
The observation that DIDS inhibits both volume-activated K+and AA fluxes suggests that DIDS interacts with a central target in the control of volume-sensitive transport systems (Garcia-Romeu et al. 1991). It has been proposed that the cytoskeleton, which is anchored to the membrane (e.g. to band 3), could function as a volume sensor and, via a conformational change brought about by cell deformation, could activate the transport systems (Motais et al. 1991). In this model, the binding of DIDS to band 3 would cause a conformational change that would be transmitted to the cytoskeleton and possibly to other transport sites in the membrane (Motais et al. 1991). A similar argument applies to the different binding of deoxy- and oxyhaemoglobin to the cytoplasmic fragment of band 3 (Salhany, 1990), which may be essential for the oxygenation-induced K+efflux (Jensen, 1990; Borgese et al. 1991; Nielsen et al. 1992) as well as the oxygenation-dependence of volume-activated K+ (Fig. 1; Nielsen et al. 1992) and AA (Fig. 2) fluxes.
K+/Cl2 cotransport in mammalian RBCs is governed by phosphorylation/dephosphorylation of an unidentified protein and is sensitive to sulphhydryl group modification (Jennings and Schulz, 1991; Cossins, 1991; Lauf et al. 1992). The same is true of the oxygenation-activated K+/Cl2cotransport in trout RBCs (Cossins et al. 1994). These events may, of course, involve transport or regulatory proteins other than band 3, but it is interesting to note that the cytoplasmic fragment of band 3, to which haemoglobin and the cytoskeleton bind, also contains critical sulphhydryl groups (whose modification by sylphhydryl-reactive reagents alters conformation) and amino acid residues that are phosphorylated by protein kinase activity (which alters cytosolic protein binding) (Salhany, 1990). Further study is needed in order to merge the various factors influencing KCl (and AA) transport into an integrated model.
Physiological implications
Carp do not normally experience the same fluctuations in ambient and extracellular osmolality as euryhaline species, but they may depend on the mechanisms of RVD for steady-state volume regulation and in situations where the extracellular osmolality is decreased by factors other than a decreased ambient salinity (for instance, during acid or metal exposures that cause a net ion efflux at the gills). Additionally, teleost RBCs can be isotonically swollen in stressful situations by b -adrenergic stimulation of Na+/H+ exchange across the RBC membrane (Nikinmaa, 1990). Volume regulation following b -adrenergic stimulation of trout RBCs resembles that seen following hypotonic swelling in that it is more complete at high than at low oxygen saturation (O. B. Nielsen, personal communication). An up-regulated RBC volume at low oxygen saturation may be advantageous by contributing to increased haemoglobin oxygen affinity under hypoxic conditions. Thus, an increased RBC volume could increase oxygen affinity by reducing the cellular concentrations of haemoglobin and nucleoside triphosphates and thus their complex binding (Weber, 1982). Such a specific influence of cell volume on
oxygen affinity is supported by data on carp haemoglobin (Lykkeboe and Weber, 1978) and by in vivo and in vitro studies with intact shrunken (Jensen, 1990) and swollen (K. Holk and G. Lykkeboe, personal communication) RBCs. The incomplete volume regulation of deoxygenated carp RBCs following hypotonic swelling may accordingly reflect the need for an up-regulated RBC volume in hypoxic conditions.
Miss Bente Frost Jacobsen is thanked for skilful technical assistance. The work was supported by the Danish Natural Science Research Council (11-9659-1).
References
BORGESE, F., MOTAIS, R. ANDGARCIA-ROMEU, F. (1991). Regulation of Cl-dependent K transport by oxy-deoxyhemoglobin transitions in trout red cells. Biochim. biophys. Acta 1066, 252–256. BOURNE, P. K. ANDCOSSINS, A. R. (1982). On the instability of K+
influx in erythrocytes of the rainbow trout, Salmo gairdneri and the role of catecholamine hormones in maintaining in vivo influx activity. J. exp. Biol. 101, 93–104.
BRILL, S. R., MUSCH, M. W. AND GOLDSTEIN, L. (1992). Taurine
efflux, band 3 and erythrocyte volume of the hagfish (Myxine glutinosa) and lamprey (Petromyzon marinus). J. exp. Zool. 264, 19–25.
CALA, P. M. (1977). Volume regulation by flounder red blood cells
in anisotonic media. J. gen. Physiol. 69, 537–552.
COSSINS, A. R. (1991). A sense of cell size. Nature 352, 667–668. COSSINS, A. R., WEAVER, Y., LYKKEBOE, G. AND NIELSEN, O. B.
(1994). The role of protein phosphorylation in the control of K flux pathways of trout red cells. Am. J. Physiol. (in press).
FINCHAM, D. A., WOLOWYK, M. W. AND YOUNG, J. D. (1987). Volume-sensitive taurine transport in fish erythrocytes. J. Membr. Biol. 96, 45–56.
FUGELLI, K. (1967). Regulation of cell volume in flounder (Pleuronectes flesus) erythrocytes accompanying a decrease in plasma osmolarity. Comp. Biochem. Physiol. 22, 253–260. FUGELLI, K. AND ZACHARIASSEN, K. E. (1976). The distribution of
taurine, gamma-aminobutyric acid and inorganic ions between plasma and erythrocytes in flounder (Platichthys flesus) at different plasma osmolalities. Comp. Biochem. Physiol. 55A, 173–177.
GARCIA-ROMEU, F., COSSINS, A. R. AND MOTAIS, R. (1991). Cell volume regulation by trout erythrocytes: characteristics of the transport systems activated by hypotonic swelling. J. Physiol., Lond. 440, 547–567.
GOLDSTEIN, L., BRILL, S. R. ANDFREUND, E. V. (1990). Activation of taurine efflux in hypotonically stressed elasmobranch cells: inhibition by stilbene disulfonates. J. exp. Zool. 254, 114–118. GUIZOUARN, H., HARVEY, B. J., BORGESE, F., GABILLAT, N., GARCIA
-ROMEU, F. AND MOTAIS, R. (1993). Volume-activated Cl2
-independent and Cl2-dependent K+pathways in trout red blood
cells. J. Physiol., Lond. 462, 609–626.
HOFFMANN, E. K., LAMBERT, I. H. AND SIMONSEN, L. O. (1988). Mechanisms in volume regulation in Ehrlich ascites tumor cells. Renal Physiol. Biochem. 11, 221–247.
JENNINGS, M. L. ANDSCHULZ, R. K. (1991). Okadaic acid inhibition of KCl cotransport. Evidence that protein dephosphorylation is necessary for activation of transport by either cell swelling or N-ethylmaleimide. J. gen. Physiol. 97, 799–818.
165
K
+and amino acid transport in carp RBCs
JENSEN, F. B. (1990). Nitrite and red cell function in carp: control factors for nitrite entry, membrane potassium ion permeation, oxygen affinity and methaemoglobin formation. J. exp. Biol. 152, 149–166.
JENSEN, F. B. (1992). Influence of haemoglobin conformation, nitrite
and eicosanoids on K+ transport across the carp red blood cell
membrane. J. exp. Biol. 171, 349–371.
JENSEN, F. B. ANDWEBER, R. E. (1985). Kinetics of the acclimational responses of tench to combined hypoxia and hypercapnia. I. Respiratory responses. J. comp. Physiol. B 156, 197–203.
KIRK, K., ELLORY, J. C. ANDYOUNG, J. D. (1992). Transport of organic
substrates via a volume-activated channel. J. biol. Chem. 267, 23475–23478.
LAUF, P. K. (1982). Evidence for chloride dependent potassium and
water transport induced by hyposmotic stress in erythrocytes of the marine teleost, Opsanus tau. J. comp. Physiol. 146, 9–16.
LAUF, P. K., BAUER, J., ADRAGNA, N. C., FUJISE, H., ZADE-OPPEN, A.
M. M., RYU, K. H. AND DELPIRE, E. (1992). Erythrocyte KCl
cotransport: properties and regulation. Am. J. Physiol. 263, C917–C932.
LYKKEBOE, G. ANDWEBER, R. E. (1978). Changes in the respiratory properties of the blood in the carp, Cyprinus carpio, induced by diurnal variation in ambient oxygen tension. J. comp. Physiol. 128, 117–125.
MOORE, S. ANDSTEIN, W. H. (1948). Photometric ninhydrin method
for use in the chromatography of amino acids. J. biol. Chem. 176, 367–388.
MOTAIS, R., GUIZOUARN, H. ANDGARCIA-ROMEU, F. (1991). Red cell volume regulation: the pivotal role of ionic strength in controlling swelling-dependent transport systems. Biochim. biophys. Acta
1075, 169–180.
NIELSEN, O. B. ANDLYKKEBOE, G. (1992). In vitro effects of pH and
hemoglobin-oxygen saturation on plasma and erythrocyte K+levels
in blood from trout. J. appl. Physiol. 72, 1291–1296.
NIELSEN, O. B., LYKKEBOE, G. AND COSSINS, A. R. (1992). Oxygenation-activated K fluxes in trout red blood cells. Am. J. Physiol. 263, C1057–C1064.
NIKINMAA, M. (1990). Vertebrate Red Blood Cells. Adaptations of Function to Respiratory Requirements. Berlin: Springer.
SALHANY, J. M. (1990). Erythrocyte Band 3 Protein. Boca Raton, Fl: CRC Press.
THOROED, S. M. AND FUGELLI, K. (1994). The Na+-independent taurine influx in flounder erythrocytes and its association with the volume regulatory taurine efflux. J. exp. Biol. 186, 245–268.
WEBER, R. E. (1982). Intraspecific adaptation of hemoglobin function