MUTATION AND RECOMBINATION
IN
THE VICINITY OF A COMPLEX GENE’L. C. DUNN, DOROTHEA BENNETT, AND A. B. BEASLEY
Columbia University, N e w York, N e w York
Received September 1, 1961
HE processes by which a genetic locus assumes diverse forms of expression are of interest both as sources of the genetic variability on which evolutionary forces may act and as indicators of the potentialities of the locus in controlling metabolic and developmental processes. The first type of diversification to be detected was that which results in the origin of a new allele, with properties different from the allele from which it arose. Allele mutation of this kind is probably the elementary step in the diversification of the genetic material. In practice, however, it is often difficult to distinguish such an event from the intragenic recombination which has been shown to occur within many series of multiple alleles. ( PONTECORVO 1958). “Absence of recombination” amongst the different forms expressed by a locus cannot be used as an absolute criterion of allelism, first because its value is only that of a probability statement limited by a known number of negative observations and second because recombination may be prevented by structural peculiarities at the locus or in its vicinity. A better criterion is that alleles should show noncomplementary interaction with each other. This is based on the assumption that allelic changes occur at the same site of mutation within a larger multisite unit of function. Except in the case of lethal alleles, this criterion is also difficult to apply in practice.
Because these two criteria for allelism are based on different kinds of data, which may not be obtained with equal ease in any particular situation, it is to be expected that some loci will be found at which variants fulfill one criterion but not the other. The finding of either recombination or complementarity alone must therefore sometimes suffice as evidence of diversification within a functional unit region; where evidence of diversification has been found, the functional unit has been referred to as a complex locus or complex gene (DUNN 1956). PONTECORVO (1958) has pointed out that most genes within which such evidence of diversification has been sought by methods competent to disclose it have proved to be complex in the above sense.
In the cases in which complexity is shown only by complementarity, and not by recombination, one may suspect that recombination is prevented by some form of chromosomal aberration. The first example of this sort was provided by mutations at o r near one locus in the house mouse. The first mutant identified
1 Research carried out at Nevis Biological Station, Irvington-on-Hudson, New York, under contract A T (30-1) 1804 with the U. S. Atomic Energy Commission, with the assistance of Frank Burnett, Bill Bdand, JoAnne Munigle, and Charlotte Stoeber.
286 L. c. DUNN, et al.
had a short-tailed phenotype (known as Brachyury or Brachy) in heterozygotes T / + and was lethal when homozygous T / T (CHESLEY 1935). Subsequently a number of forms of this locus, known collectively as t alleles, were identified in laboratory stocks and in different wild populations of Mus, most of which proved to be polymorphic for variant alleles at this locus.
All t alleles, in spite of diversity in other effects, share the common property (by which they are detected) of producing a tailless phenotype when combined with T , the first lethal mutant to be identified at this locus. Some t alleles are viable when homozygous but differences amongst some of these have been de- tected by the study of other properties, e.g. male transmission ratios (Table 3 ) , relative fertility of homozygous males and sterility of males heterozygous for
certain viable and certain lethal t alleles.
The lethal t alleles also include a variety of forms differing in the time and manner of homozygous expression, in effects on certain early organization processes in the embryo, in the ratios in which they are transmitted by male heterozygotes and in the degree to which they interfere with recombination in the vicinity of T locus. The decisive evidence of difference or nonidentity of any two lethals is that they complement each other producing a viable phenotype when combined in compounds.
These indications of diversity in some respects and of similarity in others, coupled with lack of evidence of recombination within the locus, led to extensive breeding analyses of variants at this locus, which had as their object the assess- ment of the relative roles of allele mutation and of recombination in the vicinity
of the locus in producing the great variety at this complex locus. In this paper, breeding results obtained up to the end of 1961 are summarized and analyzed in relation to the above problems.
M A T E R I A L S A N D METHODS
The method of maintaining variants at this locus in balanced lines has been described in previous papers (DUNN 1956, 1957; DUNN and SUCKLING 1956). The essential results from studies of the first alleles were: ( I ) that whereas all homozygotes T / T , to/to, and tl/tl acted as lethals in early embryonic stages, compounds of these ( T/to, T/P, and t o / t l ) were viable, the first two being tailless, the last having a normal tail; (2) recombination between the three forms was not observed to occur in some 6775 observations (DUNN and GLUECKSOHN- SCHOENHEIMER 1943). Subsequently other lethal alleles were found which interacted with T to produce taillessness, but could be distinguished from each other by complementarity without evidence of recombination (DUNN 1956). These conditions made it possible to maintain individual lethal alleles in balanced lethal lines ( T / t x T / t ) without selection (CHESLEY and DUNN 1936).
Later other forms of this gene were discovered which in compound with the original mutation T also produced a tailless phenotype ( T / t )
,
but when homozy- gous produced a viable, normal-tailed phenotype ( t / t ).
R E C O M B I N A T l O N I N M I C E 28 7
a recurring process. Exceptions which were normal-tailed were found in the long-maintained balanced lethal tailless lines ( T / t " ) . When analyzed these exceptions proved to be compounds tn/t", tn being the parental form, t", a new and different form. From these, new balanced lines T/t" were derived, the t" forms receiving numbers t3, t4, etc., in the order in which they were detected. When crossed with the parental form, T/t" tailless animals regularly produced normal- tailed tn/tx offspring in addition to the expected tailless ones T / t n and T/t". The complementarity of the old and new forms of the gene was expressed in two ways, tn/tx having a normal phenotype in respect of effects on the tail, and being viable even when t n and t" were both lethals. This complementarity provides the
essential criterion by which forms of the gene are distinguished as different. Secondly, new forms of the gene were found in heterozygotes +/tw as regular components of various wild populations. These have given rise to new balanced lines (T/t"', T / P , etc.) maintained in the laboratory. These in turn have pro- duced exceptions from which Eurther balanced T/t" tailless lines have been derived. In three cases, a single +/tw heterozygote from a wild population gave rise to two different balanced lethal lines, one exceptional gamete having trans- mitted a form of t different from that for which the parent was heterozygous. Thirdly, following the discovery by LYON (1956) of a new recessive hair loss mutant (tufted, t f ) located near to T locus, this marker was introduced into all balanced lines in this laboratory. (We are grateful to DR. LYON for stock of this mutant.) Most t alleles have since been carried by matings of T tf/tn
+
XT t f / t n
+.
When normal-tailed exceptions tn/tx occur in such balanced lethal lines, they can provide the additional information whether the chromosome bearing tx has also acquired the marker by recombination. If in such lines a new phenotype tailless tufted ( T t f / t n t f ) is found it can be tested to determine whether the t allele in the recombinant chromosome is the parental one or a different one, t". LYON and PHILLIPS (1959) have already shown, as discussed later, that in several cases the occurrence of normal-tailed exceptions in a balanced lethal line is associated with exceptional recombination in the T-tf interval.The breeding behavior of 49 balanced lines has now been observed. I n 27 lines, the t allele is known to be lethal; in 22 lines it is viable, homozygotes having normal tails.
R E S U L T S
Lnthal alleles: The results of tailless by tailless matings within each of the balanced lethal lines are shown in Tables 1 and 2. Omitted are results from lines discarded: t Z which appeared to be a duplicate of t'; tw4, a duplicate of t o ; tW9,
about which doubt arose as to its origin.
288 L. c. D U N N , et al.
TABLE 1
Results of matings within balanced lethal tailless lines classified at birth. All matings were of the type tailless T / t 0 >( tailless T/t 8 . The chromosome containing T carried
no known marker gene
Offspring
Nommal Normal No. of Exceptions
Line Tailless tail alive tail dead' litter..; (nt indicates normal tail)
t o 34321
t' 3462
t 1 (newdata) 204
24 334
t 9 225
t'2 982
t20 30
tu' 599
t Z V 3 492
tw5 1239
t l U 6 81 7
tQ11 738
tw12 733
tW1S 328
t W ' 4 238
t W 1 5 108
t W 1 6 22 1
t W 1 7 285
t i 0 1 8 1110
t W 2 0 344
t t V 2 1 117
5 11
1
3
15
1 5
4
2 1
2
1 1
930 942
53
240
193 144 332
254 21 1 191 90 67 30 60 95 285 94 32
1 n t lost; 4 nt allele not recovered
cf. D U N N and GLUECKSOHN-WAELSCH, 1953.
( t alleles isolated: t S , t 4 , ~ 7 , t R , t 9 , t " , t ' 2 ) 1 0 n t t l / t 2 2
cf. D U N N and GLUECKSOHN-WAELSCH, 1953
Totals 16,038 31 24
Dead at birth with microcephaly and diagnosed as surviring homozygotes t"'/t"', etc. (BENNETT, D U N N and
-j Includes 144.9 tailless reported i n Drrivx and C T ~ n r . c ~ ~ o a r \ - S c r r o ~ ~ ~ ~ r ~ i ~ ~ (1943)
BADENIIAUSEN 1959).
other symptoms of the lethal syndrome associated with homozygosity for the lethal t allele in lines ttul, tqu3, t z v f 2 , tfc20 and tfUz1 (BENNETT, DUNN and BADEN-
HAUSEN 1959). These represent homozygotes which survived until just before
birth. Of the remaining 46 exceptions progeny were obtained from 28 when test- crossed by Brachys ( T / f ) . The objective of this cross was to discriminate be- tween two hypotheses of the origin of these exceptions. One is that they represent recombinants
+
+/+
t produced by crossing-over between T and t. In this case they should produce Brachy offspring when tested by T/+. The other is that the normal-tailed exceptions result from changes from the parental form of t% to a new form t", complementary to it. The genotype of the exceptions should on this hypothesis be t"/t" and when tested by T / + should produce only tailless ( T / t n , T/t") and normal (+/t", + / t n ) , never Brachy.RECOMBINATION IN M I C E 289
TABLE 2*
Results of matings within balanced lethal tailless lines classified at birth. All matings were of the type T tf/t+ 9
x
T tf/t+ 8, tf being a recessiue marker about ten crossouer units from TOffspring
NFrmal Normal No. of Exceptions
Line Tailless tail alive tail dead+ litters (nt indicates nonnal tail) ____
t o 1823 2 530 1 8 nt to/t23; 1 9 nt t 0 / t a 7 t f
t' 949 3 226 1 8 nt t l / t 2 8 tf;. 1 Q nt (being tested) ; 1 nt lost
t'2 6 1 4 1 . 164 1 9 n t t 1 2 / t 8 5 if
t 2 o 436 1 124 1 8 nt sterile
t W' 726 1 1 236 1 8 nt (being tested)
t W 8 545 1 140 . . .
tw5 368 99 . . .
2 6 4 7 9 . . .
1 9 0 51 . . .
tw1o
223 65 . . .
tW"
tw1e 760 3 202 1 9 nt tWI*/tt~3o t f ; 1 9 nt died;
t W 1 S
t W 1 5
tW16 3 7 9 2 99 2 8 nt sterile
tw17
tWI8 576 1 132 1 0 nt tt018/tW29 tf
t W 2 0 tw21
t W 3 2 185 1 5 2 1 0 nt (being tested)
Totals 9,713 15 6 2,663
t W 6
1 9 nt-allele lost 267 71 . . .
250 75 . . .
152 46 . . .
507 1 131 . . .
4 9 9 3 141 . . . .
* The,ongins of balanced lethal lines in this table have been reported in previous publications except T tf/t"'lO,+ which was demved from wild heterozygotes kindly collected for us by Miss VIRGINIA VINCENT from Clark Fork River near Clinton, Montana.
t Dead at birth with microcephaly and diagnosed as surviving homozygotes t w D ' / t W ' , etc. (BENNETT, DWNN and BADENHAUSEN 1959).
detected with a normal allele (+) at this locus. All exceptions tested proved to be t"/t". New tailless lines were extracted from the T/t" progeny from the testcross. It is of interest that of the exceptions which gave progeny 21 were females yield- ing an average of 31 offspring each while only five were males yielding an av- erage of only nine offspring each after long-continued testing with four fertile females. These males would be classed as quasi-sterile (DUNN 1952), a condition associated with heterozygosity for different t alleles, at least one of which is a lethal (t"/t"). Nine male exceptions yielded no progeny at all after long- continued testing. In only three cases was a new balanced line derived from a male exception.
290 L. c. D U N N , et al.
total progeny (25,751/2 = 12,876) can be used in computing fiducial limits. The 95 percent limits for 0/12,876 are 0.0-.0001.
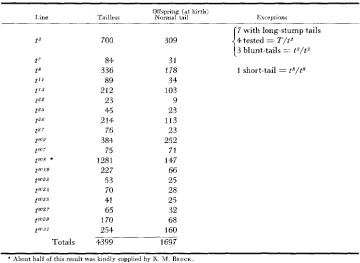
The results of matings of tailless animals in lines containing viable t alleles are given in Table 5. These produced regularly only two phenotypes-tailless and normal-with occasional variants which proved not to be genotypically different from these. Changes from t" to P cannot of course be detected in such matings since T/t" is expected to be tailless and t E / t n normal-tailed, like the regular offspring. But if recombination between T and t were occurring, the recombinant class f
+
(x
of all) should be detectable when combined with one class of gametes, T , constituting about 1/2 of the regular gametes. The efficiency of this method of detection is thus limited to about of the gametes. The total number of offspring observed was 5.096 (Table 5 ) and no exceptions represent- ing recombination of T and t were found. The 95 percent fiducial limits of 5096/4 = 0/1224 are 0.0-.0015. Without excluding such low probability of recombination between T and t , we can say that the observations to date include no exceptions which can be ascribed to recombination between T and any t alleles. Thus with the reservations noted, we may treat T and t as genetical alleles.Probabilities of detecting new uariant alleles: The probability of detection of a variant t" allele in progenies from a balanced lethal line T/t" x T/t" depends upon the detection of the exceptional phenotype tn/t" (with normal tail) at birth. This will depend upon (1 ) the probability ( U ) of occurrence of a variant gamete;
(2) the probability ( m ) of the occurrence, in the mate, of a gamete competent to produce. in combination with the variant gamete, the exceptional phenotype t"/t"; ( 3 ) the probability (s) of survival of this phenotype until birth. The total probability of detection will thus be ums assuming the events concerned to be independent and other parameters to be normal.
In the case of most of the balanced lethal lines studied, m (the gametic ratio)
is always 0.5 for females T / t ; for males it is usually between 0.85 and 0.99, say 0.9. Hence for variants occurring in sperm from T/t" males (to be detected through eggs from T/t" females) the probability of detection is 0.5~s; for variants in eggs it is 0.9us. Hence, from males about half of the variants producing excep- tions with normal viability might be detected at birth and about 9/10 of similar variants in females. But it is known from direct observations that some variant phenotypes have very low viability before birth. For compounds t l / P for example s = 0.15 (DUNN 1956). The probability of detection of a variant such as
t Z 2 from t' would thus be 0.5 x 0 . 1 5 ~ from males, 0.9 x 0 . 1 5 ~ from females. Thus
only about seven percent to 13 percent of such variants would be detected at occurrence.
RECOMBINATION I N MICE 29 1
then the probability of detection would be less for new lethals than for new viables and we should by this method underestimate the occurrence of lethals. The observed numbers of variant alleles derived from exceptions and tested for viability is at present 19 viables and nine lethals. The difference may be due to different probabilities of detection rather than to a higher rate of occurrence of viable variants.
Apart from probability of detection, the probability of establishment of lines carrying new t variants will depend on the fertility of compounds tn/t" at origin. Most male compounds of this type are either quasi-sterile or completely sterile and even with continued mating in the laboratory these rarely give rise to new balanced lines.
In summary then estimates of frequency of occurrence of new forms of t when detected through exceptions tn/tz will in general be underestimates. If full proof that a variant is new and different requires its isolation in a balanced line for comparison with all others, this will be obtained for a small fraction of the exceptions that occur, at least for the conditions of these experiments.
A reference summary of all t alleles observed in this laboratory through SeDtember 1961 is given in Table 3.
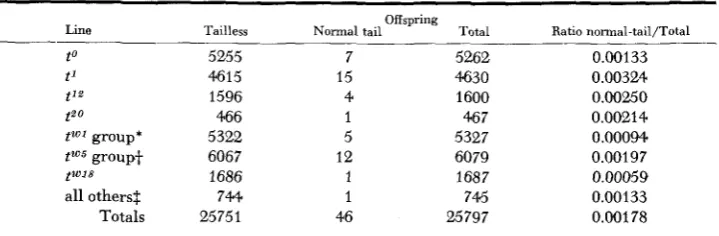
Frequencies of exceptions in lines balanced with different t alleles: W e may now examine the frequencies of exceptions in tailless lines balanced for different lethal t alleles. The data are shown in Table 4. Here comparisons are made among different lines known to contain t alleles which are nonidentical, that is, those which in crosses between lines produce viable normal-tailed compounds tn/'tx. The average frequency of exceptions in all lines is just under two per thousand offspring at birth, but the frequencies vary considerably. The
x2
for homogeneity among the frequencies shown in Table 4 is 23.78, df 7, P = 0.01. This is chiefly due to the contribution toxz
of 17.95 from the ti line which has produced the highest frequency of exceptions. However, it has recently been found (SILAGI, in press) that the allele in line t' is now genetically and embryo- logically indistinguishable from t o and it is probable that the change in this line occurred about 1953. The data for t 1 in Table 4, therefore, probably include some animals carrying a to-like allele. If comparison is restricted to the period before this change occurred (second line of Table 1) it appears that ti then showed an exception frequency about twice as high as t o . The difference between the fre- quencies in the two lines at that time however is not significant at the five percent level. Thus it cannot be concluded that the production of a higher frequency of exceptions is an inherent property of the t 1 allele.292 L. c. DUNN, et al.
TABLE 3
Source and chief effects of uariant alleles at locus T
Ma!e
transmission
Allele ratiot Homozreote Derived from
.50 .73 .87 .45 .4u .50 .50 .45 .50 .90 3 5 .84 .50 .50 .50 .57 .41 .89 .95 lethal lethal lethal lethalt
(v) normal-tail
I
lethal ? lost lethal
(v) normal-tail+ (v) normal-tail lethal
? lost
(v) normal-tail+ lethal
(v) normal-tail
lethal ? lost
(v) normal-tail+ ? lost
(v) normal-tail+
(v) normal-tail+ (v) normal-tail
( v ) normal-tail (v) normal-tail lethal
( v ) normal-tail; 8 sterile lethal
lethalt lethal lethal
(v) normal-tail; 8 sterile+
(v) normal-tail; 8 sterile lethalt lethal lethal lethal lethalt lethalt lethalt lethalt lethalt lethal
DOBROVOLSKAIA-ZAWADSKAIA, 1927 From outcross of T/+ by normal-tailed
stock-DOBROVOLSKAIA-ZAWADSKAIA
and KOBOZIEFF, 1932
and GLUECKSOHN-WAELSCH, 1953 exception t l / t 3 from T / t l X T/t'-DUNN
exception t'/t4 from T/t1 X T/t'
exception t'/t5 from T / t l X T / t l
found in normal-tailed stock by CARTER
exception tl/t7
1
exception t'/t8
I
From T/t' X T / t lexception t'/t9 } DUNN and
GLUECKSOHN-
J
WAELSCH, 1953 exception t'/t'oexception t'/tIl
exception tl/t'S from T / t l X T/tl--
DUNN, 1956
exception t12/t'3 from T/t12 X T/t12 and PHILLIPS, 1950
exception t'2/tzo from T / t l 2 X T/t12
exception t'/tZz from T/tl X T/t'
exception tO/W from T/tO X T / t o exception t'z/t24 from T/t'* X T/t12
exception t'2/t25 from T / t I 2 X T / t l *
exception T t f / t 2 6 t j from T tf/t'
exception t o / t 9 7 from T/tO X T / t o
exception t J / P from T / t l X T/t' X T t f / t l
Each from single f / t found in a wild population (DUNN and SUCKLING, 1956)
Each from single +/t
found in a wild
population (DUNN, 1957)
exception T/t1u'8 from +f/t7"" X T / +
RECOMBINATION I N MICE
TABLE 3-Continued
Source and chief effects of variant alleles at locus T
293
-
Male transmission
ratio$ Honiozygote
- Allele .38 .99 .99 .5 .5 .5 .5 .48 .41 .53 .83
(v) normal-tail; 8 fertile lethal
lethal
(v) normal-tail; 8 fertile+ lost
(v) normal-tail; 8 fertile+ (v) normal-tail; 8 fertile+ ( v ) normal-tail; 8 fertile+ ( v ) normal-tail; 8 fertile+ ? lost
(v) normal-tail; 8 fertile lethal
(v) normal-tail; 8 fertile lethal
Derived from
exception T/rW20 from + / P I 5 X T / +
exception T/tw*l from + / P I 7 X T / +
exception tw5 / t w z 2 from T p 5 X T/tW5
exception 1203 /tw24 from T / F X T / t w s
exception t w 1 7 / ~ 5 from T/tw17 x T / F 7
exception tto* / P Efrom T / P 6 x T/tW6
exception T/tW27 from + / F o X T / +
exception tW12/twzB from T / P l Z X T / t W 1 z
exception tw18/t*z9 from T/"8
x
T / t W l 8exception tW'P/tW3'J from T/tw12 X T/twlz
exception tw5 /tw31 from T / t w 5 X T/tW5
exception t W 1 O / t W S * from T / P l O x T/tw10
* t*4-t*9 numbers reserved for exceptions to be analysed by other investigators.
+
Line discarded.t For detailed data cf. DUNN 1953, 1960; ratios for P, tS7, ~ ' 9t'3o , based on new tests of five T / t males; t W J ' on three
and t W 3 * on four T / t males.
(v) Homozygote viable a t birth.
TABLE 4
Frequencies of normal-tailed exceptions found alive at birth in balanced lethal tailless lines, obtained b y combining results of Tables 1 and 2
Line
t o t' t'2 t2o
tWJ group'
tW5 group+
twJ8 all others$ Totals Tailless 5255 461 5 1596 466 5322 6067 1686 744 25751 Offspring
Normal tail Total
7 52.62
15 4630
4 1600
1 46 7
5 5327
12 6079
1 1687
1 745
46 25797
Ratio normal-tail/Total - 0.00133 0.00324 0.00250 0.00214 0.00094. 0.00197 0.00059 0.00133 0.00178
* t"', t w 3 , t"l2, t"l", t ~ ~ l - al le le s not distinguishable genetically or embryologically (BENNETT, DUNN and BADEN-
t t"', t"', t""', t"", i'0'3, t a x * , tW1', t"16, Px7-alleles not distinguishable (DUNN and BENNETT 1960).
HAUSEN 1959).
$ t4, t 9 , 1 - 3 1 ,
Effect of different t alleles on regular recombination: Difference amongst t alleles in their effects on recombination was first shown (DUNN and CASPARI
294 L. c. D U N N , et al.
T and tufted ( t f ) a distance of about eight crossover units, while t6, a lethal indistinguishable from to, suppresses recombination in this region.
In order to classify other t alleles according to their effect on recombination in this region, the marker tf, obtained from DR. LYON, was used. Before inserting it in the balanced lines, however, the location of tf was determined with reference to
Ki,
the marker previously used. The location data are given in Table 6. The locus of tf is obviously close toKi
hence on the same side ofT
asKi.
The average recombination of tf with
Ki
is somewhat less than one percent,TABLE 5
Results classified at birth of matings within balanced tailless lines in which the
t allele was viable when homozygous
Offspring (at birth)
I l n e Tailless Normal tail Exceptions
Totals
700
84 336 89 212 23 45 214 76 384 75 1281
227 53 70 41 65 170 254 4399
7 with long-stump tails 4 tested = T/t3
309
I
(3 blunt-tails = t3/t”
31
178 1 short-tail = tR/t8
34 103 9 23 113 23 252 71 147 66 25 28 25 32 68 160 1697
* About half of this result was kindly supplied by K. M. BRUCK
TABLE 6
Results of test crosses t o measure recombination between the loci of Kink
(Ki) and tufted ( t f )
Parents Offypring
Female LMde Ki
+
+
tf++
Ki tf TotalsK i
+/+
t f+
t f 21 7 296 5 1 519+
t f Ki+/+
t f 279 270 1 1 55 1Totals 496 566 6 2 1070
Recombination fraction: p 61519 = o . n i i 5 c n . n o 4 ~
o” 21551 =n.on3i*n.ooz6
RECOMBINATION I N MICE 295
being somewhat more frequent in female heterozygotes as is usual in the mouse. The accurate determination of the Ki-tf interval permits a better estimate of the T-Ki interval as 8-1 0 crossover units.
W e have also tested recombination between tf and a mutant ( T " ) with effects like T found by DR. NAOMI FITCH and supplied to us for testing. Testcrosses by tufted gave the following results:
from male heterozygotes-seven recombinants/l58 off spring = 4.4% from female heterozygotes-12 recombinants/ll5 off spring = 10.4%. The recombination data are compatible with the assumption that DR. FITCH'S T is unilocal with the original T . Whether it is a recurrence of T by independent mutation or an example of T hitherto suppressed in the stock in which it occurred cannot be decided from recombination data.
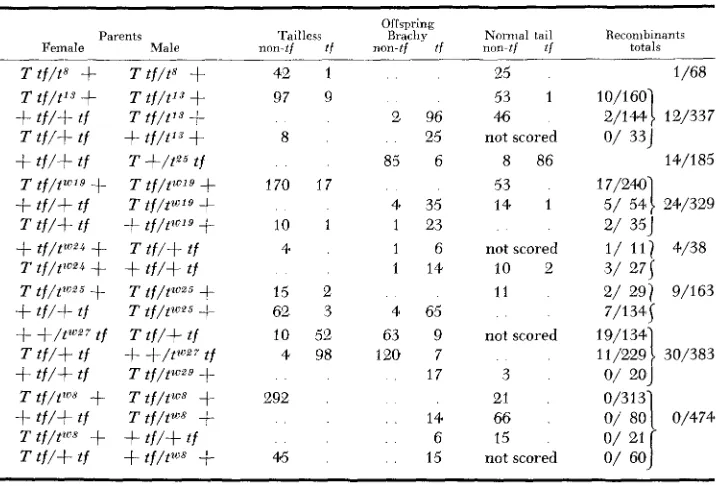
Tests of lethal t alleles as suppressors of regular recombination: The results of testing lethal t alleles for recombination suppression are shown in Table 7. Most
of the test data result from matings T tf/tn
+
X T tf/tn+.
Recombination be- tween the loci of T and tf should produce gametes T+
and tn tf. In this mating, union of these with nonrecombinant gametes T tf and tn+
should produce (1) T +/T tf (lethal) ; ( 2 ) T +/tn+
(tailless nontufted, not distinguishableTABLE 7
Tests for effects of lethal t alleles on recombination in the T-tf interval, offspring scored for tufted after 28 days
Par en t s
Female Male non-tf Tailless tf
Offspring Brachy
non-tf t j non-tf Noma1 tail t j Recombinants totals
481 30
259 I * 15 133 178
252 151 27 93 110 118
15 I t 104
57
13 2 116 10
26 3 24 1
40
8
not scored
not scored
18
15
2 38 12 17
not scored
. .
4 4 2 not scored
0/48 1
O/ 70
1 /NO
O / 23 0/133 0/178
O / 18
0/252
0/151
O/ 27
o/
93 0/110 0/1181/ 31 0/104
a/
572/ 15 10/126 4/ 86 3/ 41 1/ 42
* = T t f / P t f .
296 L. c. DUNN, et al.
from the nonrecombinant class T t f / t n +); ( 3 ) T tf/tn tf (tailless tufted); (4) tn t f / t n
+
(lethal). Only class 3 can be detected in F, so the test is not efficient.From such matings in which the t allele was to, t', t 1 2 , t", (tZV', ttviZ, tLoZo, t W z 2 ) ,
(tW6, tW", tWIG) or t7"32 (alleles belonging to the same group of indistinguishable alleles are enclosed in brackets) 1964 tailless offspring were observed after 28 days and all except one was nontufted. The exception, a tailless tufted from T tf/t'
+,
w a s shown to contain an allele which was not t', i.e. t S F , a viable. From other testcrosses of four of the above lethals 138 offspring were observed, all nonrecombinants except one tailless tufted from+
tf/tlL'Zo+
but this exception proved to contaiq an allele other than tweo, a viable tWZ7. Thus of 2102 offspring observed, none represented regular recombinants and we may tentatively classify the lethals tested as recombination suppressors.The results were different with lethal alleles t9 and tW18 which are probably indistinguishable although of independent origin. From testcrosses of these, 20 regular recombinants out of 310 were observed, so these lethals do not suppress recombination.
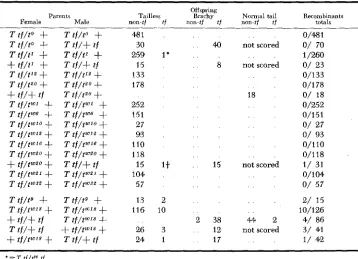
Tests of eflects of viable alleles on recombination: Table 8 shows the results of matings of animals heterozygous for a viable t allele and for the marker, tf. Only offspring scored for tufted at 28 days and later are included. Recombinant ex- ceptions were found in testcrosses of seven such alleles; but in the case of viable
TABLE 8
Tests for effects of viable alleles on recombination in T-tf interual, offspring scored f o r tufted after 28 days
Parents
Female Male non-tf Tailless tf
Oflspring
Bracliy
non-tf tf
Komral tail
non-tf t f
Reconibinants totals
42 1
97 9
8
170 17
10 1 4
15 2 62 3 10 52
4 98
292
46
2 96 25 85 6
4 35
1 w
1 6
1 14
4 65 63 9 120 7 17
14 6 15
25
53 1 46 not scored
8 86
53 14 1
not scored 10 2 31 not scored 3 21 66 15 not scored 1 /68 10/160
2/144) 12/337
14/185
o/
33151 541 24.1329 2/ 35J
3/ 2775 17/240
1/ 11) 4/38
2/ 29) 9/163 7/1341 19/134 11/229\ 30/383
o/
2001 0/3 13 O/ 801 0/474RECOMBINATION I N MICE 29 7 alleles, there is no direct test available to determine whether any of the re- combinants contain a t allele which is different from that in the parental line. At most, the qualitative conclusion can be drawn that the viable alleles t8, tis, tZ5, tW”, t2u24, tzua5 and tTu2? permit some recombination in the T-tf interval. All of these originated from exceptions found in balanced lethal lines in which the t allele was a suppressor of regular recombination. These derived alleles thus differ from the parent allele in at least two properties, those of lethality and of crossover suppression. Most of them differ also in male transmission ratio (Table 3 ) . The recombination fractions observed are shown in Table 9.
One viable allele, tW8. appears to suppress regular recombination, no recombi- nants having been detected in 474 test offspring observed. This allele was the only viable allele derived directly from a heterozygote found in a wild (Virginia) population. It is also the only viable allele tested which did not permit regular recombination. This may represent a difference in effect on recombination of alleles which have survived in the wild and have thus been subject to natural selection, but data on more “wild” viable alleles would be necessary to test such a relationship. I n addition, matings of T / t W 8 x T / t w B have produced fewer than the expected proportion of tW8 homozygotes. Until this complication is resolved, we can make no judgment concerning this exceptional association of a viable allele with suppression of recombination.
I n summary then: of 14 lethal alleles for which moderate or small amounts of test data exist, only two (and these may be the same allele) fail to suppress recombination in a region adjacent to T ; while of eight viable alleles tested only one (derived directly from the wild) suppressed recombination.
Exceptional recombinants: Exceptional recombinants are defined as ( 1 ) those arising in balanced lethal lines in which the t allele appears to suppress regular recombination in the T-tf region, and (2) those which can be shown to differ in their effects from the parent t allele. Such effects include viability of the derived allele when homozygous, or if the derived allele is itself lethal, it complements the parental lethal; and difference in male transmission ratio of the derived from the parental allele.
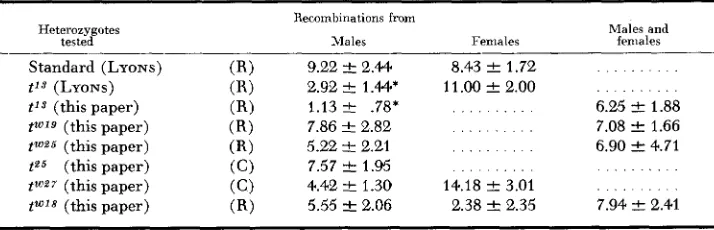
TABLE 9
Recombination percentages for the T-tf interual from Table 8 compared with values reported b y LYONS and PHILLIPS (1959). R , tests in repulsion (e.g. T tf/tn
+);
C , in couplingHeterozygotes tested
Recombinations from
Males and
Males Females females
Standard (LYONS) ( R ) 9.22 ‘-c 2.44 8.43 ? 1.72 . . .
t’3 (LYONS) ( R ) 2.92
*
l.44* 11.00 2 2.00 . . . . .t’3 (this paper) (RI
tP5 (this paper) (C) 7.57 & 1.95 . . . . . . . . . .
tW27 (this paper) (C) 4.42 k 1.30 14.18 f 3.01 . . . . . . .
1.13 k .78’ . . . . . . . . 6.25
-+
1.88t w 1 9 (this paper) ( R ) 7.86 k 2.82 . . . . . . . . 7.08 2 1.66 twa5 (this paper) (RI 5.22 & 2.21 . . . . . . . 6.90 2 4.71
tWJ8 (this paper) ( R ) 5.55 k 2.06 2.38 -C 2.35 7.94
-+
2.41298 L. c . D U N N , et al.
Such exceptions were first interpreted as exceptional recombinants by LYON and PHILLIPS (1959) who observed that tG, a lethal indistinguishable from to, suppresses recombination in the interval between T and the break point of trans- location T-190, near to t f . A recombinant from T-I90 t6/+ T x
+/+
proved to be+
t"/++.
2'' (now called t h ' ) being a viable allele and thus different from t6. They suggested as one interpretation that the change in the t allele from tG to t"' resulted from recombination in the region covered by the suppressing effect of t G ; and that exceptions in balanced lethal lines might thus represent exceptional recombinants. LYON (1960) later reported four recombinants tG +/tn tf and four T tf/t" tf from matings of T tf/t6+
x T tf/i6+;
in all eight cases, the t r allele in the recombinants proved to be viable and Ihus different from t G . A ninth exception contained a new t allele but was not a recombinant. T h e frequency of the proved recombinsnts-eight in a population at birth of 3768-is similar to the frequency of exceptions from balanced lethal lines reported in this paperWe have now analysed seven further exceptions from tests of other lethal t alleles as follows (cf. list in Table 3 ) .
tI2 + / t Z 5 tf from T t f / t l 2
+
X T t f / t Z 2+
T if/tZ6 tf from T t f / t '+
x T tf/t'+
to + / t Z 7 tf from T t f / t o+
x T tf/to+
t' + / t Z R tf from T tf/P f X T t f / t l+
T t f / t I c z 7 tf from+
tf / P o 4 X T tf/+ tf t7U'X +/ttbZ9 tf from T t f / t Z " ' *+
X T i f / t 1 ( ' 8 ft 2 u ' ~ + / t u P o tf from T i f / P 2
+
X T tf tibZ2+
One other exception iZb?*, a lethal, was detected under circumstances which did not permit a decision whether its origin preceded or followed the cross by which the tf marker was added to the balanced lethal stock, T/t'"'O. It is therefore not included in the following discussion.
I n all cases in which the exception arose from a parent heterozygous for a lethal t allele and the tf marker, the exception contained a new t allele and the marker. I n the first six of these cases, the new t allele was viable, in the seventh
( ti( ?(') it was lethal but complementary with the parental lethal.
It is clear that in our cases, as in LYON'S, change in the form of a lethal t allele usually accompanies exceptional recombination. Usually the change is from a lethal to a viable allele but in one of our cases it was from a lethal to a different lethal.
The association of the new allele with the marker ( t f ) from the homologous chromosome of the parent could of courss be due to mutation of T 4 t" or
+
+ t'R E C O M B I N A T I O N IN M I C E 299 but did not alter the property by which t alleles are recognized, namely interac- tion with T to produce a tailless phenotype.
I n the two cases (t"', tzU") tested for recombination suppression the new viable allele arising by recombination had also lost the recombination suppressing effect
of the parental allele. Five other viable alleles which arose from exceptions before the tf marker was inserted (and are thus not proved to have arisen in re- combinants) were also shown to have lost the suppressing effect (Table 8). The effect of the lethal recombinant ( t Z U 3 0 t f ) on recombination suppression has not
yet been tested.
Of five lethals previously derived from suppressor alleles, four ( P , t Z 0 , tto2', ttU2') probably suppress recombination, one ( tWIU) does not.
Six of the seven new alleles found in exceptional recombinants have also been tested for effect on the male transmission ratio. Five of these ( t z 5 , tZ6, tes, tZo27, t""") which had arisen from the LLhigh-ratio" alleles t l Z , t', t O , ttcSo and t'c1'2 re- spectively proved to have significantly lower male transmission ratios than the parental allele. The allele tZUzg which arose from t W 1 8 (male transmission ratio
about 0.6) has a slightly lower ratio ( 0 . 5 3 ) . I n addition, tests of 19 alleles which arose from exceptions from high-ratio alleles before the marker was added showed that in all 12 viable alleles studied the male transmission ratio was normal ( 0 . 5 ) or low ( 0 . 3 - 0 . 4 ) ; two of the new lethals had low ratios (t4--0.4; t9-0.45) while four retained the high ratio of the parental allele; in one case
( P I 8 ) the ratio was above normal but significantly lower than that of the
parental allele.
It is thus evident that of the several properties of t alleles which have been studied, one, the interaction effect with T is not altered when the form of the allele changes, probably as a result of recombination. The lethal effect may be lost, or retained in a different form and is thus separable from the interaction effect. The recombination suppressing effect has been tested for seven viable alleles derived from exceptions and was lost together with the lethal effect in all seven, whereas this effect is usually retained when the derived allele is lethal. But in one case, ( t Z u I 8 ) the lethal effect was retained, and the suppressing effect lost, showing that these properties are also separable. This case is of special interest since an exceptional recombinant ( tZUz9 t f ) was detected from T tf/tZGlu
+
showing that recombination suppression is not an essential condition for the occurrence of exceptional recombination.The effect on male transmission ratio is separable from the other effects and is usually altered by the event which results in an exception, always when the lethal effect is lost, less often (three cases out of eight) when the derived allele is also lethal. I t must be remembered, however, that we have detected exceptions only from lethal alleles having high ratios. The "low-ratio" lethals t4 and P o
have not yet been extensively studied.
300 L. c . D U N N , et al.
lethal which suppresses regular recombination in this region. The exceptional recombination event so alters some of the properties of the parental allele that a recognizably different t allele is recovered in the recombinant chromosome. The fact that certain other effects of t alleles, such as lethality, recombination suppres- sion and male transmission ratio are separable from the interaction effect with T by which t alleles are identified suggests that the other effects are those of a region rather than of a single locus. The hypothesis proposed by LYON (1960) to explain the relation of t6 to this region appears to apply to other t lethals which are also suppressors of regular recombination, namely that they are chromosomal aberrations adjacent to but not unilocal with T . When a recombination break occurs within them. new and different arrangements arise with effects which differ from those of the parental aberration.
DISCUSSION
The first point to be emphasized is the lack of any evidence of recombination between the original mutant T and the interaction effect of the numerous t alleles which have been studied. It is in this sense that T and this one property of t alleles can be regarded as allelic.
Secondly, exceptional gametes produced by heterozygotes T/tn or +/t" (t'l being any one of eight different lethals) have been shown in each of 28 cases analysed by us and in nine cases analysed by LYON and PHILLIPS (1959) and LYON (1 960) to carry a newly arisen t" allele differing from the parental form, t". These have been shown not to represent recombinants between T and t". In all seven cases analysed by us in which recombination in a marked region adjacent to T could be detected and in all of LYON'S similar cases the origin of the new tx allele was accompanied by recombination in this region, and this appears to be the usual mode of origin of new t alleles. New alleles arising in this way include a variety of different combinations of effects-lethal or viable, suppressors or nonsuppressors of recombination, high, low or normal male trans- mission ratio. These differences have been ascribed to different alterations in the region adjacent to T .
It has been suggested previously (DUNN 1956) that the t lethals which sup- press recombination may be properties of inversions in the vicinity of T locus. The new evidence that changes from lethal alleles which suppress normal recom- bination to viable alleles which permit such recombination are accompanied by recombination on one side of T would require a series of spatially separated inversions each associated with a different t lethal. These could be separated by rare acts of crossing-over by which the normal gene order of the homologous chromosome could be combined with the locus controlling the t-interaction effect. Lethality would be an inherent property of each such inversion and comple- mentarity of different lethals in compound would be assumed to occur because spatially different inversions did not overlap.
RECOMBINATION IN M I C E 301
sections which suppress recombination in regions adjacent to them. The homozy- gous lethal effect of each t lethal would be due to homozygous deficiency and complementarity between different lethals to possession by such compounds of one complete set of normal genes (nonoverlapping deficiencies). Occasional indi- cations of partial complementarity between different t lethals (as in the case of
t 1 / t l Z , DUNN 1956) might then be due to partial overlapping of adjacent deficien- cies.
Both hypotheses assume that a long region of chromosome is subject to the occurrence of such aberrations because of the known minimal length of the region over which suppression of recombination extends. Any aberration in this region must then show close linkage to the locus, T , at which the interaction effect on the tail is determined; and the crossing-over event which separates the locus T from the known marker on one side must occur between the aberration and the locus of T . It is unlikely that the aberrations in any of the lethals detected from exceptions include the locus of T which as a lethal itself would probably be inviable in compound with a deficiency which includes its locus, whereas compounds of T with t lethals are viable. Changes to viable alleles which are also nonsuppressors would on this hypothesis involve loss by recombination of the deleted or otherwise aberrant segment.
From present evidence discrimination between inversion and deficiency cannot be made and must in any event await cytogenetic study. There is at present no information on the effects of either type of aberration on recombination in mam- malian chromosomes. If the effects follow the pattern found in Drosophila and maize, recombination suppression if due to inversion should be accompanied by formation of bridges and acentric fragments arising from crossing-over within the inversion loop and these should give rise to inviable zygotes; while if due to deficiency should lead to local failure of pairing between homologues one or both of which contain deficiencies, and to shortening of the linkage map of the region by at least the length of the deficient segment or segments. The inversion hy- pothesis in the above form assumes that the lethal effect is a consequence of the inverted order, and that each inversion (of which several must be assumed to account for the diversity of t lethals) has this effect; whereas the deficiency hypothesis accounts for lethality more simply by absence of genes in the deficient section.
One consequence of thinking of the region on one side of T as consisting of a
number of segments between which breakage and recombination may occur, is that if breaks recur at the same point then duplicate recombination products should be found. That is to say, some alleles arising by this process should be indistinguishable from each other. At present there is no direct test of identity or nonidentity between viable alleles which have arisen from the same lethal, but methods for such a comparison are being developed. Two lethals twzo and
twzz which arose successively from the same parental allele (of the tW5 group)
302 L. c . DUNN, et al.
The processes which gave rise to the first variant alleles at this locus are of course unknown but we may now put some limits to speculation about them. The origin of T may be ascribed to mutation in the narrow sense; there is no
evidence that it involves any chromosomal aberration. The first t alleles ( t o and t ' ) had occurred in normal stock (in the wild in one case) before a cross with stock carrying T brought them into the balanced lethal combination in which they were identified. They must have had from their origin some transmission ratio advantage in order to have persisted under the selection pressure against homozygous lethals. The same is true for the numerous cases in which t alleles have been found in natural and in laboratory populations. If the initial event producing the first t allele was the occurrence of an aberration such as deficiency or inversion, then subsequent ones could arise from it by events connected with rare acts of recombination, as has been observed. The first event. whatever it was. can be thought of as initiating an instability which leads a given t allele to produce probably by exceptional recombination, a succession of derived t alleles with different properties.
SUM MARY
The genetic relationships of a series of mutations at locus T have been exam- ined in breeding experiments.
Combinations of T (a lethal) with other members of the series ( t alleles) yield a tailless phenotype. When the t allele is lethal, matings T / t X T / t constitute a balanced lethal system. In general such matings produce only tailless offspring at birth but 46 living exceptions with normal tails, out of 25,751 were observed. All 28 exceptions analysed proved to be tn/tx, tn being the parental t allele, tx a
new allele. No instance of recombination between T and any t allele was found either in the above or in 4399 offspring from T / t X T / t in which the t allele was viable.
By use of a marker gene ( t f ) near locus T , it was shown that in general lethal t alleles (with one exception) suppress recombination in the T-tf interval, (8-10 crossover units). Seven exceptional off spring arising from such marked balanced lethal lines ( T tf/t" +) or from
+
tf/tn+
were t" + / t r t f , that is, recombination between T and tf was involved in the origin of a new t allele which in six of these cases was viable, in one case lethal. Seven viable t alleles appear to be nonsup- pressors of recombination in the T-tf interval.The hypothesis is discussed that lethal t alleles which are also suppressors of
R E C O M B I N A T I O N I N M I C E 303 L I T E R A T U R E C I T E D
BENNETT, DOROTHFA, and L. C. DUNN, 1960
BENNETT, D., L. C. DUNN, and S. BADENHAUSEN, 1959
CARTER, T. C., and R. J. S. PHILLIPS, 1950
A lethal mutant (tW18) in the house mouse
The embryological effects of four late
Three recurrences of mutants in the house mouse. showing partial duplications. J. Exptl. Zool. 143: 203-219.
lethal 2-alleles in the mouse. J. Morphol. 105: 105-143.
J. Heredity 41 : 252. CHESLEY, P., 1935
70: 429-459.
CHESLEY, P., and L. C. DUNN, 1936
DOBROVOLSKAI.4-ZAWADSKAIA, N., 1927
Development of the short-tailed mutant in the house mouse. J. Exptl. Zool.
The inheritance of taillessness (anury) i n the house mouse.
Sur la mortification spontanbe de la queue chez la
souris nouveau-n6e et sur l'existence d'un caracti're (facteur hCrPditaire) non-viable. C.R. Soc. de Biol. 97: 114-119.
DOBROVOLSKAIA-ZAWADSKAIA, N., and N. KOBOZIEFF, 1932 Les souris anoures et la queue filiforme qui se reproduisent entre elles sans disjunction. C.R. Soc. Biol. 110: 782-784.
DUNN, L. C., 1952 Genetically determined variations in male fertility i n the house mouse. Edited by E. ENGLE. Studies on Testis and Ovary, Egg and Sperm. C. C Thomas, Springfield, Illinois.
Analysis of a complex gene in the house mouse. Cold Spring Harbor Symposia Quant. Biol. 21 : 187-195.
Studies of the genetic variability in populations of wild house mice. 11. Analysis of eight additional alleles at locus T. Genetics 42 : 299-31 1.
Variations i n the transmission ratios of alleles through egg and sperm in M u s musculus.
Am. Naturalist 94: 385-393.
A comparison of the effects, in compounds, of seven genetically similar lethal t alleles from populations of wild house mice. Genetics
45: 1531-1538.
A case of neighboring loci with similar effects. Genetics
Tests for recombination amongst Genetics 21 : 525-536.
1956
1957
1960
DUNN, L. C., and DOROTHEA BENNETT, 1960
DUNN, L. C., and E. CASPARI, 1945
DUNN, L. C., and S. GLUECKSOHN-SCHOENHEIMER, 1943
30: 543-568.
three lethal mutations in the house mouse. Genetics 28: 2 9 4 0 .
DUNN, L. C., and S. GLUECKSOHN-WAELSCH, 1953 The failure of a 2-allele ( P ) to suppress crossing over in the mouse. Genetics 38: 512-517.
DUNN, L. C., and J. SUCKLING, 1956 Studies of the genetic variability i n wild populations of house mice. I. Analysis of seven alleles at locus T. Genetics 41 : 344-352.
LYON, M. F., 1956 Hereditary hair loss in the tufted mutant of the house mouse. J. Heredity
47: 101-103.
1960 Effect of X-rays on the mutation of t-alleles in the mouse. Heredity 14: 247-252.
LYON, M. F., and RITA J. S. PHILLIPS, 1959 Crossing over in mice heterozygous for t-alleles. Heredity 13: 23-32.
PONTECORVO, G., 1958
SILAGI, SELMA, 1962
Trends in Genetic Analysis. Columbia Univ. Press. New York.