Suppressor Analysis to Identify Proteins that Work with the
Mycobacterium
smegmatis
SecA2 Protein Export System
Lauren Stone Ligon
A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Department of Microbiology and Immunology, School of Medicine.
Chapel Hill 2013
Approved by:
iii
ABSTRACT
LAUREN STONE LIGON: Suppressor Analysis to Identify Proteins that Work with the
Mycobacterium smegmatis SecA2 Protein Export System (Under the direction of Miriam Braunstein)
Mycobacterium tuberculosis, the causative agent of the disease tuberculosis, is a serious threat to human health, responsible for 1.4 million deaths annually. When M. tuberculosis is spread from person to person by aerosol and is phagocytosed by alveolar macrophages, the proteins exported by the bacterium are ideally positioned to interact with the host. Therefore, exported proteins and the systems responsible for their export are important for M. tuberculosis
virulence. All bacteria, including mycobacteria, contain the Sec protein export system, which is responsible for the bulk of protein export and is composed of a motor ATPase protein, SecA, and a membrane-embedded channel complex, SecYEG. Mycobacteria, including both M.
tuberculosis and the model organism Mycobacterium smegmatis, along with some Gram-positive bacteria, are unique in containing two functionally distinct SecA proteins. The SecA protein responsible for housekeeping protein export is termed SecA1 and is essential for cell survival, while the second SecA protein, termed SecA2, is required for the export of a small subset of proteins and is important for M. tuberculosis virulence. While the canonical Sec system is well understood, the mechanism of SecA2-dependent export is not. Like canonical SecA proteins, the mycobacterial SecA2 requires ATPase activity to function. Furthermore, an
iv
at the membrane. Here, we analyze extragenic suppressors of secA2 K129R in an effort to better understand SecA2-dependent export and identify additional components of the SecA2 pathway. Using this approach, we demonstrate a connection between M. smegmatis SecA2 and SecY, the major membrane-embedded component of the housekeeping Sec export channel. In addition, we demonstrate a connection between the SecA2 system and Msmeg_1684, a protein of unknown function that is found throughout mycobacteria and may represent a novel
component of the SecA2 pathway. Our findings suggest a new model in which the
v
ACKNOWLEDGEMENTS
I would like to thank the following people who contributed to the work described in this dissertation. Dr. Nathan Rigel isolated the suppressor strains discussed in Chapters 2 and 3. Artur Romanchuk and Dr. Corbin Jones performed whole genome alignment and mutation identification for suppressors 4S in Chapter 2 and 7S and 20B in Chapter 3. Brittany Miller performed the SecA1 overexpression experiment in Chapter 2, the quantitative RT-PCR on genes
msmeg_1677-1683 in Chapter 3, and the msmeg_1684 deletion phenotype testing in Chapter 3.
In addition, I would like to thank several people who cloned or provided plasmids and strains used in this work. Dr. Adrie Steyn provided plasmid pCV125, used in Chapter 2. Dr. William Jacobs provided plasmid pYUB2063, used in Chapter 2. Brandon Anjuwon-Foster created strain BAF1, used in Chapter 3. Chelsea Moriarty created plasmid pCM2 and strains CM11 and CM14, used in Chapter 3. Courtney Sutphen created plasmids pCS1-4, used in Chapter 3. Meagan Martin created plasmids pMEM3 and pMEM4, used in Chapter 3. Dr. Henry Gibbons created plasmid pHSG93, used in Chapter 4.
vi
TABLE OF CONTENTS
LIST OF TABLES ... vii
LIST OF FIGURES ... viii
LIST OF ABBREVIATIONS AND SYMBOLS ... x
Chapter
I. Introduction ... 1
II. Suppressor Analysis Reveals a Role for SecY in the SecA2-
Dependent Protein Export Pathway of Mycobacteria ... 24
III. Identification of Mycobacterium smegmatis secA2 K129R
Suppressor Mutations in Gene msmeg_1684 ... 62
IV. Searching for Evidence of a Physical Interaction Between
Mycobacterium smegmatis SecA2 and SecY ... 117
V. Discussion ... 138
vii
LIST OF TABLES
Table
1.1 M. smegmatis and M. tuberculosis homologs of
conserved export system components ... 16
2.1 Mycobacterium smegmatis strains used in this study ... 48
2.2 Plasmids used in this study ... 49
2.3 Primers used in this study ... 50
3.1 Plasmids used in this study ... 94
3.2 Primers used in this study ... 95
3.3 Mycobacterium smegmatis strains used in this study ... 97
3.4 Genes found in suppressor 7S and 20B deletions ... 98
3.5 Suppressors affecting msmeg_1684 ... 99
4.1 Mycobacterium smegmatis strains used in this study ... 130
4.2 Plasmids used in this study ... 131
viii
LIST OF FIGURES
Figure
1.1 A preliminary model of the mycobacterial housekeeping Sec and
SecA2-dependent protein export systems ... 17
2.1 Azide sensitivity and Mueller-Hinton agar growth phenotypes ... 51
2.2 4S and 24S mutations are located within the secY promoter ... 52
2.3 Suppressor mutations cause increased secY expression ... 53
2.4 Suppressor mutations increase SecY protein levels relative to the secA2 K129R strain ... 54
2.5 Increased SecY levels are sufficient to suppress secA2 K129R ... 55
2.6 Additional extragenic suppressors increase SecY protein levels, but not secY transcript levels ... 56
2.7 SecA1 depletion does not affect SecY and is required for SecA2-dependent protein export ... 57
2.8 Overexpression of M. smegmatis SecA1 does not cause SecY degradation .... 58
3.1 Azide sensitivity and Mueller-Hinton agar growth phenotypes ... 100
3.2 Suppressors affect subcellular localization of SecA1 and SecA2 ... 101
3.3 Suppressors 7S and 20B contain large deletions ... 102
3.4 Genomic region affected by 7S and 20B suppressors ... 103
ix
3.6 A 15-gene region is responsible for suppression ... 105
3.7 Genes msmeg_1677-1683 are expressed in suppressor 20B ... 106
3.8 Suppressor mutations do not affect SecA1 and SecA2 protein levels ... 107
3.9 Eight suppressor mutations affect msmeg_1684 ... 108
3.10 Δmsmeg_1684 can suppress secA2 K129R phenotypes ... 109
3.11 Export-related stress causes increased secA2 expression from the hsp60 promoter ... 110
3.12 Suppressor strains can suppress secA2 K129R expressed from its native promoter ... 111
3.13 Alignment of Msmeg_1684 and Rv3311 proteins ... 112
3.14 Phylogenetic tree of Msmeg_1684-like proteins showing co-occurrence of SecA2 proteins ... 113
4.1 Schematic representation of experimental design for protein crosslinking and purification ... 133
4.2 SecY does not crosslink to SecA2-His ... 134
4.3 SecY does not crosslink to SecA2 K129R-His ... 135
x
LIST OF ABBREVIATIONS AND SYMBOLS
Δ deletion
2D Two-dimensional
Å angstrom
β-ME beta-mercaptoethanol
µg microgram
µL microliter
A adenine
ABC ATP binding cassette ADP adenosine diphosphate ANOVA analysis of variance
Asp aspartate
Atc anhydrotetracycline ATP adenosine triphosphate ATPase ATP hydrolase
BLAST basic local alignment search tool BLAT BLAST-like alignment tool
bp base pair
C Celsius or cytosine
C. Clostridium or Corynebacteria
xi CW cell wall fraction
Cys cysteine
DNA deoxyribonucleic acid
DSP dithiobis(succinimidyl proprionate)
E glutamate
E. Escherichia
ECF enhanced chemifluorescence ENV envelope fraction
ESX ESAT secretion system
F phenylalanine
g gravity
G guanine or glycine
Gly glycine
HA hemagglutinin
His hexahistidine tag
HIV Human Immunodeficiency Virus
hyg hygromycin resistance gene
I isoleucine
IgG Immunoglobulin G
K lysine
kan kanamycin resistance gene kbp kilobase pair
kDa kilodalton
xii
L. Listeria
M molar or methionine
M. Mycobacterium
Mb mega base pair
MDR Multi-drug resistant MEM membrane fraction
Met methionine
mg milligram
min minute
mL milliliter
mM millimolar
msmeg_ Mycobacterium smegmatis gene designation Msmeg_ Mycobacterium smegmatis protein designation
n nucleotide
N asparagine
N/A not applicable
NCBI National Center for Biotechnology Information
Ncgl Clostridium glutamicum gene designation Ni-NTA nickel nitrilotriacetic acid
ng nanogram
nm nanometer
N-terminal amino-terminal N-terminus amino-terminus
xiii
P promoter
PAGE polyacrylamide gel electrophoresis PCR polymerase chain reaction
Pi inorganic phosphate
Pro proline
Q glutamine
R arginine
revTetR reverse Tet repressor
RLM-RACE RNA ligase mediated rapid amplification of cDNA ends RNA ribonucleic acid
RT reverse transcriptase
rv Mycobacterium tuberculosis gene designation
Rv Mycobacterium tuberculosis protein designation
S serine
S. Streptococcus
SDAD NHS-SS-Diazirine SDS sodium dodecyl sulfate
Sec secretion
SNP single nucleotide polymorphism
SOAP short oligonucleotide alignment program SOD superoxide dismutase
SOL soluble fraction SP signal peptidase
xiv
T thymine
TAP tobacco acid pyrophosphatase Tat Twin-arginine translocation
TB tuberculosis
Thr threonine
Tn-seq transposon deep sequencing TraSH transposon site hybridization
Trp Tryptophan
UV ultraviolet
Val valine
WCL whole cell lysate
XDR extensively drug resistant
CHAPTER 1
Introduction
1Mycobacterium tuberculosis is the causative agent of the disease tuberculosis, which the World Health Organization estimates kills 1.4 million people every year (1). This pathogen is spread from person to person by aerosols created by sneezing and coughing. When aerosols are inhaled by an uninfected person, M. tuberculosis is able to reach the alveolar spaces and is taken up by alveolar macrophages. However, M. tuberculosis is not killed by macrophages, but rather can survive and persist, sometimes for decades, before eventually causing active disease. HIV-positive individuals are especially susceptible to developing active tuberculosis disease and, as a result, HIV/tuberculosis co-infection is the primary cause of death worldwide for the HIV-infected population. No effective vaccine is available and although tuberculosis is treatable, it requires a long antibiotic regimen that is expensive and difficult to complete, especially for people in the developing world. Furthermore, multidrug resistant (MDR) and extensively drug resistant (XDR) M. tuberculosis strains make treatment even more problematic. Therefore, novel drugs are desperately needed to combat this disease.
Because of their importance to virulence and bacterial viability, the exported proteins of
M. tuberculosis and their respective protein export systems can be considered potential drug
1
2
targets (recently reviewed in (2)). Many exported proteins, which we define as including proteins in the cell envelope of M. tuberculosis and proteins secreted by M. tuberculosis, interact with the host and are crucial for causing disease. Without its systems for transporting these proteins across the cytoplasmic membrane and to their final destination, M. tuberculosis
cannot deliver effector proteins that are necessary for virulence. Furthermore, some M. tuberculosis protein export systems are essential.
In M. tuberculosis, as well as the non-pathogenic model organism Mycobacterium smegmatis, there are two highly conserved protein export systems that are responsible for the majority of protein export: the Sec and Tat export pathways. M. tuberculosis and M. smegmatis
also possess specialized protein export systems dedicated to the export of a more limited set of proteins: the accessory SecA2 export pathway and ESX pathways. ESX pathways are also referred to as Type VII secretion systems. Interestingly, both of these specialized protein export systems were first identified in M. tuberculosis but later found to also exist in some other bacteria. A better understanding of the mycobacterial protein export systems could lead to new strategies to combat tuberculosis disease. This dissertation describes efforts to understand the mechanism of the SecA2 export pathway, which is important for M. tuberculosis virulence.
Housekeeping Sec Export System
3
comes from extensive study in Escherichia coli (for recent reviews see (4, 5)). While Sec export functions in essentially the same manner in all bacteria in which it has been studied, only a few aspects of this system have been directly investigated in mycobacteria.
Exported proteins and targeting. The Sec system exports proteins across the cytoplasmic membrane post-translationally. Proteins exported by the Sec system, termed preproteins or precursors, are synthesized with an N-terminal signal peptide that is important for targeting the protein for export. Following export, the signal peptide is cleaved to generate the mature protein. Sec signal peptides consist of a positively charged N-terminus, a
hydrophobic central domain, and an uncharged polar C-terminus containing the cleavage site (5). Some Sec-exported proteins are lipoproteins, and in these cases the C-terminal end of the signal peptide contains a lipobox motif with a conserved cysteine that is the site of lipid attachment (6). Sec signal peptides and lipoprotein signal peptides can be predicted using the bioinformatic programs PSORTb v3.0 (7), SignalP v4.0 (8), and LipoP v1.0 (9).
Proteins must be in an unfolded state to be exported by the Sec system. In many Gram-negative bacteria there is a Sec export chaperone, SecB, that binds preproteins, maintains them in an unfolded state, and delivers them to SecA – a central component of the Sec system discussed below. However, not all preproteins of Gram-negative bacteria require SecB, and Gram-positive bacteria lack a SecB ortholog (10). In the absence of SecB, it is thought that other proteins fulfill the role of export chaperone. In M. tuberculosis, Bordes et al. recently identified Rv1957 as a SecB-like protein (11). Rv1957 prevents proteolysis and aggregation of M.
4
However, there is currently no evidence that Rv1957 functions as a chaperone for protein export and it is possible that the function of Rv1957 is restricted to the HigBA system.
System components and mechanism. SecA is a multifunctional component of the Sec export system that works with the heterotrimeric SecYEG membrane channel to export proteins across the cytoplasmic membrane. SecA recognizes and binds to the signal peptide, as well as portions of the mature domain, of preproteins (12, 13). SecA is also an ATPase that harnesses energy from multiple rounds of ATP binding and hydrolysis to “push” preproteins through the SecYEG channel (14, 15). The proton motive force can also contribute to Sec export, but is not absolutely required (15).
Mycobacteria and some Gram-positive bacteria have two SecA proteins, each with distinct functions. The primary housekeeping SecA, which functions like SecA of E. coli, is referred to as SecA1, while the accessory SecA is referred to as SecA2 (see later section for SecA2 discussion). SecA1 is predicted to be essential in M. tuberculosis (16, 17) and proven to be essential in the model mycobacterium M. smegmatis (18). Because secA1 deletion mutants are not viable, conditional silencing has been used to study SecA1 function. As would be expected for a housekeeping SecA, depletion of SecA1 in M. smegmatis causes both growth inhibition and reduced export of a test protein with a Sec signal peptide (19, 20). M. tuberculosis SecA1 is also a demonstrated ATPase (21), and the X-ray crystal structure of M. tuberculosis SecA1 is similar to other bacterial SecA structures (22).
5
membrane through which proteins pass during export. SecE is suggested to act as a “clamp” to stabilize SecY (25). SecG improves efficiency of protein export (26). Additional Sec components that improve export efficiency are SecD, SecF, and YajC (27).
Once a protein has been exported across the membrane, the signal peptide is removed by one of two signal peptidases – the Type I signal peptidase LepB, or the Type II signal
peptidase LspA (28). LspA acts specifically on lipoprotein signal peptides. The M. tuberculosis
LepB (29) and LspA (30) have been studied and confirmed to function in protein
export. Furthermore, lepB is essential (29) and an lspA mutant of M. tuberculosis is attenuated in macrophage and mouse models of infection (30), demonstrating the importance of correctly exported proteins for M. tuberculosis survival and virulence.
In addition to the process described above for exporting proteins completely across the cytoplasmic membrane, the SecYEG channel is also used for a significantly different process – co-translational insertion of integral membrane proteins into the cytoplasmic membrane (for a recent review, see (31)). In this process, transmembrane domains of nascent integral
6
co-translational insertion of integral membrane proteins has only barely begun to receive research attention, with the recent confirmation that M. tuberculosis SRP displays biochemical properties comparable to SRP of E. coli (36).
Contribution to disease and physiology. Because the Sec system performs the bulk of protein export, many Sec-exported proteins are important for both M. tuberculosis viability and virulence. Several M. tuberculosis proteins proven to function in virulence contain Sec signal peptides (37, 38). Also, as mentioned above, the importance of LspA demonstrates that Sec-exported lipoproteins are important for M. tuberculosis virulence (30). In fact, several
lipoproteins with Sec signal peptides are known to be important for M. tuberculosis virulence, including LpqH (19kD) (39), LppX (40), and LprG (41). While SecYEG is homologous to the Sec61 complex found in eukaryotic cells, there is no eukaryotic homolog of SecA. The lack of SecA in mammalian cells, combined with the critical importance of SecA1 for both viability and virulence in bacteria, makes SecA1 a compelling potential drug target (as reviewed in (2)).
7
In the future it will be important to expand our understanding of the Sec export system in mycobacteria. For example, nothing is currently known about mycobacterial chaperone(s) involved in Sec export. Also, the process of inserting integral membrane proteins into the mycobacterial cytoplasmic membrane is a topic that has so far received little research attention.
Accessory SecA2 Export System
All mycobacteria are unusual in having two SecA proteins. In contrast to SecA1, SecA2 is non-essential and secA2 deletion mutants have been constructed in several mycobacteria, including M. tuberculosis (43), M. smegmatis (18), and Mycobacterium marinum (44). SecA2 functions in protein export, but its role appears limited to a much smaller subset of proteins than those exported by SecA1. SecA2 is necessary for the full virulence of M. tuberculosis (43, 45) and M. marinum (44), suggesting that one or more of the proteins it exports are important in pathogenesis. Accessory SecA2 export has been studied directly in mycobacteria; however, there is still much that is unknown.
Exported proteins and targeting. Comparative 2D-PAGE analysis of exported proteins from wild-type and secA2 deletion mutant strains has been used to identify proteins that require SecA2 for their export (43, 46). Only a small number of proteins were identified in these studies, and they include examples both with and without signal peptides. In M. smegmatis, cell wall proteins were analyzed by 2D-PAGE, identifying Msmeg_1704 and Msmeg_1712 as proteins exported by the SecA2 system (46). These proteins share many similarities – both are
8
although they have yet to be evaluated for SecA2-dependence. The SecA2-dependent proteins with signal peptides are currently indistinguishable from proteins exported by the housekeeping Sec export system, and experimental evidence shows the signal peptide is required for export of Msmeg_1712 (46). However, it is the mature domain of M. smegmatis SecA2-dependent proteins, not the signal peptide, that imparts the requirement for SecA2 in their export (47).
In M. tuberculosis, proteins exported into the culture media (culture filtrates) were analyzed by 2D-PAGE (43). Among the very few proteins identified was the antioxidant enzyme SodA (superoxide dismutase). SodA does not have a signal peptide, but its dependence on SecA2 for export was confirmed by western blot and SodA activity assay (43, 48). Given the role of SodA in detoxifying oxygen radicals, the export of M. tuberculosis KatG (catalase), another antioxidant enzyme, was directly evaluated for SecA2-dependence. Like SodA, KatG lacks a signal peptide and is dependent on SecA2 for export (43).
9
does not contain a SecY2 protein or any other known additional components. The mycobacterial SecA2 system is therefore referred to as a SecA2-only system.
SecA2 has been directly studied in both M. tuberculosis and M. smegmatis. Importantly, complementation experiments show that SecA2 of M. tuberculosis and M. smegmatis can substitute for one another to complement secA2 deletion mutant phenotypes (20). Because these cross-species complementation experiments demonstrate that the SecA2 proteins of M. tuberculosis and M. smegmatis can carry out the same functions, M. smegmatis has been used as a model for studying the mechanism of mycobacterial SecA2-dependent export.
SecA2 exhibits several similarities to the housekeeping SecA1 protein. SecA1 and SecA2 proteins are present in equivalent amounts in M. tuberculosis, and both proteins have proven ATPase activity (21). Like housekeeping SecA proteins, ATP hydrolysis is also required for SecA2 function in both M. tuberculosis (21) and M. smegmatis (20).
There are also several differences between SecA1 and SecA2. SecA2 is smaller than SecA1 because it lacks the C-terminal linker region, which in E. coli has been implicated in binding to phospholipids, SecB, and zinc (59-61). SecA2 also differs from SecA1 in how it localizes within the bacterium. Like E. coli SecA, SecA1 is evenly distributed between soluble and cell envelope fractions, but SecA2 is found predominantly in the cytosol-containing soluble fraction (20). Most importantly, it is clear that SecA1 and SecA2 have independent functions. Even when SecA2 is overexpressed, SecA1 cannot be deleted, indicating that SecA2 cannot substitute for SecA1. Similarly, overexpression of SecA1 does not rescue the phenotypes of a
secA2 deletion mutant (18).
10
housekeeping Sec export system. To address this possibility, export of the SecA2-dependent protein Msmeg_1712 was assessed following depletion of SecA1 in M. smegmatis. In the presence of SecA1 depletion, export of Msmeg_1712 was found to be significantly compromised (20). The simplest interpretation of this result is that the SecA2 export system needs the
housekeeping SecA1 to function. However, the possibility that SecA1 depletion has an indirect effect on SecA2 export cannot currently be ruled out.
Listeria monocytogenes, Clostridium difficile, and Corynebacterium glutamicum also have SecA2-only type export systems. A notable similarity between the L. monocytogenes and mycobacterial SecA2 systems is that both are reported to export proteins with signal peptides as well as proteins without signal peptides (62, 63). In L. monocytogenes, this includes export of MnSOD (manganese superoxide dismutase), which lacks a signal peptide (63). In addition, SecA2 of L. monocytogenes, like that of M. tuberculosis, is important for virulence but is not essential for growth (62, 64). Interestingly, L. monocytogenes SecA2 has been found to be enriched at cell division sites, and the polarly-localized cell division protein DivIVA is required for export of SecA2-dependent proteins p60 and MurA (65). It will be interesting to see if
mycobacterial SecA2 proteins also exhibit polar localization.
Unlike in mycobacteria, SecA2 of C. difficile and C. glutamicum are essential for growth (66, 67), which may reflect a difference in the essentiality of proteins exported by each of these systems. Despite this difference, there are also similarities between the mycobacterial and C. difficile SecA2 systems. In both species, SecA2 is found localized predominantly to the
11
these non-functional, dominant negative SecA2 KRvariants cause growth inhibition in both species, suggesting they interfere with an essential cell process. As discussed in chapter 2 of this dissertation, the essential process disrupted in M. smegmatis is believed to be the canonical Sec export pathway.
Contribution to disease and physiology. The M. tuberculosissecA2 mutant is
attenuated for growth in both macrophages (45) and the mouse model of tuberculosis infection (43). Similarly, the M. marinum secA2 mutant is attenuated for growth in both zebrafish and mouse models of infection (44). These results indicate that the SecA2 system exports proteins important for M. tuberculosis and M. marinum virulence. The fact that both M. tuberculosis
12
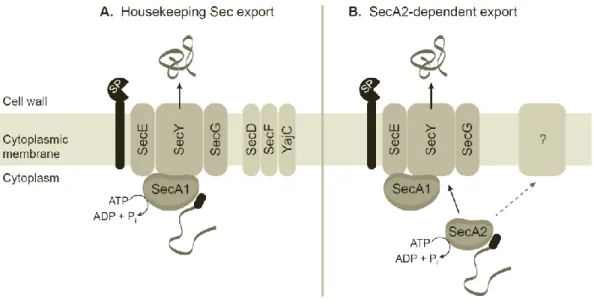
Model, summary, and future questions. More research is needed to elucidate the details of SecA2-dependent export. However the studies conducted prior to this dissertation, particularly those using M. smegmatis, help build a preliminary model for this system (Fig. 1.1). SecA2 recognizes a small subset of proteins that are normally not compatible with Sec export, and uses its ATPase activity to assist in their export. It seems most likely that SecA2 works with components of the housekeeping Sec export system, either by delivering proteins to SecA1 or by actively participating in exporting proteins through the SecYEG channel. It remains possible, however, that other unknown components are required in addition to or in lieu of the housekeeping Sec components.
13
Summary
While the canonical Sec export system has been studied extensively over several decades and is now well characterized, the SecA2 export system of mycobacteria is poorly understood. In addition to questions about the identity and targeting of SecA2-dependent proteins and the role of SecA2-dependent export in M. tuberculosis virulence, the mechanistic steps of SecA2 export remain unknown. In this dissertation, we describe a search for additional proteins that are either required for or connected to SecA2-dependent protein export. We demonstrate a relationship between SecA2 and the canonical Sec pathway, specifically the SecY channel protein. We also identify a genetic link between SecA2 and the conserved hypothetical protein Msmeg_1684. Finally, we demonstrate that the relationship between SecA2 and SecY does not appear to represent a direct, physical interaction.
In chapter 2, we describe a suppressor analysis carried out using the dominant-negative
M. smegmatis secA2 K129R allele. Interestingly, two extragenic suppressor strains carry mutations located upstream of the secY gene, encoding the main component of the canonical Sec channel complex. We demonstrate that these mutations are located within the secY
14
export (20). Depletion of SecA1 reduces export of a SecA2-dependent protein, but does not cause degradation of SecY, confirming the requirement for SecA1 in SecA2-dependent protein export.
In chapter 3, we extend our suppressor analysis of the M. smegmatis secA2 K129R
allele. Here, we identify several extragenic suppressors affecting the conserved hypothetical protein Msmeg_1684. We further show that an in-frame, unmarked deletion of msmeg_1684 is also able to suppress the secA2 K129R allele. The function of Msmeg_1684 is unknown.
However, the homologous Rv3311 protein of M. tuberculosis, like SecA2, is predicted by TraSH to be important for growth in macrophages (69). Intriguingly, Msmeg_1684-like proteins and SecA2 proteins frequently co-occur within the order Actinomycetales, of which mycobacteria are members. Our findings suggest that Msmeg_1684 is required for the interaction of SecA2 with the housekeeping Sec pathway and may participate directly in SecA2-dependent protein export.
15
Our results suggest a model in which SecA2-dependent protein export is integrated into the canonical Sec pathway. SecA1 is required for dependent export, and
16
Table 1.1. M. smegmatis and M. tuberculosis homologs of conserved export system components.
Export system components
Required for
E. coli growtha M. smegmatis homolog M. tuberculosis homolog
Required for
M. tuberculosis growthb
SecA yes Msmeg_1881 Rv3240c (SecA1) yes
Msmeg_3654 Rv1821 (SecA2) no*† (43)
SecY yes Msmeg_1483 Rv0732 yes
SecE yes Msmeg_1344 Rv0638 yes
SecG no Msmeg_3087 Rv1440 no
SecD no Msmeg_2961 Rv2587c yes
SecF no Msmeg_2962 Rv2586c yes
YajC no Msmeg_2960 Rv2588c no
SecB no Rv1957‡ no
Ffh (SRP) yes Msmeg_2430 Rv2916c yes
FtsY yes Msmeg_2424 Rv2921c yes
YidC yes Msmeg_6942 Rv3921c yes
LepB yes Msmeg_2441 Rv2903c yes* (29)
LspA yes Msmeg_3174 Rv1539 no*† (30)
a
information compiled from references (70-78)
b
unless marked by *, requirement for growth is predicted by deep sequencing of transposon libraries (Tn-seq) (17) * experimentally demonstrated
17
Figure 1.1. A preliminary model of the mycobacterial housekeeping Sec and SecA2-dependent protein export systems. (A) Preproteins (grey ribbon) with N-terminal signal
peptides (black oval) are recognized by SecA1, which interacts with the SecYEG channel complex to form the translocase. SecA1 performs repeated cycles of ATP hydrolysis, pushing the
18
References
1. 2012. Global Tuberculosis Report 2012. World Health Organization.
2. Feltcher ME, Sullivan JT, Braunstein M. 2010. Protein export systems of Mycobacterium tuberculosis: novel targets for drug development? Future Microbiol 5:1581-1597. 3. Finlay BB, Falkow S. 1997. Common themes in microbial pathogenicity revisited.
Microbiol Mol Biol Rev 61:136-169.
4. Papanikou E, Karamanou S, Economou A. 2007. Bacterial protein secretion through the translocase nanomachine. Nat Rev Microbiol 5:839-851.
5. Driessen AJ, Nouwen N. 2008. Protein translocation across the bacterial cytoplasmic membrane. Annu Rev Biochem 77:643-667.
6. Braun V, Wu HC. 1994. Lipoproteins, structure, function, biosynthesis and models for protein export., p. 319-342. In Ghuysen JM, Hackenbeck R (ed.), Bacterial cell wall. Elsevier, Amsterdam; New York.
7. Yu NY, Wagner JR, Laird MR, Melli G, Rey S, Lo R, Dao P, Sahinalp SC, Ester M, Foster LJ, Brinkman FS. 2010. PSORTb 3.0: improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 26:1608-1615.
8. Petersen TN, Brunak S, von Heijne G, Nielsen H. 2011. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat Methods 8:785-786.
9. Juncker AS, Willenbrock H, Von Heijne G, Brunak S, Nielsen H, Krogh A. 2003.
Prediction of lipoprotein signal peptides in Gram-negative bacteria. Protein Sci 12: 1652-1662.
10. van der Sluis EO, Driessen AJ. 2006. Stepwise evolution of the Sec machinery in
Proteobacteria. Trends Microbiol 14:105-108.
11. Bordes P, Cirinesi AM, Ummels R, Sala A, Sakr S, Bitter W, Genevaux P. 2011. SecB-like chaperone controls a toxin-antitoxin stress-responsive system in Mycobacterium tuberculosis. Proc Natl Acad Sci U S A 108:8438-8443.
12. Cunningham K, Wickner W. 1989. Specific recognition of the leader region of precursor proteins is required for the activation of translocation ATPase of Escherichia coli. Proc Natl Acad Sci U S A 86:8630-8634.
13. Lill R, Dowhan W, Wickner W. 1990. The ATPase activity of SecA is regulated by acidic phospholipids, SecY, and the leader and mature domains of precursor proteins. Cell
19
14. van der Wolk JP, de Wit JG, Driessen AJ. 1997. The catalytic cycle of the escherichia coli
SecA ATPase comprises two distinct preprotein translocation events. EMBO J 16: 7297-7304.
15. Schiebel E, Driessen AJ, Hartl FU, Wickner W. 1991. Delta mu H+ and ATP function at different steps of the catalytic cycle of preprotein translocase. Cell 64:927-939.
16. Sassetti CM, Boyd DH, Rubin EJ. 2003. Genes required for mycobacterial growth defined by high density mutagenesis. Mol Microbiol 48:77-84.
17. Griffin JE, Gawronski JD, Dejesus MA, Ioerger TR, Akerley BJ, Sassetti CM. 2011. High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PLoS Pathog 7:e1002251.
18. Braunstein M, Brown AM, Kurtz S, Jacobs WR, Jr. 2001. Two nonredundant SecA homologues function in mycobacteria. J Bacteriol 183:6979-6990.
19. Guo XV, Monteleone M, Klotzsche M, Kamionka A, Hillen W, Braunstein M, Ehrt S, Schnappinger D. 2007. Silencing Mycobacterium smegmatis by using tetracycline repressors. J Bacteriol 189:4614-4623.
20. Rigel NW, Gibbons HS, McCann JR, McDonough JA, Kurtz S, Braunstein M. 2009. The Accessory SecA2 System of Mycobacteria Requires ATP Binding and the Canonical SecA1. J Biol Chem 284:9927-9936.
21. Hou JM, D'Lima NG, Rigel NW, Gibbons HS, McCann JR, Braunstein M, Teschke CM.
2008. ATPase activity of Mycobacterium tuberculosis SecA1 and SecA2 proteins and its importance for SecA2 function in macrophages. J Bacteriol 190:4880-4887.
22. Sharma V, Arockiasamy A, Ronning DR, Savva CG, Holzenburg A, Braunstein M, Jacobs WR, Jr., Sacchettini JC. 2003. Crystal structure of Mycobacterium tuberculosis SecA, a preprotein translocating ATPase. Proc Natl Acad Sci U S A 100:2243-2248.
23. Brundage L, Hendrick JP, Schiebel E, Driessen AJ, Wickner W. 1990. The purified E. coli
integral membrane protein SecY/E is sufficient for reconstitution of SecA-dependent precursor protein translocation. Cell 62:649-657.
24. Hanada M, Nishiyama KI, Mizushima S, Tokuda H. 1994. Reconstitution of an efficient protein translocation machinery comprising SecA and the three membrane proteins, SecY, SecE, and SecG (p12). J Biol Chem 269:23625-23631.
25. Van den Berg B, Clemons WM, Jr., Collinson I, Modis Y, Hartmann E, Harrison SC, Rapoport TA. 2004. X-ray structure of a protein-conducting channel. Nature 427:36-44. 26. Flower AM, Hines LL, Pfennig PL. 2000. SecG is an auxiliary component of the protein
export apparatus of Escherichia coli. Mol Gen Genet 263:131-136.
20
28. Paetzel M, Karla A, Strynadka NC, Dalbey RE. 2002. Signal peptidases. Chem Rev
102:4549-4580.
29. Ollinger J, O'Malley T, Ahn J, Odingo J, Parish T. 2012. Inhibition of the sole type I signal peptidase of Mycobacterium tuberculosis is bactericidal under replicating and
nonreplicating conditions. J Bacteriol 194:2614-2619.
30. Sander P, Rezwan M, Walker B, Rampini SK, Kroppenstedt RM, Ehlers S, Keller C, Keeble JR, Hagemeier M, Colston MJ, Springer B, Bottger EC. 2004. Lipoprotein processing is required for virulence of Mycobacterium tuberculosis. Mol Microbiol
52:1543-1552.
31. Facey SJ, Kuhn A. 2010. Biogenesis of bacterial inner-membrane proteins. Cell Mol Life Sci 67:2343-2362.
32. Egea PF, Stroud RM. 2010. Lateral opening of a translocon upon entry of protein suggests the mechanism of insertion into membranes. Proc Natl Acad Sci U S A
107:17182-17187.
33. Dalbey RE, Kuhn A. 2004. YidC family members are involved in the membrane insertion, lateral integration, folding, and assembly of membrane proteins. J Cell Biol 166:769-774. 34. Dalbey RE, Chen M. 2004. Sec-translocase mediated membrane protein biogenesis.
Biochim Biophys Acta 1694:37-53.
35. Sonnhammer EL, von Heijne G, Krogh A. 1998. A hidden Markov model for predicting transmembrane helices in protein sequences. Proc Int Conf Intell Syst Mol Biol 6: 175-182.
36. Palaniyandi K, Veerasamy M, Narayanan S. 2012. Characterization of Ffh of
Mycobacterium tuberculosis and its interaction with 4.5S RNA. Microbiological research
167:520-525.
37. McCann JR, Kurtz S, Braunstein M. 2009. Secreted and Exported Proteins Important to
Mycobacterium tuberculosis Pathogenesis, p. 265-297. In Wooldridge K (ed.), Bacterial secreted proteins : secretory mechanisms and role in pathogenesis. Caister Academic Press, Wymondham.
38. McCann JR, McDonough JA, Sullivan JT, Feltcher ME, Braunstein M. 2011. Genome-wide identification of Mycobacterium tuberculosis exported proteins with roles in intracellular growth. J Bacteriol 193:854-861.
39. Henao-Tamayo M, Junqueira-Kipnis AP, Ordway D, Gonzales-Juarrero M, Stewart GR, Young DB, Wilkinson RJ, Basaraba RJ, Orme IM. 2007. A mutant of Mycobacterium tuberculosis lacking the 19-kDa lipoprotein Rv3763 is highly attenuated in vivo but retains potent vaccinogenic properties. Vaccine 25:7153-7159.
21
2006. LppX is a lipoprotein required for the translocation of phthiocerol
dimycocerosates to the surface of Mycobacterium tuberculosis. EMBO J 25:1436-1444. 41. Bigi F, Gioffre A, Klepp L, Santangelo MP, Alito A, Caimi K, Meikle V, Zumarraga M,
Taboga O, Romano MI, Cataldi A. 2004. The knockout of the lprG-Rv1410 operon produces strong attenuation of Mycobacterium tuberculosis. Microbes Infect 6:182-187. 42. Owens MU, Swords WE, Schmidt MG, King CH, Quinn FD. 2002. Cloning, expression,
and functional characterization of the Mycobacterium tuberculosis secA gene. FEMS Microbiol Lett 211:133-141.
43. Braunstein M, Espinosa BJ, Chan J, Belisle JT, Jacobs WR, Jr. 2003. SecA2 functions in the secretion of superoxide dismutase A and in the virulence of Mycobacterium tuberculosis. Mol Microbiol 48:453-464.
44. Watkins BY, Joshi SA, Ball DA, Leggett H, Park S, Kim J, Austin CD, Paler-Martinez A, Xu M, Downing KH, Brown EJ. 2012. Mycobacterium marinum SecA2 promotes stable granulomas and induces tumor necrosis factor alpha in vivo. Infect Immun 80: 3512-3520.
45. Kurtz S, McKinnon KP, Runge MS, Ting JP, Braunstein M. 2006. The SecA2 secretion factor of Mycobacterium tuberculosis promotes growth in macrophages and inhibits the host immune response. Infect Immun 74:6855-6864.
46. Gibbons HS, Wolschendorf F, Abshire M, Niederweis M, Braunstein M. 2007. Identification of two Mycobacterium smegmatis lipoproteins exported by a SecA2-dependent pathway. J Bacteriol 189:5090-5100.
47. Feltcher ME, Gibbons HS, Ligon LS, Braunstein M. 2013. Protein export by the mycobacterial SecA2 system is determined by the preprotein mature domain. J. Bacteriol. 195:672-681.
48. Hinchey J, Lee S, Jeon BY, Basaraba RJ, Venkataswamy MM, Chen B, Chan J, Braunstein M, Orme IM, Derrick SC, Morris SL, Jacobs WR, Jr., Porcelli SA. 2007. Enhanced priming of adaptive immunity by a proapoptotic mutant of Mycobacterium tuberculosis. J Clin Invest 117:2279-2288.
49. Feltcher ME, Braunstein M. 2012. Emerging themes in SecA2-mediated protein export. Nat. Rev. Microbiol. 10:779-789.
50. Bensing BA, Sullam PM. 2002. An accessory sec locus of Streptococcus gordonii is required for export of the surface protein GspB and for normal levels of binding to human platelets. Mol Microbiol 44:1081-1094.
51. Chen Q, Wu H, Fives-Taylor PM. 2004. Investigating the role of secA2 in secretion and glycosylation of a fimbrial adhesin in Streptococcus parasanguis FW213. Mol Microbiol
22
52. Siboo IR, Chaffin DO, Rubens CE, Sullam PM. 2008. Characterization of the accessory Sec system of Staphylococcus aureus. J Bacteriol 190:6188-6196.
53. Takamatsu D, Bensing BA, Sullam PM. 2004. Genes in the accessory sec locus of Streptococcus gordonii have three functionally distinct effects on the expression of the platelet-binding protein GspB. Mol Microbiol 52:189-203.
54. Takamatsu D, Bensing BA, Sullam PM. 2005. Two additional components of the accessory sec system mediating export of the Streptococcus gordonii platelet-binding protein GspB. J Bacteriol 187:3878-3883.
55. Bensing BA, Gibson BW, Sullam PM. 2004. The Streptococcus gordonii platelet binding protein GspB undergoes glycosylation independently of export. J Bacteriol 186:638-645. 56. Mistou MY, Dramsi S, Brega S, Poyart C, Trieu-Cuot P. 2009. Molecular dissection of the
secA2 locus of group B Streptococcus reveals that glycosylation of the Srr1 LPXTG protein is required for full virulence. J Bacteriol 191:4195-4206.
57. Wu H, Zeng M, Fives-Taylor P. 2007. The glycan moieties and the N-terminal
polypeptide backbone of a fimbria-associated adhesin, Fap1, play distinct roles in the biofilm development of Streptococcus parasanguinis. Infect Immun 75:2181-2188. 58. Zhou M, Zhu F, Dong S, Pritchard DG, Wu H. 2010. A novel glucosyltransferase is
required for glycosylation of a serine-rich adhesin and biofilm formation by Streptococcus parasanguinis. J Biol Chem 285:12140-12148.
59. Breukink E, Nouwen N, van Raalte A, Mizushima S, Tommassen J, de Kruijff B. 1995. The C terminus of SecA is involved in both lipid binding and SecB binding. J Biol Chem
270:7902-7907.
60. Fekkes P, van der Does C, Driessen AJ. 1997. The molecular chaperone SecB is released from the carboxy-terminus of SecA during initiation of precursor protein translocation. EMBO J 16:6105-6113.
61. Dempsey BR, Wrona M, Moulin JM, Gloor GB, Jalilehvand F, Lajoie G, Shaw GS, Shilton BH. 2004. Solution NMR structure and X-ray absorption analysis of the C-terminal zinc-binding domain of the SecA ATPase. Biochemistry 43:9361-9371.
62. Lenz LL, Mohammadi S, Geissler A, Portnoy DA. 2003. SecA2-dependent secretion of autolytic enzymes promotes Listeria monocytogenes pathogenesis. Proc Natl Acad Sci U S A 100:12432-12437.
63. Archambaud C, Nahori MA, Pizarro-Cerda J, Cossart P, Dussurget O. 2006. Control of
Listeria superoxide dismutase by phosphorylation. J Biol Chem 281:31812-31822. 64. Lenz LL, Portnoy DA. 2002. Identification of a second Listeria secA gene associated with
23
65. Halbedel S, Hahn B, Daniel RA, Flieger A. 2012. DivIVA affects secretion of virulence-related autolysins in Listeria monocytogenes. Mol Microbiol 83:821-839.
66. Fagan RP, Fairweather NF. 2011. Clostridium difficile has two parallel and essential Sec secretion systems. J. Biol. Chem. 286:27483-27493.
67. Caspers M, Freudl R. 2008. Corynebacterium glutamicum possesses two secA
homologous genes that are essential for viability. Archives of microbiology 189:605-610. 68. Sullivan JT, Young EF, McCann JR, Braunstein M. 2012. The Mycobacterium tuberculosis
SecA2 system subverts phagosome maturation to promote growth in macrophages. Infect Immun 80:996-1006.
69. Rengarajan J, Bloom BR, Rubin EJ. 2005. Genome-wide requirements for
Mycobacterium tuberculosis adaptation and survival in macrophages. Proc Natl Acad Sci U S A 102:8327-8332.
70. Murphy CK, Beckwith J. 1996. Export of Proteins to the Cell Envelope in Escherichia coli, p. 967-978. In Neidhardt FC (ed.), Escherichia coli and Salmonella : cellular and
molecular biology, 2nd ed. ASM Press, Washington, D.C.
71. Shiba K, Ito K, Yura T, Cerretti DP. 1984. A defined mutation in the protein export gene within the spc ribosomal protein operon of Escherichia coli: isolation and
characterization of a new temperature-sensitive secY mutant. EMBO J 3:631-635. 72. Pogliano KJ, Beckwith J. 1994. Genetic and molecular characterization of the Escherichia
coli secD operon and its products. J Bacteriol 176:804-814.
73. Phillips GJ, Silhavy TJ. 1992. The E. coli ffh gene is necessary for viability and efficient protein export. Nature 359:744-746.
74. Gill DR, Salmond GP. 1990. The identification of the Escherichia coli ftsY gene product: an unusual protein. Mol Microbiol 4:575-583.
75. van Bloois E, Nagamori S, Koningstein G, Ullers RS, Preuss M, Oudega B, Harms N, Kaback HR, Herrmann JM, Luirink J. 2005. The Sec-independent function of Escherichia coli YidC is evolutionary-conserved and essential. J Biol Chem 280:12996-13003.
76. Bogsch EG, Sargent F, Stanley NR, Berks BC, Robinson C, Palmer T. 1998. An essential component of a novel bacterial protein export system with homologues in plastids and mitochondria. J Biol Chem 273:18003-18006.
77. Sargent F, Bogsch EG, Stanley NR, Wexler M, Robinson C, Berks BC, Palmer T. 1998. Overlapping functions of components of a bacterial Sec-independent protein export pathway. EMBO J 17:3640-3650.
78. Sargent F, Stanley NR, Berks BC, Palmer T. 1999. Sec-independent protein translocation in Escherichia coli. A distinct and pivotal role for the TatB protein. J Biol Chem
CHAPTER 2
Suppressor Analysis Reveals a Role for SecY in the SecA2-Dependent
Protein Export Pathway of Mycobacteria
1All bacteria use the conserved Sec pathway to transport proteins across the cytoplasmic membrane, with the SecA ATPase playing a central role in the process. Mycobacteria are part of a small group of bacteria that have two SecA proteins: the canonical SecA (SecA1) and a second, specialized SecA (SecA2). The SecA2-dependent pathway exports a small subset of proteins and is required for Mycobacterium tuberculosis virulence. The mechanism by which SecA2 drives export of proteins across the cytoplasmic membrane remains poorly understood. Here we performed suppressor analysis on a dominant negative secA2 mutant (secA2 K129R) of the model mycobacterium Mycobacterium smegmatis to better understand the pathway used by SecA2 to export proteins. Two extragenic suppressor mutations were identified as mapping to the promoter region of secY, which encodes the central component of the canonical Sec export channel. These suppressor mutations increased secY expression, and this effect was sufficient to alleviate secA2 K129R phenotypes. We also discovered that the level of SecY protein was greatly diminished in the secA2 K129R mutant, but at least partially restored in thesuppressors. Furthermore, the level of SecY in a suppressor strongly correlated with the degree of
1
25
suppression. Our findings reveal a detrimental effect of SecA2 K129R on SecY, arguing for an integrated system in which SecA2 works with SecY and the canonical Sec translocase to export proteins.
Introduction
Bacteria use both conserved and specialized protein export systems to deliver proteins to the bacterial cell surface and to the extracellular environment. These exported proteins are important for critical cell functions like nutrient acquisition, cell structure, and in the case of pathogens, virulence. All bacteria possess the conserved Sec protein export system, which performs the bulk of “housekeeping” protein export. More recently, it was discovered that mycobacteria and some Gram-positive bacteria possess a second, specialized Sec export system (1-7). This specialized protein export system is only partially understood, but in pathogens is often required for virulence.
26
In mycobacteria, SecA2 is responsible for the export of a small subset of proteins (15, 16) and is required for virulence of the pathogen Mycobacterium tuberculosis (16). The model organism Mycobacterium smegmatis also possesses a SecA2 system, which is functionally conserved with that of M. tuberculosis (6, 17). Studies with M. smegmatis show that SecA2-dependent proteins can contain N-terminal signal peptides indistinguishable from classical Sec signal peptides and that it is the mature domain of the protein that determines the requirement for SecA2 (18). However, the mechanism of SecA2-dependent protein export in mycobacteria is unknown. Some SecA2 export systems, referred to as “SecA2-SecY2” systems, include a second distinct SecY protein (named SecY2) that is required for export of SecA2-dependent proteins. In these systems, SecY2 is presumed to function as the channel for protein translocation across the membrane (5, 7, 14). In mycobacteria, there is no SecY2 protein evident, making it unclear how SecA2-dependent proteins are exported in this system. One possibility is that the mycobacterial SecA2 system exports proteins through the SecYEG channel, sharing this channel with the housekeeping Sec export system; however, this model has not been proven.
To increase our understanding of SecA2 export and identify additional components of the SecA2 system, we carried out a suppressor analysis of a secA2 mutant of M. smegmatis. For this purpose, we used a secA2 K129R mutant that encodes for a SecA2 protein with an amino acid substitution (K129R) in the Walker box, which is an amino acid motif important for ATP binding and hydrolysis. We previously demonstrated that this K129R substitution disrupts the ATPase activity of SecA2, rendering it non-functional (12, 17). In addition, SecA2 K129R has a dominant negative effect on wild type SecA2 (17). This suggests that SecA2 K129R is still able to interact with its normal binding partners within the cell, but because it is non-functional,
27
secA2 deletion mutant. These properties make the secA2 K129R allele a good starting point for suppressor analysis.
In the present study, we characterized two extragenic suppressor mutations of secA2 K129R, both of which are located in the promoter region of the only secY gene of mycobacteria. We also discovered that SecY protein levels are drastically reduced in the secA2 K129R mutant, and that these suppressor mutations increase secY expression to partially restore SecY protein levels and suppress secA2 K129R phenotypes. Finally, we found that SecY levels were increased relative to the secA2 K129R strain in six additional extragenic suppressors. Taken together, these findings suggest that SecA2 K129R disrupts the housekeeping Sec export system, causing SecY degradation and thus the severe phenotypes of the secA2 K129R mutant. The data presented here argues for SecA2 working in concert with SecY and the housekeeping Sec pathway to export SecA2-dependent proteins.
Materials and Methods
Bacterial strains and culture conditions. M. smegmatis strains used in this study are described in Table 2.1, and were grown at 37°C or 30°C, using Middlebrook 7H9/7H10 or Mueller-Hinton media. To limit acquisition of suppressors when working with the secA2 K129R
strain (NR178), starter cultures were generally grown at 30°C, but the actual experiments performed at 37°C. Media were supplemented with 0.5% glycerol plus 0.2% glucose (7H9/7H10 media only) and 0.1% tween-80 (all media). Antibiotics kanamycin (20 μg/mL) and hygromycin B (50 μg/mL) were added as needed. When required, plasmids were introduced into M.
28
lysogeny broth or on lysogeny broth agar (otherwise known as Luria-Bertani media). Antibiotics kanamycin (40 μg/mL) and hygromycin B (150 μg/mL) were added as needed.
Suppressor collection. The secA2 K129R strain encodes for a SecA2 protein in which the lysine at position 129 is substituted with an arginine (amino acid numbering based on NCBI GenBank accession number AF287049 (6)). Suppressors of secA2 K129R were isolated by plating 38 independently grown cultures of the secA2 K129R strain onto Mueller-Hinton agar at 37°C. Spontaneous suppressors (i.e. colonies that grew on Mueller-Hinton agar) were obtained, and one small (S) and one large (B) suppressor colony was chosen from each independent culture.
Plasmid construction. Plasmids used in this study are described in Table 2.2 and
primers are described in Table 2.3. In all cases, newly constructed plasmids were sequenced. To create secY’-‘lacZ fusion plasmids, a region upstream of secY was PCR amplified from M.
smegmatis genomic DNA of strains NR178 and NR236 (suppressor 4S) to generate wild type and 4S secY promoter sequences, respectively. Each PCR product contained 379 bp upstream of
secY along with 34 bp of secY coding sequence and engineered EcoRI restriction sites, and was cloned into pCR2.1-TOPO (Invitrogen), yielding plasmids pLL5 and pLL6. The EcoRI secY
promoter fragments were cut from pLL5 and pLL6 and ligated into EcoRI-digested pCV125, yielding plasmids pLL11 and pLL8, which contain secY’-‘lacZ translational fusions. pLL11 was mutated by site-directed mutagenesis (Stratagene QuikChange II) to re-create the mutation found upstream of secY in suppressor NR151 (24S), yielding plasmid pLL15.
29
genomic DNA, with engineered NotI and EcoRV restriction sites for cloning purposes. Each PCR product was cloned into pCR2.1-TOPO (Invitrogen), yielding plasmids pLL17 and pLL19. The NotI-EcoRV secY fragments were cut from pLL17 and pLL19 and ligated into NotI-EcoRV-digested pLL2, yielding plasmids pLL21 and pLL23.
Azide sensitivity assays. Cultures were plated for azide sensitivity as previously
described (17), by mixing 200 µL of a saturated culture with 7H9 top agar and pouring over 7H10 agar plates (with tween supplementation omitted) in three technical replicates. Discs soaked with 10 µL of 0.15 M sodium azide were added to the center of each plate. The diameter of the zone of inhibition was measured after two days and presented as a percentage of the entire plate diameter, yielding percent azide inhibition.
Whole genome and directed sequencing. Genomic DNA was isolated (as described previously (21)) from the 4S suppressor strain (NR236) and submitted for whole genome sequencing at the High-Throughput Sequencing Facility at the University of North Carolina at Chapel Hill. Sequencing was performed using Illumina GA II technology with 36 bp single-end reads. Reads were aligned to the M. smegmatis mc2155 reference genome (NCBI RefSeq accession number NC_008596.1) using SOAP (22) (with default parameters), resulting in an average sequence coverage of 47.5. Single nucleotide polymorphisms, insertions, and deletions were located using SOAP and BLAT (23) (default parameters), resulting in a total of 86 mutations identified. Mutations also identified in other mc2155-derived strains sequenced in our
30
5’ RLM-RACE. RNA ligase-mediated rapid amplification of cDNA ends (RLM-RACE) was performed using the GeneRacer kit (Invitrogen) according to manufacturer instructions. Briefly, total RNA was isolated from strain NR172, treated with TAP to remove 5’ triphosphates, and ligated to the GeneRacer RNA oligo. secY 5’ ends were reverse-transcribed to cDNA using a gene-specific primer located within the secY coding sequence (primer R1, see Fig. 2.2B). 5’ end cDNA was then amplified using a nested PCR strategy involving a first round of PCR with a second gene-specific primer located within the secY coding sequence (primer R2) and the GeneRacer 5’ primer (homologous to the GeneRacer RNA oligo), followed by a second round of PCR with a primer located upstream of the secY coding sequence (primer R3) and the GeneRacer 5’ primer. PCR products were separated by agarose gel electrophoresis, individually gel purified, and cloned into pCR2.1-TOPO (Invitrogen). A minimum of 4 clones originating from each PCR product were submitted to Genewiz, Inc. for sequencing. Transcript 5’ ends were identified as the first nucleotide following the sequence of the GeneRacer RNA oligo. Nucleotide positions were numbered relative to the translational start site of secY (NCBI GenBank accession number ABK75688).
Quantitative RT-PCR. Strains were grown in 7H9 broth to an OD600 nm of approximately
1.0, pelleted by centrifugation at 1,600 x g for 10 minutes, and flash-frozen. RNA was isolated using one of two protocols. For both protocols, bacteria were lysed in 1 mL 3:1
31
(Promega SV Total RNA Isolation System). All RNA samples were treated with DNAse (Promega) and then column-purified (Zymo DNA-Free RNA kit). Following RNA isolation, quantitative RT-PCR was performed in triplicate technical replicates using 25 or 50 ng RNA in each reaction. Products were reverse transcribed and amplified from total RNA using the Bioline SensiMix SYBR & Fluorescein One-Step Kit, and amplified from a DNA standard using the Bioline SensiMix SYBR & Fluorescein Kit. For quantitative RT-PCR on secY, products were amplified from the 5’ ends of
secY and rpoB gene sequences, starting quantity of each transcript was calculated relative to DNA standards, and secY transcripts were normalized to rpoB transcripts (as a control) in each sample (24).
LacZ (β-galactosidase) activity assays. LacZ activity assays were performed similar to those described previously (25). Each strain was grown in 7H9 broth to late-exponential or saturated phase and 800 μL was pelleted in a microcentrifuge. Cell pellets were resuspended in 800 μL Z buffer (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, 50 mM
β-mercaptoethanol), then lysed with 35 μL chloroform and 1 μL 0.1% SDS, and vortexed for 30 seconds. 160 μL ONPG (4 mg/mL in Z buffer) was added to each reaction and incubated for 1 hour at room temperature. Reactions were terminated by addition of 400 μL of 1 M Na2CO3.
Debris was removed by centrifugation at 16,000 x g for 3 minutes, and the OD420 nm was read
from the supernatant. LacZ activity (Miller units) was calculated by the formula: (1,000 x OD420
nm) / (reaction time (minutes) x culture volume used in reaction (mL) x OD600 nm).
Subcellular fractionation and western blotting. Each strain was grown in Mueller-Hinton broth to an OD600 nm of approximately 1.0. Subcellular fractions were prepared as
32
lysates (supernatants) were standardized for equal protein content using the Pierce BCA Protein Assay. Clarified whole cell lysates were centrifuged at 100,000 x g for 2 hours to separate cell envelope (pellet) and soluble (supernatant) fractions. Alternately, clarified whole cell lysates were centrifuged at 50,000 x g for 30 minutes to separate cell wall (pellet) and the resulting supernatant was centrifuged at 100,000 x g for 2 hours to separate cell membrane (pellet) and soluble (supernatant) fractions. In some cases, 1% Igepal CA-630 was added during cell envelope resuspension to facilitate solubilization of SecY.
Whole cell lysates (WCL, loaded for equal protein content) or subcellular fractions (loaded for equivalent starting cell material to WCL samples) were boiled, separated by SDS-PAGE, and transferred to nitrocellulose membranes for western blotting. Proteins were detected using a rabbit αSecY polyclonal antiserum at a 1:150 dilution, a rabbit αSecA1 polyclonal antiserum at a 1:50,000 dilution (26), and a mouse αHA monoclonal antibody at a 1:10,000 dilution (Covance). Primary antibodies were detected using alkaline phosphatase-conjugated goat anti-rabbit or goat anti-mouse IgG secondary antibodies, ECF substrate (GE Healthcare), and a Molecular Dynamics Storm 860 phosphorimager or Syngene G:BOX machine. To measure localization of Msmeg_1712-HA in subcellular fractions, fluorescence was quantified using GeneTools software (Syngene) and the percent located in each fraction was calculated relative to the total fluorescence of cell wall + cell membrane + soluble fractions.
33
Construction of SecA1 depletion strain. The SecA1 depletion strain, JM693, was created as described previously (26). Briefly, wild type M. smegmatis mc2155 was transformed with the pKIsecA1 suicide plasmid (gift of Dr. Sabine Ehrt, Weill Cornell Medical College) and transformants were selected by plating on media containing hygromycin B. Integration of pKIsecA1 at the native secA1 locus in the chromosome was confirmed by southern blot.
Regulation of SecA1 in strain JM693 was achieved by introducing plasmid pNR55 (expressing the revTetR repressor) and growing cultures in the dark with anhydrotetracycline (1200 ng/mL) supplementation. This level of anhydrotetracycline was sufficient to completely deplete SecA1 protein levels (as assessed by western blotting) without impacting cell growth.
Results
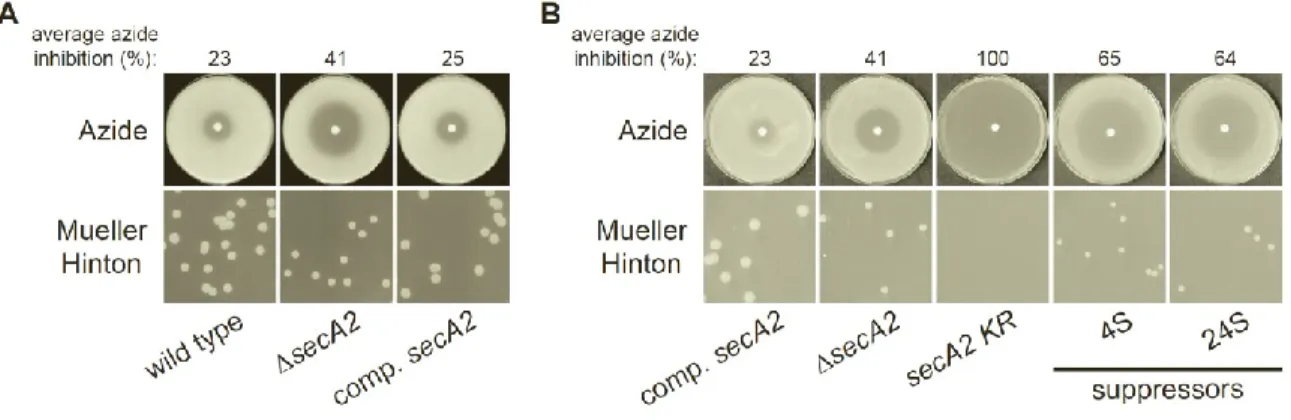
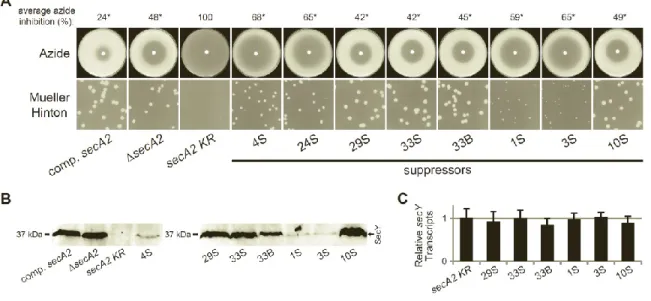
Spontaneous suppressor mutations alleviate the severe phenotypes of the secA2 K129R allele. Compared to wild type M. smegmatis, a secA2 deletion mutant exhibits a moderate growth defect on Mueller-Hinton agar and increased sensitivity to azide (6). This phenotype is complemented by adding the wild type secA2 allele, expressed from the constitutive hsp60 promoter, into the secA2 deletion mutant (Fig. 2.1A) (17). However,
introduction of the secA2 K129R allele, also expressed from the hsp60 promoter, into the secA2
deletion mutant fails to complement secA2 deletion phenotypes. SecA2 K129R is non-functional as a result of an amino acid substitution in the ATP binding Walker box. Moreover, the secA2 K129R allele exacerbates both the growth defect on Mueller-Hinton agar and sensitivity to azide phenotypes (i.e. secA2 K129R phenotypes are more severe than phenotypes of the secA2
34
Suppressor mutations spontaneously arise in the secA2 K129R background, and are easily identified by their ability to grow on Mueller-Hinton agar. To begin our suppressor analysis, we collected 63 suppressor mutants. All suppressors alleviated the severe phenotypes associated with the secA2 K129R allele, improving both growth on Mueller-Hinton agar and resistance to azide (Fig. 2.1B). Throughout this study, we routinely compared suppressor strains to the complemented secA2 strain (NR172) as a control, instead of the wild type strain. By doing so, we could study the effect of the suppressor mutations across a series of strains that all express secA2 alleles from the same hsp60 promoter. In no case did suppressors rescue phenotypes to the level of the complemented secA2 strain. Instead, suppressor phenotypes more closely resembled the phenotypes of the secA2 deletion strain, suggesting that suppressor mutations overcome the detrimental effect of SecA2 K129R, but do not necessarily restore SecA2-dependent export. Of the 63 suppressors isolated, 40 did not produce SecA2 K129R protein (by western blot, data not shown), suggesting that these suppressors alleviate secA2 K129R phenotypes by preventing SecA2 K129R production. This is a category of suppressors that we expected to observe. The remaining 23 suppressors produced detectable levels of full-length SecA2 K129R protein.
We next sequenced the secA2 K129R allele in the 23 suppressors producing SecA2 K129R protein. Of these 23 suppressors, 9 strains contained mutations in the secA2 K129R allele (intragenic), while the remaining 14 strains had no mutations in the secA2 K129R allele and are therefore extragenic suppressors. Because our goal was to identify proteins that work with SecA2 during export, we focused on characterizing extragenic suppressors.
35
and performed whole genome sequencing. Alignment of sequence reads to the M. smegmatis
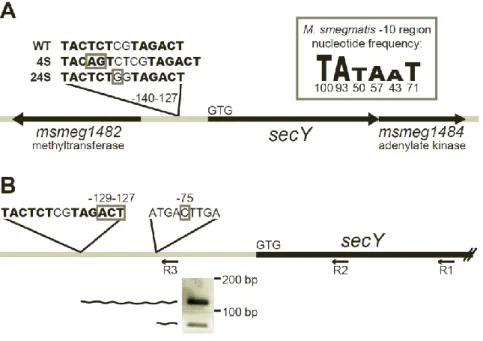
mc2155 reference genomeresulted in average sequence coverage of 47.5, sufficient to identify mutations in the 4S genome. Each candidate suppressor mutation was PCR amplified and re-sequenced from both 4S and its parent secA2 K129R strain. Following this directed sequencing, false positives and background mutations were discarded, leaving only a single mutation uniquely present in suppressor 4S. This unique mutation was a 2 bp insertion located 137 bp upstream of the translational start site for the predicted M. smegmatissecY gene, msmeg_1483
(Fig. 2.2A). The msmeg_1483 open reading frame is the only secY homolog in M. smegmatis
(43% identical and 63% similar at the amino acid level to the canonical E. coli SecY).
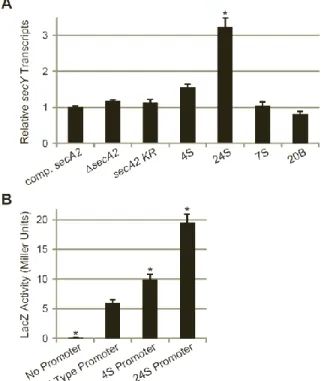
To determine whether any other extragenic suppressors in our collection contained mutations upstream of secY, we sequenced approximately 1000 bp upstream of secY in the 13 additional extragenic suppressor strains. One additional suppressor, identified as 24S (strain NR151), contained a single C→G polymorphism located 134 bp upstream of the translational start site for secY (Fig. 2.2A). The close proximity of the 4S and 24S mutations suggests that they act in a similar manner to suppress the secA2 K129R allele.
4S and 24S suppressor mutations are located within the secY promoter. Because the mutations identified in suppressors 4S and 24S are located upstream of the secY coding sequence, we hypothesized that they would map to the secY promoter and suppress the secA2 K129R phenotypes through an effect on secY transcription. Mycobacterial promoters do not always resemble the classical promoter structure defined for E. coli and other bacteria, and it can be difficult to predict their location. However, manual inspection of the sequence upstream of secY revealed two potential M. smegmatis -10 regions, each beginning with the highly
36
consensus sequence. These two -10 regions, positioned at nucleotides -132 and -140 upstream of the secY start codon, are the closest potential -10 regions to secY, and are located in the same region as the 4S and 24S mutations (Fig. 2.2A). No obvious -35 region was found, but this is not surprising given that mycobacteria have low homology at this region (27), and may not even require the -35 region for promoter function (28).
The location of the potential promoter -10 regions suggests that the 4S and 24S
mutations could directly impact transcription. The 4S mutation is an insertion that improves the -10 region at nucleotide -140 from a 3/6 nucleotide match to the -10 region consensus to a 4/6 nucleotide match. The 24S mutation is a C→G polymorphism upstream of the -10 region at nucleotide -132. Interestingly, this mutation creates a “TGn” motif upstream of the -10 region. Addition of a “TGn” motif to a -10 region can result in a 3-5 fold increase in transcription (29).
To determine the location of the secY transcriptional start site in relation to the 4S and 24S mutations, we performed 5’ RLM-RACE on M. smegmatis RNA (using primer R1, Fig. 2.2B). Following the RACE procedure, we PCR amplified the 5’ ends of cDNA created from secY
transcripts, using a nested PCR strategy. The first round of PCR (using primer R2) amplified out from the secY coding sequence and yielded multiple products. A second round of PCR with a primer upstream of secY (primer R3) narrowed in on the longest secY transcripts. This nested PCR amplification produced two products (Fig. 2.2B) that we cloned and sequenced to