Copyright 0 1985 by the Genetics Society of America
COMPLEMENTATION AND NONCOMPLEMENTATION
AMONG NONALLELIC MUTATIONS ALTERING
DEVELOPMENT IN
SCHZZOPHYLLUM COMMUNE
KEITH K. KLEIN*.t AND CAROL S. DEPPE*
*Department of Genetics and Cell Biology, University of Minnesota, St. Paul, Minnesota 55101, and tDepartment of Biology, Hamline University, St. Paul, Minnesota 55104
Manuscript received January 24, 1984 Revised copy accepted October 5, 1984
ABSTRACT
Mutations resistant to compounds that alter the development of Schizophyllum commune (cyclic adenosine monophosphate, caffeine and imidazole) were selected. These mutations defined four genes, car (CAMP resistance), caf-1 and caf-2
(caffeine resistance) and imr (imidazole resistance). The mutations were found to
be noncomplementing in certain pairs when tested against each other and against two previously isolated developmental mutations, B-con and sep (septateless), in fully compatible, forced heterokaryons. The pattern of complementation in fully compatible diploids showed noncomplementation only between allelic mutations, except for one case, imr us. car.
HE control of dikaryosis in Schirophyllum commune is a complex process
T
involving two genetic elements, the mating-type factors A and B (PAPAZIAN1950). Various parts of the process of dikaryosis have been attributed to the control of one or the other of the mating-type factor (RAPER 1966). We shall describe here a new series of genes, outside the A and B factors, which are active during dikaryosis, and present evidence that suggests a mode of action for these genes.
The strategy we employed to select new mutations in developmentally signifi- cant loci involved the use of compounds known to interfere with developmental processes or suspected of interfering with such processes. These included caf- feine, which alters levels of 3’-5’-cyclic adenosine monophosphate (CAMP) in S.
commune (SCHWALB 1978; DUBOVOY and M U ~ ~ O Z 1979), CAMP added exoge- nously (SCHWALB 1978) and imidazole, a general antifungal compound of un- known molecular action (GODEFROI et al. 1969).
The action of imidazole upon S. commune is complex. At a concentration of 1.8 mg/ml, it causes the disruption of established dikaryons and prevents the formation of new dikaryons by compatible mates. Homokaryons will grow on medium containing this concentration of imidazole, but the morphology of these colonies is altered to a dome-like form with few hyphae penetrating below the surface of the agar. The A = B # heterokaryon and its analog, the B-constitutive mutant (B-con), undergo the most profound changes on this medum. When grown on imidazole medium, these forms lose the morphology characteristic of
334 K. K. KLEIN AND C. S. DEPPE
the A-off, B-on phentoype and gain the morphology of the A-on, B-off phenotype characteristic of A # B = heterokaryons and A-constitutive mutants (A-con). T h e growth rate of these strains on imidazole medium approaches that of homokar- yons grown on medium without imidazole (C. S. DEPPE and M. R. ANDERSON, unpublished results). Caffeine and cAMP have a less severe effect turning off only the B-sequence event of hook cell fusion (DUBOVOY and MuAoz 1979).
We expected to obtain two morphologically distinct types of mutants by selecting for resistance to these compounds. T h e first type would be morpholog- ically similar to its homokaryotic progenitor. This
type
could include those mutants that exclude the antimetabolite from the cell, those that detoxify the antimetabolite and those with an altered structure for some key enzyme normally inhibited by the antimetabolite. T h e second type, and the type that interested us, would be those that displayed a morphology on minimal medium associated with the portion of normal development blocked by the antimetabolite used. For the compounds used these would be “B-on” in character.Analysis of these mutants was carried out by genetic mapping and complemen- tation. T w o distinct sets of complementation tests were performed. T h e first in dikaryons (or fully compatible heterokaryons) and the second with the use of a mutation that causes the formation of diploids rather than dikaryons from compatible mates (KOLTIN and RAPER 1968).
MATERIALS AND METHODS
Minimal medium (MIN) was prepared by the method of ANDERSON and DEPPE (1976). Caffeine
( 5 mg/ml), imidazole (1.8 mg/ml) and cAMP (5 mg/ml) were added to MIN for selecting resistant mutants and scoring resistance in the progeny of a cross. For scoring of auxotrophic mutations used for mapping, arginine, adenine, lysine, uracil, histidine, tryptophan and methionine were added to
M I N at a final concentration of M. Para-aminobenzoic acid, nicotinamide, choline, pyridoxine HCI and pantothenate were added to a final concentration of
All strains used were derived from strains 1736 and 1737 and are coisogenic with strain 699 of the late professor JOHN R. RAPER (ANDERSON and DEPPE 1976). Mutant alleles used are as follows: ade-2, ade-4, ade-11, ade-12 and two other mutations, ade-X and ade-Y of unknown complementation groups, all of which require adenine for growth; arg-6 and arg-8 require arginine; cho-1 and cho-2 require choline; hop requires histidine or pyridine; lys-2 requires lysine; met-l requires methionine; nic-1, nic-2 and nic-3 requires nicotinamide or nicotinic acid; not requires nicotinamide or tryptophan; pab requires para-aminobenzoic acid; panm requires pantothenate; ura-2 requires uracil; caf-1 and caf-2 confer caffeine resistance; car confers cAMP resistance; imr confers imidazole resistance; Dik- renders compatible matings diploid; sep renders strains septateless; A-con renders the Adevelopmental sequence constitutive; and B-con renders the B-developmental sequence constitutive.
Caffeine-resistant mutants were selected by placing plugs of medium containing mycelium from wild-type strain M7 on medium containing 5 mg/ml of caffeine and isolating any mycelium that grew from the inoculum. Imidazole-resistant mutants were sought among colonies from inocula of wild- type strain M4 on medium containing 1.8 mg/ml of imidazole. Only those colonies that displayed a wild-type morphology on this medium were selected, as the wild-type will grow on imidazole medium albeit with a very distinctive, abnormal morphology. The CAMP-resistant mutant was isolated fortuitously as an outgrowth of strain M4 on a plate with an inhibitory concentration of CAMP during a test of the effects of cAMP on a series of strains.
All mutants were crossed to wild-type and auxotrophic strains. Progeny were assessed for resistance and auxotrophy. Any strain that did not yield 1 : 1 segregation for resistance among its progeny was discarded.
The phenotypes of resistant mutants were tested in crosses of A- and B-compatible strains on MIN.
COMPLEMENTATION IN SCHIZOPHYLLUM 335
TABLE 1
Linkage data for caf-1
~~ Progeny type Genotype of progeny Observed no.
Parentals n i c - p , hop-2+, caf-1- 109
n i c - 2 , hop-2-, caf-1' 101
Recombinants nic-2+, hop-2-, caf-1' 19 nic-a-, hop-2+, caf-1- 10 nic-Z, hop-2+, caf-1' 4 nic-2+, hopS-, caf-1- 4 nic-2+, hop-2+, caf-1' 10 nic-2-, h o p - l , caf-1- 22
Inferred gene order:
nic-2-hop-2-caf-1
Crosses were nutritionally forced by use of two complementing auxotrophic mutations, one in each of the strains used for any given cross. Plugs of medium containing mycelia of the two mates were placed side by side to initiate mating. Crosses were fruited where possible (see RESULTS) and monosporous isolates transferred to CYM (SNIDER and RAPER 1965). Subcultures of the progeny were transferred to selective media for scoring. Scoring for growth on MIN and supplemented media was after 72 hr at 30". Scoring for resistance was as follows: after 2 wk at 30" on caffeine medium, after 24 hr at 30" on CAMP medium and after 72 hr at 30" on imidazole medium. Morphology was scored after 72 hr at 30". Mapping was in accord with the linkage groups defined by FRANKEL and
ELLINCBOE (1 977).
Complementation analysis of mutations was performed using fully compatible, nutritionally forced matings on MIN. The analysis was performed twice-once with strains yielding dikaryons and once with Dik- strains yielding diploids. Dikaryosis and diploidy were checked by microscopy and marker recovery from monosporous progeny of mated strains.
RESULTS
A group of
20
mutants was obtained by selection for growth on medium containing 5 mg/ml of caffeine. Only one isolate per selection plate was used in further study. Due to the uncertainty of the number of nucleiper
plug of inoculum, no attempt was made to estimate rates of mutation. Six mutants were found to be due to single-gene mutations, based upon 1:l segregation for the resistance trait (data not included), and were chosen for further study. These mutants occupied two classes, five cuf-1 and one cuf-2 based on complementation analysis and mapping.Cufl mutants are characterized by depressed growth, gnarled hyphae, proto- plasmic extrusions from the hyphal wall and a general resemblance to B-con mutants. T h e five cuf-1 mutants failed t o complement each other when tested pairwise and scored for normal morphology. All five mutants were outcrossed
to isogenic Schizophyllum strains. Mapping by three-factor crosses placed all five of these mutants on linkage group I11 to the right of the last previously known marker on this group, a marker for a histidine or pyridoxine requirement (see Table 1).
336 K . K. KLEIN AND C. S . DEPPE
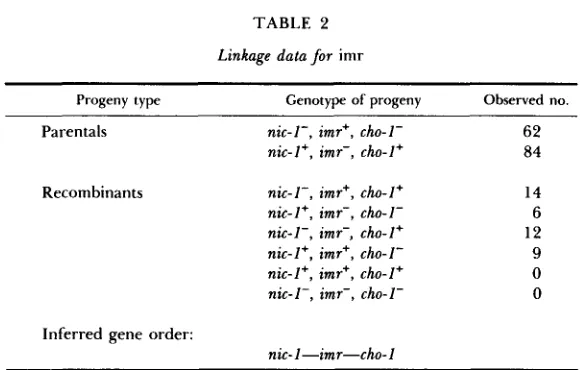
T A B L E 2
Linkage data for imr
Progeny type Genotype of progeny Observed no.
Parentals nic-I-, imr+, cho-1- 6 2
nic-I+, imr-, cho-I+ 84
Recombinants nic- I - , imr+, cho- I + 14
nic-I+, imr-, cho-1- 6
9
0 nic-I-, imr-, cho-I+ 12 nic- I + , imr+, cho- I -
nic-I+, imr’, tho-I+ O nit- I - , imr-, tho- I -
Inferred gene order:
nic-I-imr-tho-1
mentioned. This mutant was labeled caf-2. T h e morphology of this mutant on MIN is of less radical deviation from wild type and consists of a “felty” colonial appearance with shorter aerial hyphae than wild type and twisted, highly branched submerged hyphae. Growth of this mutant on medium containing
2
mg/ml of caffeine is indistinguishable from growth of the wild
type
on caffeine- free medium. In test crosses with the caf-1 mutants, normal complementation and dikaryotization was observed. Mapping crosses failed to assign the gene to any known linkage group or to demonstrate linkage to a variety of unassigned markers. T h e markers tested were: linkage group I, A factor; 11, B factor; 111,caf-1, leu-1, nic-2; IV, ade-12, arg-2; V, ade-4, arg-1, arg-8, lys-2, met, cho, ade-9;
VI, pan-1, arg-6; VII, nic-3; unassigned, nic-1; and several arg- and ade- markers of unknown complementation groups.
Eighty inocula of homokaryotic wild-type strain M4 were placed on separate plates of medium containing 1.8 mg/ml of imidazole. From these inocula one imidazole-resistant colony was isolated. T h e colonial morphology was wild type when grown on imidazole-containing medium. Morphology on medium without imidazole was “flat” with many hyphal aberrations and was reminiscent of B-con
mutants. Upon outcrossing to isogenic
S.
commune strains the characters of abnormal growth on imidazole-free medium and normal growth on imidazole medium were observed to cosegregate in all cases. T h e mutation (imr) segregates 1 : 1 in crosses with wild type. Mapping crosses placed the locus on linkage group IV between loci determining requirements for niacin (nic-1) and choline (cho-1)(Table 2).
A single CAMP-resistant mutant (car) was recovered as an outgrowth of an inoculum of strain M4 during a test of the sensitivity of various
S.
communeCOMPLEMENTATION IN SCHIZOPHYLLUM
TABLE 3
Linkage data for car
Progeny type Genotype of progeny Observed no.
337
Parental arg-6-, car+ 32
arg-6+, car- 67
Recombinan t arg-6-, car- 2
arg-6+, car+ 1
TABLE 4
Complementation data for mutants
Mutation caf- I caf-2 imr car B-con
caf- 1 -1- +I+ +I+ -I+ -I+ +I+
caf-2 -1- -I+ +I+ -I+ +I+
imr -1- NH/- +I+ -I+
car -/NT +I+ +I+
S 9 -1- +I+
B-con NT/NT
+
= complementation;-
= lack o f complementation. Signs before the shilling(4
are results in forced dikaryons (heterokaryons) and signs after the shilling are 'results in forced diploids. Ten trials of each complementation test were performed. N T = not tested. NH = no heterokaryon forms.mapping crosses were possible with this linkage group due to the paucity of
markers and the unavailability of some of the type strains (Table 3).
Mutant strains were tested in all pairwise combinations for the ability to form stable dikaryons. In addition, two other mutations B-con (PARAG 1962) and sep-
(septateless, C. S. DEPPE, unpublished data) were included in these tests. The results, scored for the ability to form dikaryons (+) or the inability to do so (-) in ten trials of each cross, are summarized in Table 4. Cells of complementing crosses were observed by phase microscopy to have two nuclei
per
cell and clamp connections; they displayed none of the nuclear aggregations or gnarled hyphae typical of the mutant homokaryons or of noncomplementing crosses. Mutants mated with fully compatible wild-type strains produced dikaryons which were indistinguishable from wild-type dikaryons, either in morphology or with respect to resistance; the mutations are thus recessive.338 K. K. KLEIN AND C. S. DEPPE
All mutants displayed a unilateral mating (i.e., acting as nuclear donors only and not acceptors) in crosses not subject to nutritional forcing. This inability to accept nuclei always segregated with the resistance phenotype or the morpholog- ical phenotype.
DISCUSSION
We present data describing several new mutations of
S.
commune which affect development. These mutations confer abnormal phenotypes upon homokaryons that carry them. Furthermore, matings between otherwise compatible mutants yield abnormal development during dikaryosis. The pleiotropic nature of the cafand imr mutations is revealed by the morphology that they confer upon strains carrying them. This morphology is altered from the wild-type homokaryon and more closely resembles the B-constitutive mutant. The mutations define five previously undescribed loci: car, caf-1 and caf-2; imr; and sep. None of these loci is closely linked to any other or to the A and B mating-type loci. When otherwise compatible pairs of mutants are nutritionally forced on MIN, the normal process of dikaryosis fails in six of a possible 15 cases. The resultant heterokaryon in these cases does not form a stable dikaryon but, instead, persists in a state that does not have cells with fixed numbers of nuclei, has many partial septa, cell wall abnormalities and other characteristics of the parental mutant strains. The use of Dik- to produce diploids leads to normal development in crosses between 14
of the 15 pairs. One pairing, that of imr and car strains, never produces a mycelium capable of growing on MIN heterokaryotic or diploid. Since such strains were shown to be compatible for mating type and they contained auxo- trophic markers that complemented in strains not carrying imr or car, it may be that there is no hyphal fusion between strains carrying imr and strains carrying
car.
Two possible interpretations present themselves for the other five of 15 pairs of mutants that did not form dikaryons. First, it may be that the two mutant strains do not form stable dikaryons as a consequence of the overlapping effects of two pleiotropic mutations together in one heterokaryon. This would not be
simply due to some interaction between mutations but, rather, to mutational disruption of physical events necessary to the maintenance of dikaryosis which would result in a departure from a balanced 1 : 1 nuclear ratio and, therefore, to an assertion of the phenotype associated with one or the other of the homokar- yons (DUBOVOY 1976). The diploid always maintains a 1 : 1 genomic ratio and thus would be stable.
COMPLEMENTATION IN SCHIZOPHYLLUM 339
homokaryons by sectoring (data not shown). Examination of the sections shows each sector to contain one or the other of the nuclear types present in the heterokaryon and no recombinants. We cannot rule out recombination, however, since very few markers were present in each cross.
Clear precedents exist of mutations that complement in diploids but not heterokaryons. APIRION (1966) showed this pattern for thefac genes of Aspergil-
lus nidulans. ROBERTS (1 964) showed similar results for sb mutants of A. nidulans. Of these two examples, the first involvingfac loci is more like the current work.
In both that case and the current work, the genes involved are unlinked by
recombination, whereas in the case of sb very close linkage has been shown. Although we prefer a mechanism based upon nucleus-limited complementa- tion, we have not ruled out a more mechanical model. Experiments are now underway to try to distinguish between these two possibilities.
LITERATURE CITED
ANDERSON, M. R. and C. S. DEPPE, 1976 Control of fungal development. 1. T h e effects of two
Recessive mutants at unlinked loci which complement in diploids but not in
A class of genes affecting B factor-regulated development in Schizophyllum
Effect of methylxanthines in sexuality of Schizophyllum commune.
New mutations and a 7-chromosome linkage map of
GODEFROI, E. F., J. HEEVES, J. VAN CUTSUM and P. A. A. J. JANSSEN, 1969 T h e preparation and
KOLTIN, Y. and J. R. RAPER, 1968 Dikaryosis: genetic determination in Schizophyllum. Science
PAPAZIAN, H. P., 1950 Physiology of the incompatibility factors in Schizofihyllum commune. Bot. Gaz.
Mutations in the B incompatibility factor of Schizophyllum commune. Proc. Natl. regulatory genes on growth in Schizophyllum commune. Dev. Biol. 53: 21-29.
heterokaryons of Aspergillus nidulans. Genetics 53: 935-94 1.
commune. Genetics 82: 423-428. APIRION, D., 1966
DUBOVOY, C., 1976
DUBOVOY, C. and A. MuSoz, 1979 Mycologia 71: 855-861.
FRANKEL, C. and A. H. ELLINGBOE, 1977 Schizophyllum commune. Genetics 85: 4 17-425.
antimycotic properties of derivatives of 1-phenethylimidazole. J. Med. Chem. 12: 784-79 1.
1 6 0 85-86.
1 1 2 143-164. PARAG, Y., 1962
Acad. Sci. USA 48: 743-750. RAPER, J. R., 1966
ROBERTS, C. R., 1964 Complementation in balanced heterokaryons and heterozygous diploids of
SCHWALB, M. N., 1978 Regulation of fruiting. pp. 135-165. In: Genetics and Morphogenesis in the
SNIDER, P. J. and J. R. RAPER, 1965 Nuclear ratios and complementation in common-A heterokar- Genetics of Sexuality in Higher Fungi. Ronald Press, New York
Aspergillus nidulans. Genet. Res. (Camb.) 5 21 1-229.
Basidiomycetes, Edited by M. N. SCHWALB and P. G. MILES. Academic Press, New York.
yons ofSchizophyllum commune. Am. J. Bot. 5 6 547-552.