["dian I. Plant Physfol., Vol. XXXV, No.4. pp. 311-320 (Dec., 1992)
SOME INSIGHTS INTO DIFFERENCES IN SEED SIZE IN WHEAT
p.e. PANDE. SHANTRA NAGARAJAN. D. SINGE.* AND R.N. PANDE*
Nuclear Research Laboratory, Indian Agricultural Research Institute, New Delhi-llOOl2
Received on 14 Nov., 1991
SUMMARY
In immature wheat seeds, more accumulation of water in the large seeded type leads to greater enlargement of seed coat, increase in grain weight and volume and decrease in density as compared to small seeded genotypes. More sugar per se as the cause for . higher accumulation of water in immature wheat seed does not seem to be true. The changes associated with the enlargement of seed coat are also reflected in the peduncle as increase in mean diameter and number of vascular bundles per unit area, area covered by vascular bundles per unit area of the peduncle transverse section and increase in peduncle wall thickness. Thus. both the enlargement of grain-sink anj the transport mechanism appear to be closely linked genetic traits and _should be simultaneously con sidered for improvement.
INTRODUCTION
The genotypic variation in grain weight of wheat is the net result of the interaction between variation in potential storage capacity/volume (Jenner, 1974) and variation in realisation of this potential storage capacity which depends on length of grain filling period (Asana and Bagga. 1966), rates of grain filling (Asana and Williams, 1965, Brocklehurst, t977) or a combined interaction of these factors. Moreover, variation in storage capacity of endosperm tissue of wheat grain will depend on the size and number of endosperm cells (Brockiehurst, 1977). Similarly the variation in grain filling is expected to be the result of interaction between availability of assimilates to the grain, metabolism of inter mediates and synthesizing complex (Jenner and Rathjen, 1978). Physically restrcting the development of potential storage capacity of developing grain resulted
312 P.e. PANDE et al.
in significant decrease in final seed size and shape in legumes (Duncan et al., 1978), barley, oat and wheat (Grafiug, 1978), rice (Murata and Matsushima, 1975) and soybean (Egli, Duncan and Crafts Brander, 1987), Thus, the small size of the seed-cost surrounding the endosperm in small seeds might itself act as a physical constraint and restrict the potential storage capacity of the grain, since the peri carp grows first immediately after anthesis, followed by optimal dry matter accu mulation in the endosperm (Jennings and Morton, 1963).
However, very little information is available which gives an insight into morp hological, physiological and biochemical basis for variation in seed-coat development and partitoning of assimilates. Moreover it is not known if the variation in grain weight and associated changes are confined to seed per se or there are associated changes in transport system in peduncle which allows assimilate supply to the ear. The present study is restricted to three wheat genotypes with unusually wide varia tion in grain weight so that the relevant differences associated with genotypic variation in grain weight are better understood.
MATERIALS AND METHODS
Three genotypes of wheat. viz., Kalyan Sona (Triticum aestivum L.), Se1ection III, a hexaploid wheat genotype developed through homologous recOmbination between wheat and rye (Singh and Joshi, 1981) and IWP·5308, a tetraploid wheat genotype developed by transgressing Triticum timopheevi gene (s) in tetraploid wheat genotype (Pandey, H.N., Personal communication) with wide variation in grain weight were used in the present study,
Plants grown in pots (30 cm
x
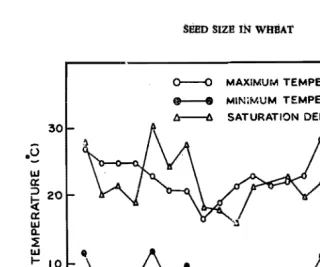
30 cm) having sandy-loam soil rich in farm yard manure. Basal dose of fertilizer consisted of N, P, K (100 : 100 :100 kgJha), The second dose of 100 kg Nlha was split equally and given at tillering and at flowering stages. Four plants per pot were maintained and the pots were kept in a randomised fashion on the benches of a green house and watered equally on alternate days. The weekly weather paramete~ during the experimentation are given in Fig. 1.r
; LLf
. .
.--r.?""-'
SHED SIZI! IN WHBAT
-0---0 MAXIMUM TEMPERATURE
... MINiMUM TEMPERATURE
01
J: tr---6 SATURATION DEFICIT
12.0
e
30e
-.~
IW
Q
0: Lt..
~ 20
8.0 ~
!;(
0: W
Q..
~
~
!;(
w
I 10 4.0 ~
!;(
I/)
o
1...JL...-_ _ _---lL...-_ _ _---I._ _-'--_~:__----'0.047 52 5 10
STANDARD METEOROLOGICAL WEEKS
Fig. 1. Weekly maximum, minimum air temperatures and saturation deficit during the growing period. Arrow mark indicates the approximate time of anthesis.
Peduncle diameter. peduncle wall thickness, number and mean diameter of vascular bundles in peduncle were measured on a thin section cut at mid-length of the peduncle on the 25th day. using a micrometer attachment. Five replicates were used for each genotype. Free sugar content in developing grain was determined according to McCready et al. (1950).
Harvest data and measurements on mature seeds: Ten main shoots were har vested in each genotype at maturity and observations on spikelet number per ear, grain number per ear and grain weight per ear were recorded. Length and width of the seeds were determined by lining up ten seeds end to end and side by side respec tively for the measurements of grain length and width. Volume of the seed was determined through displacement of liquid by putting ten seeds per replicate in a graduated tube containing a definite volume of hexane. Care was taken that no air bubble was trapped among the seeds. Density of the seed was calculated by using the relationship mass/volume=density. Nitrogen content in the seeds was determined according to Anonymous (1971) using an autoanalyser, Protein percentage was obtained by using a conversion factor of 6.25,
!
314
f
CA
S.'''·
i-.c.
PANDBet al.
RESULTS
Except for grain density which shows a reverse trend aU other grain characters show an increasing trend, in the following order: IWP-530S, Selection-III, Kalyan Sona. At maturity, Kalyan Sona showed the highest number of spikelets and grains per ear but does not markedly differ in grain weight per ear (Table I). A comparison of dry matter accumulation in the endosperm and seed coat and thickness of seed coat during grain development among the three wheat genotypes showed that (Table II), seed coat thickness was least in IWp·530S followed by SeI-1II and Kalyan Sona. Seed coat thickness, endosperm and seed weights, however, were always maximum in IWP-530S. Water content per grain in all the three genotypes increased up to the 25th day followed by a decline, whereas per cent free sugar in grain declined sharply after the 10th day (Table Ill). A comparison of morphological characters like pedu ncle waH thickness, mean diameter of peduncle, number of vascular bundles per mm2 , mean diameter of vascular bundles and the mean area covered by vascular bundles in the mid peduncle region of the.three wheat cultivars is presented in Table IV. Kalyan Sona showed lowest peduncle wall thickness, mean area covered by vascular bundle and diameter of vascular bundles, but had maximum diameter of peduncle. The number of vascular bundles was highest in Selection-Ill when com pared to the other genotypes.
Table I. Spikelet number, grain number. grain weight per ear and mean weight, length, volume, density and protein content in basal grains of three wheat genotypes (± SEm)
81. No.
Parameters Kalyan
Sona Selection-III IWP·5308
1. Spikelet number
per ear 23.0±0.48 20.0:1:0.28 17.3±0.38
2. Grain number
per ear 73.5:1: 1.11 36.0±4.51 35.3t 1.64
3. Grain weight (g)
per ear 2.72 ± 0.09 2.47±0.09 2.53:1:0.16
4. Grain weight (mg) 36.9:1:0.2 85.9±0.2 80.9±0.3
5. Grain length (cm) 0.58:1:0.01 0.77:1:0.01 0.98±0.01
6. Grain volume (mn 0.028 LO.OOl 0.05±0.002 O.070±0.OOI
7. Grain Density
(glml) 1.4l±0.O2 1.2l±O.04 1.17±O.OI
SEED sizB IN WHBAT ~lS
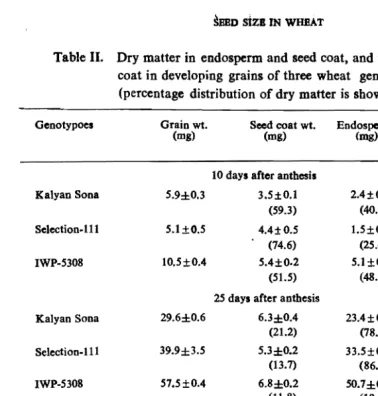
Table II. Dry matter in endosperm and seed coat, and thickness of seed coat in developing grains of three wheat genotypes (± SEm) (percentage distribution of dry matter is shown in parenthesis)
Genotypoes Grain wt. Seed coat wt. Endosperm wt. Seedcoat
(mg) (mg) Crog) tbickness
1'Dl)
10 days after anthesis
Kalyan Sona 5.9±0.3 3.5±0.1 2.4±0.2 248±3
(59.3) (40.7)
Selection.Hl 5.1±O.5 4.4±0.s 1.5±0 2 226±3
(74.6) (15.4)
IWP-5308 10.S±0.4 5.4±0.2 5.1±0.2 193±5
(51.5) (48.6)
25 days after anthesis
I
I Kalyan Sona 29.6±0.6 6.3±0.4 23.4±0.6 71±3
(21.2) (78.8)
Selection-HI 39.9±3.5 5.3±0.2 33.S±0.4 88±1
(13.7) (86.3)
; IWP-5308 57.S±0.4 6.8±0.2 SO.7±0.3 56±2
r (11.8)
(18.2)
35 days after anthesis
Kalyan Sona 39.7±1.4 8.0±0.6 31.8±1.1 51±1
(20.1) (79.9)
Selection-Ill 62.I±O.3 1l.0±0.7 51.1±0.2 45±2
(17.7) (82.3)
IWP·5308 65.9±1.9 12.4±0.7 53.5±1.9 35±1
(18.8) (81.2)
- - - _
..._ " " " "..DISCUSSION
316 P.C!PANDB el
al
Table III. Water and free sugar content in the developing grains of three wheat genotypes
(±
SEm)Genotypes
Kalyan Sona
Selection4 111
IWP-S308
Kalyan Sona Selection4 111
IWP-S308
Kalyan Sona Selection-Ill IWP-S308
Water content (ms/grain)
10 days after anthesis
21.3±3.2
lO.7±1.2
40.1±2.S
25 days after anthesls
29.H2.S 43.3±1.0
7S.7±1.2
35 days after anthesis
3.2±O.3 12.8±O.3
6.1±O.9
Free sugar content (%)
21.4:l:;O.1 34.1±O.1
23.1±O.S
3.S±O.1 3.8±O.2
3.2±O.OS
1.7±O.OS 1.8±O.1
l.S±O.l
Table IV. Morphological characteristics in the mid region of peduncle at 25 days after anthesis in three wheat genotopes (± SEm)
SI. Parameter Kalyan Sona Selection-Ill IWP-S308
No.
1. Peduncle wall thick
ness (/Lm) S08±O.3 622±O.S 793±O.3
2. Mean diameter of
penduncle (/Lm) 27S9±4.0 263S±S.O 2480±4.0
3. Number of vascular
bundles per mm' 79.0±O.2 89.0±O.2 78.0±O.2
4. Mean diameter of vascular bundle
(/Lm) 7S.3±O.3 82.2±O.2 8S.3±O.2
S. Mean area covered by vascular bundle
(mm'/mml) O.3S0±O.OO2 O.469±O.OO3 O.444±O.OO3
SEED SIZlLlN WHEAT 317
Weight, length and volume of the mature grain in the three wheat genotypes were in the fo]]owing decreasing order: IWP-5308, Selection-III and Kalyan Sona but the grain density showed a reverse trand (Table I). This observation implies that unlike Kalyan Sona, the other two genotypes have created a large structural com ponent or storage capacity and have room for further filling and can attain stm higher grain weight, since density of these grains is less than that of KaJyan Sona (Table I). In contrast, in Kalyan Sona large improvement in grain weight is not possible unless the structural component/storage capacity itself is increased. Only small increase in grain weight has been achieved by the treatment of growth regul ator Benzyladenine in Kalyan Sona (Pande and Bhardwaj, 1980). It is important to note that inspite of higher number of spikelets and. grains per ear, the grain weight per ear is not significantly higher in Kalyan Son a when compared to Selec tion-III and IWP-5308 (Table I). This means that maximum synchronisation between structural and synthetic component has been achieved in Kalyan Sona. It is quite likely that small size of seed-coat (structural component) also imposes a physical
I
restriction on final grain weight in Kalyan sona.I
Dry weight per seed-coat was highest in IWP-5308 fo]]owed by Selection-III and Kalyan Sona (Table IJ). This implied that the differential enlargement of seed-coat involves two processes i.e. deposition of more dry matter and more stretching simultaneusly. Since IWP-5308 ill slightly superior to Selection-III in the accumu lation of dry matter per seed-coat, but has nearly two times more water per seed as
r
compared to Selection-III and more than double when compared to Kalyan Sona, it results in maximum enlarged but thinner seed-coat in lWP-5308 followed by Selection-III and Kalyan Sona.
The free sugar content per seed in the three genotypes does not explain the differential maximum water content per seed on the 25th day (Table III). It is, there fore, likely that other osmotically active molecules viz. protein, amino acid and mineral nutrients might also be involved in the observed differential water content per seed.
318
0
P.C. PANDE et al
:z
0.08 (a J ct0:
\!)
~ 006
iJ.J ....
::i E
~.J :; 0·04
~
f
4'"
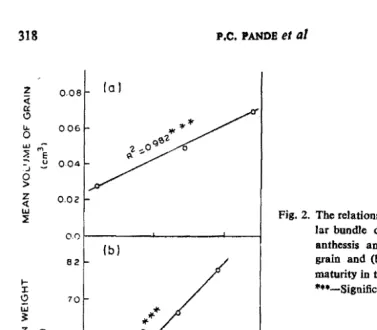
0.0 Jar bundle diameter at 2S days after
82
fb) anthessis and (a) mean volume
grain and (b) mean grain weight of at
l-X
2 10
maturity in three wbeat genotypes. ·"-Significant at 0001 % level;
1.1.1
!t
z
~ i?I 58
a: E
\!)
Z
« 46
1.1.1
.'2
BO
Z 0.02
<t 1.1.1
Fig. 2. The relationship between mean vascu ::E
86
MEAN VASCULAR BUNDLE OIAMETER tAlm I
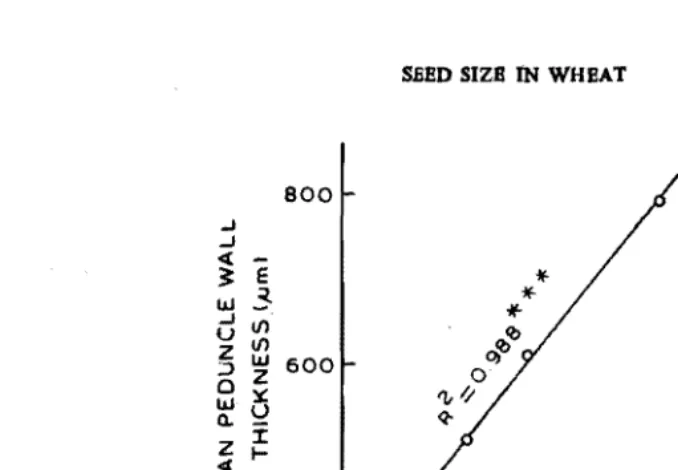
et 01., 1970). High positive correlation was also observed between peduncle-waH thickness and water content per seed on 25th day (Fig. 3).
SEED SIZB IN WHEAT 3J9
800 -'
...J
«
3: IJJE
3
...J U
Z
::>
0
IJJ
Q. til til
IJJ
Z X U
600
Z
«
::t
I
IJJ
~
400
0 40 80
mg WATER / SEED
Fig. 3. The relationship between mg water per seed and mean peduncle wall thickness in three wheat genotypes at 2S days after anthesis.
···-Significant at 0.001 level.
these factors also appear to be genetically linked. It is, therefore, important that wider varie.tion in vascular system should be exploited and incorporated for further improvement of grain weight.
REFERENCES
Anonymous, (1971). Technicon Monograph 1, Technicon Private Ltd .• Tarrytown. New York. Se:;tion 1.1-6.1.
Asana, R.D. and Bagga, A.K. (1966). Studies in physiological analysis of yield. VU. Compari son of development of upper and basal grains of spikelets of two varieties of wbeat. Indian J. Plant Physiol., 9 : 1-21.
Asana, R.D. and Williams, R.f'. (1965). Tbe effect of temperature stress on grain development in wheat. Aust. J. Agric. Res., 16 : 1-13.
Bhattacharya, M.K., Smith, A.M. Noel-Ellis, T.H., Hedley, C. and Martin, C. (1990). Th~
wrinkled-seed character of pea described by Mendel is caused by a trasposon-like in sertion in a gene encoding starch-branching enzyme. Cell, 60: 115-122.
Brocklehurst, P. (1977). Factors controlling grain weight in wheat. Nature (London),266:
348-349 .
Duncan, W G., MC Cloud, D.E. McGraw, R.C. and Boote, K.J. (1978). Physiological as,:ccts of peanut yield improvement. Crop Sci., 18: 1015-1020.
Egli, D.B., Duncan, W.G. and Crafts-Brandner (1987). aff~t ofphysic;ll restfllint QP _d ,rowtll ill sorbean. Crop Sci.~ 27 : ~9-294,
.
.
320 P.C. PANJ)E et al.
Evans, L.T., Dunston, R.L., Rawson, H.M. and Williams, R.F. (1970). The phloem of the wheat stem in relation to requirements for assimilates by the ear. Aust. I. Bioi. Sci., 23 : 743·752.
Orafius, J.B. (1978). Multiple characters and correlated response. Curro Sci" 18 : 931·934. Jenner, C.F. (1974). Factors in the grain regulating the accumulation of starch. Royal Soc.
Newzealand Bull.,ll : 901-908.
Jenner, C.F. and Rathjen, A.J, (1978). Physiological basis of genetic differences in the growth of the grain of six varieties of wheat. Aust. I. Plant Physiol., 5 : 249.262.
Jennings, A.C. and Morton, R.K. (1963). Changes in earbhohydrate, protein, non·protein and nitrogenous compounds of developing wheat grain. Aust. I. Bioi. Sci., 16 : 318·331. McCready, R.M., Ouggals, J. Silviera, V. and Owens, H.S. (1950). Determination of starch and
amylose in vegetables. Analyt. Chem., ll: 1156-1158. .
Murata, Y. and Matsushima, S. (1975). Rice, In L.T. Evans (ed.) Crop Physiology, Cambridge University Press, New York, PP. 73·99.
Pande, P.C. and Bhardwaj, S.N. (1988). Effect of Benzyladenine on endosperm free lipid pool and its fractions in grains of bread wheat (Triticum aestivum). Indian I. Agric. Sci., 58 : . 936-939.
Pande, P.C., Pathak, P.C. and Tomar, O.P.S. (1985). Possible source(s) of oxygen for developing wheat endosperm. Indian I. Plant PhysioJ., 28: 344·349.
Pande, P.C., Sastry, L.V.S., Pathak, P.C., Joshi, M.O •• Raghaviah, P. and Tomar, S.R.S. (1980). Some possible insights into the biochemical basis of grain shrivelling in triticale. Cereal
Res. Cornman., 4: 649-658.
Panwar, J.D.S., Sirohi, O.S. and Pande, P.C. (1986). Evidence for non-involvement of assimilate source in shrivelling of triticale grain. Annals Agric. Res., 7: 355-358.
Singh, D. and Joshi, B.C. (1987). Aneuploid analysis of seed weight in a wheat-rye recombinant.
Indian I. Genu 47: 239-242.