Exogenous
substrate
stimulates
autodephosphorylation
of
cyclic-AMP-dependent protein
kinase 11
Bj0rn T.
GJERTSEN, Beathe FAUSKE
and
Stein
0.D0SKELAND*
Cell Biology Research Group, Department of AnatomyandCell Biology, UniversityofBergen,Arstadveien 19, N-5009Bergen, Norway
The autophosphorylated regulatory subunit (32P-RII) of
cyclic-AMP-dependent protein kinase II was efficiently
de-phosphorylatedby its C subunit in the absence of added ADP, provided that Mg/ATP and a standard protein kinase peptide
substrate were present. This raises the possibility that
auto-dephosphorylationcould be significant in the intact cell. Only the
cyclic-AMP-complexed free form of 32P-RII was efficiently
dephosphorylated, indicating that the autodephosphorylation
was intermolecular. Autodephosphorylation of 32P-RII in the
INTRODUCTION
Enzymescaninprinciple catalyseareaction inboth theforward
andthereversedirection. This is also thecaseforthecyclic-AMP
(cAMP)-dependentprotein kinase (cA-PK), which catalyses the reversible reaction:
MgATP+protein=P-protein+MgADP (1)
Thisenzymeexists in twomain isoenzyme forms, cA-PKI and cA-PKII, and hasa pivotalrole in mediating the effects of the secondmessengercAMP byphosphorylationofkey cellproteins.
The type II isoenzyme undergoes an autophosphorylation
re-action,whereaspecific serine residue in the'hinge'region of the
regulatory moiety (RII) is phosphorylated by the catalytic (C) moiety. For general reviews see Beebe and Corbin (1986),
Edelmanetal.(1987), Tayloretal.(1990), McKnight(1991)and D0skelandetal. (1993).
The protein-kinase-catalysed reaction (eqn. 1) is considered virtually irreversible within the cell, because the forward reaction is faster thanthereverse reaction (Shizuta et al., 1975; Rosen and Erlichmann, 1975; D0skeland et al., 1984), and because of the low proportion of intracellular MgADP relative to
MgATP.
The present study reports the novel finding that, for
auto-phosphorylated RII, reversal of the kinase reaction did not require added ADP,provided that the Csubunitwas supplied
with standard peptide substrate and MgATP. This raises the
possibility that thereactioncanbe ofsignificance in the intact cell,i.e.evenwhenMgATPpredominatesrelativetoMgADP. A mechanism is proposed for the coupling between
auto-dephosphorylation of P-RII and phosphorylation of peptide substrate, involving a transient C-MgADP complex formed during C-catalysed phosphorylation of standard peptide substrates.
presence ofMgATP and kemptideoccurred withformation of
[y-32P]ATP, suggesting transfer of 32p of phospho-RII to a
transient C*(MgADP) complex formed during the forward kinase reaction with peptide as substrate.
Autodephosphoryl-ation promoted by phosphorylation of exogenous substrates couldoperatealsofor other kinasesconformingtoamechanism
whereMgADP remains boundto the active site after the other product(phosphorylatedsubstrate) has left thecatalytic complex.
EXPERIMENTAL
Materials
[y-32P]ATP and [5',8-3H]cAMP were from The Radiochemical
Centre, Amersham, Bucks., U.K. ATP-regenerating system
(phosphocreatine and creatine kinase) was from Boehringer,
Mannheim, Germany. Microcystin-LR, phosphate-acceptor heptapeptide (Leu-Arg-Arg-Ala-Ser-Leu-Gly, 'kemptide') and all unlabelled nucleotides were from Sigma, St. Louis, MO,
U.S.A. Reagents for the Bradford (1976) protein assay and a
mixed globulin protein standard wereobtained from Bio-Rad,
Richmond, CA, U.S.A. A synthetic peptide corresponding to residues 5-24 of the heat-stable cA-PK inhibitor protein (Scott et al., 1986) was generously given by Dr. J.Scott and Dr.E.G.Krebs, University of Washington, Seattle,WA, U.S.A. C subunit and cA-PKIIwerepreparedfrom bovinemyocardium,
essentiallyas describedbySugdenetal.(1976)and Ogreidand D0skeland (1980), respectively.
Determination of
[H]cAMP
binding
to R; assayforkemptide
Determination of[3H]cAMPbindingtoRIorRIIwasbasedon
measurement of their binding capacity for [3H]cAMP and
separation by using specific antibodies (Ekangeret al., 1989). The C-subunitconcentration was estimated by mixing various
concentrations withpurified RIof knownconcentration (in the absenceofcAMP)andtitrating theresultingfree kinaseactivity by assaying its phosphotransferase activity at 30°C in 50mMpotassiumphosphate/15mMHepes/1OmMmagnesium
acetate/70,M phospho-acceptor peptide (kemptide)/0. mM
[y-32P]ATP (2mCi/mmol)/ 0.5 mM EGTA /1 mM dithio-erythritol.Thisestimatewas15%lower than the estimate based on protein content and Mr=40000, suggesting either that the
Abbreviations used:cAMP,cyclic AMP; cA-PKI, cA-PKII,cAMP-dependent proteinkinasetype andtype11respectively;RI, RII,theregulatorysubunits ofcA-PKI and cA-PKII respectively; C,thecatalytic subunit ofcA-PKIor cA-PKII; kde, rateconstant forpseudo-first-order dephosphorylation; Kd,
apparent equilibrium dissociation constant; C(MgADP), C subunit with MgADP bound to its active site as a result of direct binding of MgADP; C*(MgADP), C subunit with bound MgADPformed insitufrom bound MgATP upon peptide phosphorylation; kemptide, thephosphate-acceptor
heptapeptide Leu-Arg-Arg-Ala-Ser-Leu-Gly.
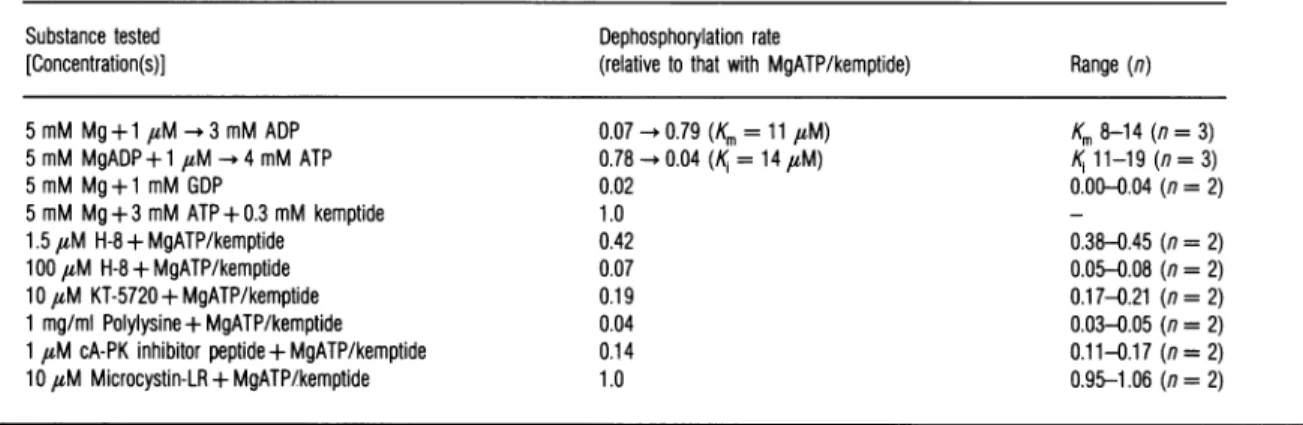
Table 1 Test of the
ability
of selected substances tomodulatetheC-subunit-catalysed
dephosphorylation
of32P-RIIPre-autophosphorylated32P-cA-PKIIwassubjected todephosphorylationin thepresence of the Csubunit of cA-PK and theagentsindicated inthe first column of the Table.Other conditionswere
asdescribed in theExperimental section. The initial rate ofdephosphorylationwasdetermined and isexpressedrelative to the rate observed in thepresence ofMgATPandkemptide.The apparent Km value for MgADP andthe K1 value ofMgATP (with30,uM or100,uM ADP)arealso shown.
Substance tested Dephosphorylation rate
[Concentration(s)] (relativetothat withMgATP/kemptide) Range(n)
5 mM Mg+1 uM-*3 mMADP 5 mM MgADP+1FM -4mMATP 5 mM Mg+ 1 mM GDP 5 mM Mg+3 mMATP+ 0.3 mM kemptide 1.5FM H-8 +MgATP/kemptide 100 FM H-8 + MgATP/kemptide 10FM KT-5720 + MgATP/kemptide 1 mg/mlPolylysine +MgATP/kemptide 1FM cA-PK inhibitor peptide + MgATP/kemptide 10FM Microcystin-LR +MgATP/kemptide
protein standard was not accurate for the C subunit when the Bradford (1976)assay was used,or thatsomeofthe C-subunit
protein wascatalytically inactive.
Theamount ofphospho-acceptor peptide (kemptide)present
in a sample was estimated byallowing the phosphorylation to
proceed to apparent completion. The assay was at 30°C in 50 mM potassium phosphate / 15 mM Hepes / 10 mM
mag-nesium acetate/ 0.3 mM
[y-32P]ATP
(1 mCi/mmol) /0.5 mMEGTA
/0.1
mMEDTA/l
mMdithioerythritol/100
nM C sub-unit.Sampleswereremoved after20,30and40minofincubation. Completeness of substrate phosphorylation wasindicatedby
aplateau ofincorporation asa function of incubation time.
Assay for
autodephosphorylatlon
of cA-PKII
ThecA-PKIIholoenzymewasroutinely autophosphorylated by preincubation for 15min at 0°C in 15 mM Hepes/NaOH,
pH7.2,
containing
20mMKCI, 1 mMEDTA,0.3 mMEGTA,0.3mg/ml BSA and 0.15mg/ml of soybean trypsin inhibitor, with 2.5 mM magnesium acetate and 2
,uM [y-32P]ATP
(60Ci/mmol) ina volume of0.25ml. Excess radioisotope was
removed by passing the preincubation mixture through a
Sephadex
G-25 column (1.2ml bedvolume)
equilibrated with50 mM potassium phosphate (pH 7.2)/l mM sodium
pyro-phosphate/20mMNaF/5mMEDTA/5mMEGTA. Fractions (60
Fl)
werecollected, andthe three firstflow-through fractionswere pooled. The amount of32P-RII recovered was the same
whether or not
2,uM
microcystin was included duringpre-incubation andinthecolumn-equilibration buffer.
The
dephosphorylation
reactionwasstartedbyadding20Fl
of thepooled 32P-labelled
cA-PKII fractions to 1.3 ml ofbuffer
(15
mM Hepes/NaOH, pH 7.2, 20 mM KCI, 1 mM EDTA, 0.3 mM EGTA, 0.3mg/ml of BSA, 0.15mg/ml of soybean trypsininhibitor)
containing routinely6 mMmagnesiumacetate, 30FM
cAMP and 50-100 nMof thecatalytic subunit ofcA-PK. Variousconcentrations ofADP, ATP, ATP-regeneratingsystem(phosphocreatine and creatine kinase) and other agents to be tested (Table 1) were present in some incubations. The
de-phosphorylation reactionwasarrested by spotting150
Ful
sampleson to
filter-paper
discs(25
mmdiameter),
which wereimmedi-ately
transferredto 10% (w/v)trichlor-oacetic
acidwith 5 mMpyrophosphate. After washing
(10
ml/filter) six times in 5%trichloroacetic acid, the filters were treated sequentially with
0.07 -0.79(Km=11 FM) 0.78 -0.04(K=14FM) 0.02 1.0 0.42 0.07 0.19 0.04 0.14 1.0 Km8-14(n= 3)
Ki
11-19(n=3) 0.00-0.04 (n =2) 0.38-0.45 (n=2) 0.05-0.08 (n=2) 0.17-0.21 (n =2) 0.03-0.05 (n=2) 0.11-0.17 (n=2) 0.95-1.06 (n=2)ethanol and acetone, dried, and counted for radioactivity in a
liquid-scintillation counter.
Determination of
pseudo-first-order rate
constantsfor
dephosphorylafton of 32P-RII:
estimat!On
of the fractional
occupancy
of thebinding
sitegoverning
the cAMP effect onautodephosphorylation
When the initial rate ofdephosphorylation of 32P-RII obeyed
apparentfirst-orderkinetics, the operationalrate constant
(kde.)
for the dephosphorylation process was calculated
simply
from the relation:kde=ln2/tl
where
tL
is the time (in seconds) required for 50%dephosphorylation of the phosphoprotein
initially
present.Assuming that cAMP accelerated the dephosphorylation of
P-RII by binding to a site, and acted proportionally with the fractional occupancyof thissite, it follows that:
Fractionaloccupancy=
[kde(x)
-kde(0)/[kde(max.
cA)
-kde(0)]
where kde(O) is the rate constant for dephosphorylation in the absence of added cAMP, kde(x) that in the presence ofcon-centration x of cAMP, and kde(max.cA) that at maximally effective cAMP concentration.Asimilarapproach has been used
to estimate the occupancy of one binding site governing the
behaviourof anotherbinding site (D0skelandetal., 1987).
Determination of
[nP]ATP
produced
during
the
autodephosphorylatlon
of
32P-RII
Pre-autophosphorylation
was asdescribedabove,
exceptthat theconcentration of cA-PKII wasincreased 5-fold and thespecific radioactivityof[y-32P]ATPwasincreased2-fold.[y-32P]ATPwas removed from the preincubation by passing it twice through SephadexG-25 columns, each twice the routine size. Since the desalted preincubation was diluted only 3-fold in the
de-phosphorylation reaction, the second columnwas equilibrated
withonly5mMNaF,5mMpyrophosphateand 2 mM each of EGTAandEDTA, toavoid excessive chelation of
Mg2+ during
the
ensuing autodephosphorylation
reaction. Less than 2% of the32p
inthe sample subjected toautodephosphorylation
wassolublein0.6-M HCl04.
C subunit, 300,M kemptide and 0.5 mM ATP. The ATP-regenerating system and other ingredients were as described above. The reaction was also performed in the absence of kemptide, ATP or ATP-regenerating system, both with and without 3 mM ADP. In one series of experiments the
auto-dephosphorylation wascompared between the incubations con-taining MgADP + complete ATP-regenerating system and those
containing MgADP with phosphocreatine and boiled creatine
kinase. The reactions were arrested by addition ofHCl04 and
separated by h.p.l.c. essentiallyasdescribed by D0skeland et al. (1992), by using a LiChrosorb RP-18 UltroPac column (4 mm x 250 mm) with 2
%
(v/v) methanol, 110mMpotassiumphosphate(pH 6.9)and0.3 mMtetrabutylammoniumhydrogen
sulphate as the mobile phase. Fractions of volume 0.3 ml were
collected and assayed for radioactivity.
To test whether the labelled substance co-chromatographing
withauthentic ATPwas[y-32P]ATP,the peak h.p.l.c.fractionsof presumed [32P]ATP were tested for ability to transfer 32P to
peptide substrate under standard protein kinaseassay conditions
with 0.1 mM ATP, 150 nM C subunit and 300
,#M
kemptide.The reaction was allowed to run to apparent completion, as
ascertained bya plateau ofincorporationof32P into substrate
(achieved after less than 20 min of incubation). The degree of incorporation of 32P from [32P]ATP produced in the
auto-dephosphorylation reaction was compared with the
incorpor-ation using as phosphate donor [y-32P]ATP freshly obtained from the producer, and dilutedinh.p.l.c. mobile phase.
Autoradlography
of cA-PKII autophosphorylated in the absence
and
presenceof
kemptide and cAMP
cA-PKII holoenzyme (0.1 ,uM with respect to subunit
con-centration) was incubated at 0°C in 15mM Hepes/NaOH,
pH 7.2, 20 mM KCl, 1 mMEDTA, 0.3 mM EGTA, 0.3 mg/ml
BSA,0.15mg/ml soybean trypsin inhibitor,2.5 mMmagnesium acetateand4,uM[y-32P]ATP (60 Ci/mmol).Insomeincubations
cAMP(30 ,uM) orsubstrate peptide (300
4uM)
waspresent. The incubation volume was 100,ll.
Samples of volume 20 ,ul wereremoved, mixed first with 0.5 ml of
100%
trichloroacetic acid,and then with5
/ug
each ofBSA, ovalbuminandsoybean trypsin inhibitor. The precipitate was collected by centrifugation, extracted with diethyl ether, and dissolved in 40,ul of10 mM Tris/phosphate, pH 8, with 9 M urea, 2 mM dithioerythritol and 1mMethylamine. This sample was diluted 1:1 insample buffer and subjected to SDS/PAGE (110%
total acrylamideconcen-tration) as described by Laemmli (1970). The washed and
subsequently dried gel was exposed to radiographic film
(/3-MAXfromAmersham).
RESULTS
Introductory experiments
The RIIsubunit of cA-PKII showedananomalous decrease in
its apparent cAMP affinity as a function of incubation time in tissue extracts supplemented with MgATP. It was considered
possible that this was due to loss of phosphate from the
autophosphorylation site of RII. However, neither inhibitors directed against phosphatases 1 and 2A (microcystin) or the
Ca2+/calmodulin-dependent phosphatase 2B (EGTA, W-7 and
trifluoperazine)norNaFpreventedthe decrease ofcAMPbinding
to RII [the nomenclature and inhibitor-sensitivity of protein phosphatases has been reviewed by Cohen
(1989)
and Bollen and Stalmans(1992)].
In a further attempt to inhibitphosphatases, the cA-PK substrate kemptide (500
1M)
was added to achieve anincreasing concentration ofthe presumedphosphatase inhibitor phosphokemptide. Surprisingly, this further decreased the cAMP binding to RII. Decreased apparent cAMP affinity was also noted when kemptide was presented to purified cA-PKII incubated with MgATP. These findings led us to suspect that kemptide and MgATP enhanced the dephosphorylation of P-RII independently of phosphatases. This possibility was studied by using isolated cA-PKII.
The
C
subunitof cA-PK catalyses dephosphorylation
of P-RII
Inthe
presenceof saturating MgATP
and substrate
peptide
Autophosphorylated 32P-RII(cAMP)2 was indeed rapidly
dephosphorylated in the presence of kemptide, MgATP and
the C subunitof cA-PK (Figure 1). The initial reaction obeyed
apparentfirst-order kinetics (Figurela), and thefirst-orderrate constant for dephosphorylation was similar for all
32P-RII-(cAMP)2 concentrations (1-50 nM) tested (resultsnotshown),as
expectedfor a reaction in which the substrate concentration is far
belowits Km value. Thereactionwasnotsupportedby phospho-kemptide (Figure la), and requiredMg2+, andwasdependenton the C-subunit concentration (Figure lb). The effect of the C
, 0.25 ._-._ E Ca 0.1 cL o 1.0 c 0 0 LL. 0 1 2
Time ofincubation (min)
Figure 1 Dephosphorylation of32P-RII is catalysed by the C subunit of cA-PK in thepresenceofMgATPand kemptide
Panel(a) shows thedephosphorylationof32P-RII byC subunit(60 nM) in the presenceof kemptide (0.3 mM), with (U) or without (0) an ATP-regenerating system (4 mM phosphocreatine+10 units/ml creatinekinase;Cr-P/CPK). With 50FM kemptide(A) the
dephosphorylation rapidly abated. Phosphokemptide (O>) failed to support rapid dephosphorylation. Panel (b)shows the Mg2+requirement and the C-subunit concentration-dependence of thereaction.The cA-PKIIholoenzymewaspre-autophosphorylatedasdescribed
in the Experimental section, and 20,1 (containing 2.5pmol of 32P-RII) was exposed to
dephosphorylation by mixing with 1.3ml of buffer(15mMHepes/NaOH pH7.2,20mMKCI,
1mMEDTA,0.3mMEGTA, 0.3 mg/mlBSA,0.15mg/mlsoybean trypsininhibitor) containing
10 mMmagnesium acetate, 3 mM ATP and 30FMcAMP. Inoneexperiment(El)magnesium
acetatewasomitted. For theexperimentsshownin(b)the buffer contained30 nM(A),90 nM
(O)or no(0)added Csubunit. About2 nMC subunitwascarriedoverwith thepreincubation
mixture. Samples (150Ful) were spottedon to filter-paper discs todetermine 32P-RII, as
subunit depended on its concentration, rather than the ratio
between P-RII andCsubunit concentrations(tested from 1:1 to 1:200), indicating a catalytic effect of the Csubunit,as expected
whenexcess cAMP is present to prevent 'trapping' of C subunit incA-PKIIholoenzyme complex.
The dephosphorylation was dependent on continuous phosphorylation of kemptide, since the reaction rate declined as
the amount of kemptide became insufficient to support the
kinase activity (Figure la). This was demonstrated by starting
the reactionwith aconcentration of kemptide (50,uM) that, at
the level of C subunit present, became nearly completely
con-verted into phospho-kemptide during the reaction period. The conversion of kemptide intophospho-kemptidewasdetermined in aseparate experiment run under similar conditions to those
shown inFigure
1(a)
with 50,uM kemptide and using cA-PKII holoenzyme preincubated without labelledATP. Samples werediluted4-fold inaprotein kinaseassaywith200,tM[y-32P]ATP and 100 nM C subunit for 15min, and 32P-kemptide was
determined.Only20
%
of theoriginally added 50ptM
kemptidewasstillphosphorylatable after30 sofincubation, and lessthan 5
%
was left after60 s(resultsnotshown).The need for catalytic activity of the C subunitwassupported bythe lower rateofautodephosphorylation in the presence of kinaseinhibitors (Table 1), such as apeptidecorresponding to
the active sequence of the heat-stable protein kinase inhibitor,
and the ATP-directed inhibitors H-8 and KT-5720 (reviewed by Hidaka and Kobayashi, 1992). The abolition of
auto-dephosphorylation by polylysine (Table 1)was presumably due
to blocking ofthe interaction between RII and the C subunit, sincepolylysineprevented the reassociation ofRIIand Csubunit
(results notshown).
Evidence that
MgADP, formed
insitu
onthe
C subunit from
MgATP,
is
thephosphate acceptor
during
autodephosphorylation
of cA-PKII
The molecule accepting the 32P from
32P-RI(cAMP)2
duringautodephosphorylation
in the presence of kemptide, MgATP andanATP-regenerating
system wassoughtby h.p.l.c. analysis of theHCl14-soluble
fraction from such anincubation. More than 90%
of theHCl14-soluble
32pwasidentified as[32P]ATP. About75%
ofthis[32P]ATP could be incorporatedinto kemptideunderprotein kinaseassayconditions, where70-75
%
of authen-tic[y-32P]ATP
was incorporated.[y-32P]ATP
was therefore the main, and possibly the only, radioactive product formed from 32P-RII duringautodephosphorylation. The origin of the ADPrequiredtoacceptthelabelledphosphate from 32P-RIItoproduce
[y-32P]ATP
was not immediately evident, since no exogenous ADPhad been addedtotheincubation.Furthermore,an efficient (seebelow and Figure 2) ATP-regenerating system was present, and any free ADP molecules would have had to compete (Table 1, Figure 2) with a vast excess ofATP in the reaction mixture. TheacceptorADP therefore hadtobepresent at asite inaccessible to theATP-regenerating system and not in
equilibrium with thegeneralATPpoolinthe incubation. Theexperimentsshown inFigure2 wereperformedtofind to what extent MgADP, added to the reaction medium, could substitute for the ADP produced in situ (see above). One would expect that onlya fraction
([C*(MgADP)]/[total
C]) of the C-subunit molecules carried MgADP during kemptidephosphorylationinthe presence ofMgATP(seeright-handside
of Table2).Therefore, dependingonthesize ofthisfraction, the autodephosphorylation with MgATP and kemptide should be slower than in the presenceofmillimolarMgADP,which should
nearlysaturatetheCsubunitwith MgADP. Surprisingly, when
c1.0 X0.5 A d. N~~~~~~ .I4- uu0 o 0.25 fl0 3mMATP+Cr-P/cPK
0A3mM ATP +0.3mMADP 3
3mMATP+3mM ADP
_ A 3mM ADP
0.1 - 0 3 mM ADP+
Cr-P/CPK
1 2
Timeofincubation(min)
Figure2 MgADP, but not MgATP, can promote
autodephosphorylatlon
in
theabsenceof
kempfide
The timecourseof32P-RIIdephosphorylation wasstudied in the presence of 10 mM Mg2+, 30uM cAMP and 75 nM Csubunit. The rateofdephosphorylation was low without added nucleotide (0) as well as with 3 mM ATP (E). With 3 mM ADP (A) alone the
dephosphorylation was about twice as fastas when equimolar (3 mM ADP/3 mM ATP)
amountsof ADP and ATP (R)were present. Theeffect of ADP (0.3mM) was efficiently counteractedby3 mM ATP(A),and eventheeffect of 3 mM ADPwasabolished(i) by
an ATP-regeneratingsystem(Cr-P/CPK,asin Figure 1).
the
dephosphorylation
ratewascompared
inincubationsruninparallel
withMgADP
andwithMgATP-kemptide,
itwasslightly
(20
%),
butsignificantly,
faster under the latter conditions(Table
1). The effect of ADP(kemptide
absent)
wascompletely
blocked
by
anATP-regenerating
system(phosphocreatine/
creatinekinase),
andwascompetitively
inhibitedby
ATP(Figure
2, Table1).
The apparent Km
(11 ,uM)
forMgADP
in theauto-dephosphorylation
reaction(Table 1)
was close to the K1 ofMgADP
estimated from theability
ofMgADP
to inhibitcompetitively cA-PK-catalysed
phosphorylation
of substrates(Whitehouse
etal., 1983;
results notshown).
A lowKm
forMgADP
wasalsonoted in theearly
study
ofreversal of 32P-R11phosphorylation by
Rosenand Erlichmann(1975).
On theotherhand,
a Km value forMgADP
of 3.3mM was found for thereversal of 32P-casein
phosphorylation
by
the C subunit in animportant
early
study
(Shizuta
etal., 1975).
This raises theinteresting
possibility
thatdephosphorylation
of standardphosphorylated substrates,
unlikeautodephosphorylation,
hasaMgADP-dependent
step(possibly
aternary
complex
betweenC,
P-casein and
MgADP),
with a lowaffinity
forMgADP.
Analternative
explanation
is that thehigh
32P-casein concentration used(four
ordersofmagnitude higher
than thatof32P-RIIinthepresent
study)
may have led to the accumulation ofenough
[y-32P]ATP
togive
significant
rephosphorylation
of substrateat moderate concentrations of ADP. A very recent
study
ofC-subunit-catalysed
dephosphorylation
ofP-kemptide
reported
apparentKmvalues for
MgADP
in the,uM
range(Quamar
etal.,
1992).
The
autodephosphorylated
RII could berephosphorylated
with retained
mobility
onSDS/PAGE,
and retained its[3H]cAMP-binding
characteristics,
indicating
that the loss of 32P from theautophosphorylation
site was not causedby
orac-companied
by
anydegradation
ordenaturation ofRII.Effect of cAMP and
kemptide
on theautophosphorylation
and
autodephosphorylation
of cA-PKII
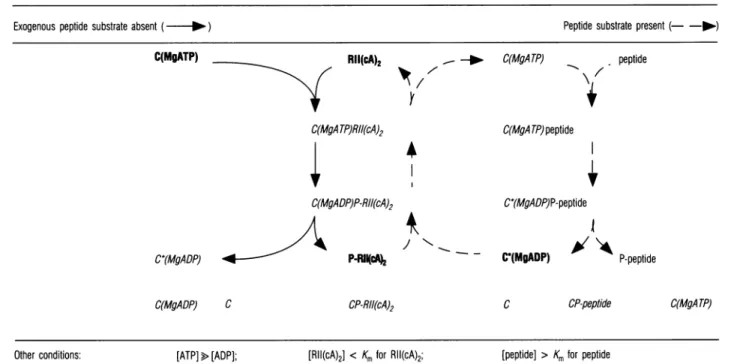
Table 2 Scheme explaining preferential dephosphorylation of P-RII In thepresence of kiasepeptide substrate
The scheme shows the possible complexes involving C subunit, MgADPor MgATP, RII(cA)2and P-RII(cA)2in the absence (left;solid arrows) and the presence (right; broken arrows)of exogenouspeptide substrate [which, unlike RII(cA)2, is presentat aconcentration well above its Km value for phosphorylation]. In the absence of exogenous substrate RII(cA)2is readily
phosphorylated. In the presence of substrate, dephosphorylation of P-RII(cA)2 prevails relative tophosphorylation of RII(cA)2, because: (1) peptide substrate occupies C(MgATP), which is thereby made unavailable forphosphorylation of RII(cA)2; (2) peptide phosphorylation generates C(MgADP), which catalyses dephosphorylation ofP-RII(cA)2. The concentration ofATP is taken to be high, and well above that ofADP (as in the intactcell). Compounds of low abundance are shown in italics and others in bold. Furtherexplanations aregiven in thetext.
Exogenous peptide substrate absent ( - )
C(MgATP) Rll(cA)2
7
C(MgATP)RII(cA)2
_--
C(MgATP)Peptide substrate present (- -l-) peptide C(MgA TP)peptide
4
C(MgADP)P-RII(cA)2 C*(MgADP)P-RIIc42
C(MgADP) C CP-RII(cA)24
- -C*(MgADP)P-peptideI
C*(MgADP)
A~\k
P-peptideC CP-peptide
[RII(cA)2] < Km forRII(cA)2; [peptide] >Km forpeptide
1.0 E 0.5 0. 0 0.25 0 U-0.125 0 1 2
Timeofincubation(min)
Figure 3 cAMP-dependenceofthecA-PKIIautodephosphorylationreaction The timecourseof32P-RII dephosphorylationwasstudied under the conditions described in
thelegendtoFigure1 (10mMMg2+,2 mMADP and 60 nM C subunitwerepresentin all
incubations).,Thedephosphorylationwasstrongly dependentontheconcentrationofcAMP in the medium,whichwas0(0),0.1,M (/\),1,uM ([1),10,uM (@),or0.1 mM(A).
the presence of added MgADP was enhanced several-fold by cAMP(Figure3). The simplest explanation is that only free32p_
RII(cAMP)2 (not 32P-RII in the cA-PKII holoenzyme) was proneto significant dephosphorylation. This was supported by the closely corresponding cAMP-saturation curves (Figure 4) basedondatacalculated from theconcentration-dependence of
the cAMP effect on autodephosphorylation and actual
[3H]cAMP-binding
data (determined in parallel assays). In another set ofexperiments isolated cA-PKII holoenzyme was incubated withMg[y-32P]ATP
in the absence and presence of0 1.00 a,; D .0 X 0.75 a, .E
~0
m 0.50 cc ,0.25 in -0 0 .r-m cu LL 0 10-7 10-6 i-Concn. of cAMP(M) 10-4 0) cm E-'na C) 0 0 cu"Figure4 Correlation between cAMP binding and cAMP promotion of autodephosphorylation
The rate constantfor thepseudo-first-order autodephosphorylationreactionwascalculatedfrom data(obtainedwith different cAMPconcentrations) asshown in Figure 3,and thefractional
occupancyof theputativecAMPbinding site(s) responsiblefor the cAMP effectwascalculated
(see the Experimental section for details) and plotted (0) as a function of the cAMP concentrationduring autodephosphorylation.TheFigurealsoshows datafor[3H]cAMPbinding toRll (0)under similarconditions(presence of MgADP,60 nM Csubunit,37OC)to those usedfortheautodephosphorylation experiment.Note the close correlation betweenthe increase inthe twoparametersas afunctionof risingconcentrations(abscissa) ofcAMP(labelledor
unlabelled). Other conditions: [ATP]>[ADP];
C(MgA TP)
0
. ~ ~~~~
+kemptide No kemptide
NocAMP+cAMP NocAMP +cAMP 20s 60s 20s 60s 20s 60s 20s 60s 67k
-44k
-2 3 4 5 6 7 8
Figure 5 Kemptide blocks the intermolecular, butnotthelntra-holoenzyme, autophosphorylation of cA-PKII
cA-PKIIholoenzyme(0.1 uMwithrespect tosubunitconcentration)wasincubatedat0 °C for
20s(lanes 1, 3, 5, 7)or60s(lanes 2, 4, 6, 8)inthebufterdescribed in thelegendtoFigure 1,but with 2.5 mM Mg2+and4,uM [y-32P]ATP.Insomeincubations (lanes 3, 4 and 7, 8)
cAMP(30 ,uM)waspresent. Inothers(1-4) 300 ,uMof the kinase substratekemptidewas present. Theautoradiographic intensity of the RII bands show that kemptide canblock the
intermolecularphosphorylation of RII (lanes 3, 4), butnotthe intramolecularphosphorylation occurringin the absenceofdissociatingcAMP(lanes 1, 2). The incubation volumewas100,ul,
and 20#l samples were removed for SDS/PAGE and autoradiography as detailed in the
Experimental section.The arrowheads at the left showthe positions (determinedfromthe protein staining pattern)ofstandardproteins (BSAandovalbumin respectively)ofMr67000 (67 k)and 44000(44 k).
cAMP. In either case a rapid phosphorylation of RII was
observed in the absence of kemptide. Kemptide selectively prevented phosphorylation of RII in the presence of cAMP
(Figure 5), i.e. when RII was dissociated from the cA-PKII
holoenzyme. It appeared therefore that kinase substrates suchas
kemptide counteract the accumulation of autophosphorylated RIIby both inhibitingphosphorylation (Figure 5; Table 2) and promotingdephosphorylation (Figure 1; Table 2).
DISCUSSION
Coupling
ofautodephosphorylation
of P-RII andphosphorylation
of standard
peptide
substrate:Implications
for the kineticmechanism
of theprotein kinase reaction
Thepresentstudy has shown that autodephosphorylation of 32p_ RII(cAMP)2 canbe coupled to phosphorylation ofa standard
protein kinase substrate with theproduction of [y-32P]ATP. This implies that MgADP producedinsituonthe C subunit of cA-PK canacceptdirectly the 32p from the autophosphorylated siteon
RII, i.e. that the kinase reactioncanproceed from theternary catalytic complex [C(MgATP)peptide] accordingto:
C(MgATP)peptide-+C*(MgADP)P-peptide
C*(MgADP)+P-peptide (2)
Furthermore, the intermediary C*(MgADP) must be able to reactwithP-RII accordingto:
C*(MgADP)+P-RII(cAMP)2--
C*(MgADP)P-RII(cAMP2-+C(MgATP)+RII(cAMP)2 (3)
[that only one reaction direction is shown does not imply
irreversibility; C*(MgADP) denotes C subunit with bound
MgADP formed in situ through transfer of phosphate from
MgATPto peptide substrate].
Several detailed studies (containing references to important earlierstudies on thesubject)deal with the kinetic mechanism of
the kinase reactionofcatalysed by cA-PK (Whitehouse et al., 1983; Kong and Cook, 1988; Quamar et al., 1992) and the multifunctional Ca2+/calmodulin-dependent protein kinase II
(Kwiatkowski et al., 1990; Katoh and Fujisawa, 1991a). Data have beenpresented supporting sequential binding of MgATP
and peptide, followed by ordered release of P-peptide and MgADP(Whitehouse etal., 1983;Kwiatkowskietal., 1990)as
well as random binding (Kong and Cook, 1988; Katoh and
Fujisawa, 1991a; Quamar et al., 1992). The present study probablyprovides the mostexplicit evidencesofar for releaseof
phosphopeptide before MgADP. Furthermore, the C*(MgADP) complexsoformedis able tobind andcatalysedephosphorylation ofP-RII(cAMP)2.This means that the kinase reaction can also be
reversed in anorderedmanner,with P-RII(cAMP)2 joiningthe
preformed C*(MgADP).
Inapparentcontradictiontothis,acarefulisotope-partitioning study failedtodetectphosphotransfer from phosphokemptideto
C(MgADP), indicating that ADP dissociated faster than
P-kemptide from the complex C(MgADP)P-peptide (Kong and Cook, 1988). Thisdiscrepancycanberesolvedby assumingthat
C*(MgADP) differs from C(MgADP) by promoting more
efficiently thereversal of the kinasereaction. This idea receives
somesupportfromthe presentstudy (Table 1): in the presence
of MgATP and kemptide [when C*(MgADP) is formed] the
autodephosphorylationwasslightly fasterthan inthepresenceof
MgADP [when C(MgADP) dominates]. Unfortunately, the prevalence of C*(MgADP) relativetoothercomplexesof the C subunit(Table 2)is unknown. If 10
%
of totalC subunit is in theC*(MgADP) state in the presence of MgATP and kemptide, C*(MgADP)isabout 12-foldmoreefficient thanC(MgADP)in
catalysing P-RII(cAMP)2 dephosphorylation. If
900%
is in theC*(MgADP) state, the difference in catalytic activity must be slight.
Whereas the
autophosphorylation
ofRII canproceedasbothanintramolecular andanintermolecularreaction(Rangel-Aldao and Rosen, 1976; Figure 5), autodephosphorylation was
detectedonly forP-RIIcomplexed with cAMP (Figures3and4), implyingthat it occurredonlyas anintermolecularreaction.
Possible
physiological
significance
of
autodephosphorylation of
Rul
In standard low-salt buffer,
500%
dephosphorylation of 32p_ RII(cAMP)2 was achieved in about 30 s by 90 nM C subunit (Figure 2). The concentration ofC subunit (0.78,umol/kg) in brain(Hofmannet al., 1977) translates to about 1.6,sM in the cell waterphase, assumingthat the latter occupies 50%
of thebrain volume. Extrapolation would indicate an
auto-dephosphorylation rate of 0.4
s-Q.
This compares favourablywith the value (0.45
s-1)
recently estimated from data onphosphataseaction onautophosphorylatedP-RII inDrosophila head extract(Aszodi etal., 1991).
We have shown that the rate of the C-subunit-mediated phosphorylationof a substrate in the intact cell was very close to thatcalculated fromexperimentswith isolatedenzymesusing a buffer of near physiological ionic strength (D0skeland et al., 1992). Using that same buffer we found the rate of
auto-dephosphorylation of P-RII(cAMP)2 to beabout 8-fold slower (resultsnotshown)than inthe standard low-salt buffer(Figure 2),suggestingthat underphysiologically relevant conditionsthe maximal rate of autodephosphorylation may be somewhat
slower than the maximal rate ofphosphoprotein phosphatase dephosphorylation of P-RII(cAMP)2. It should be stressed that, in the absence ofasuitablequantitative experimental approach
to study therateof dephosphorylation in intact cells, estimates of such rates must be considered hypothetical. A particular experimental problem applies to P-RII(cAMP)2, because enhanceddephosphorylation will leadtoenhancedre-association with the C subunit, formation of holoenzyme, and
re-autophosphorylation by an intramolecular mechanism
(Rangel-Aldao and Rosen, 1976; Figure 5). Therefore the overall phosphorylation of RII may not decrease even if
dephosphorylation is activated.
Buxbaum and Dudai (1989) have suggested that, owing to
autophosphorylation, cA-PKII can remain partially activated
formorethan 10min after cessation ofacAMPsignal,implying
long-term activation of cA-PKII as relevant for learning and memory; seealso Schwartz and Greenberg (1987) forareview of
thistopic. Thepresentdatasuggestthat, ifthe kinase is actively phosphorylating substrates giving risetoC*(MgADP), thebulk ofautophosphorylated P-RIImaybedephosphorylated in1min
evenin the absence ofphosphoprotein phosphatase activity.
Theoretical considerations
The equilibrium constants (in the forward direction) for the phosphorylation of casein by cA-PK(Shizutaetal., 1975) and of myosin light chain by myosin light-chain kinase (Geuss et al., 1985) have been reported tobe 24 and 60-70respectively. This
meansthat itrequiresoneto twoordersofmagnitudemoreADP
than of ATP to achieveequal rates of dephosphorylation and phosphorylation. Since cell ADP is one to two orders of magnitude lower than ATP,onewillexpectthe forwardreaction todominatecompletely in the intact cell. This consideration has led investigators to ignore the reversal of the kinase reaction underphysiological conditions.
The novel observation of the present study is that de-phosphorylation ofautophosphorylated P-RIIcanbe linked to phosphorylation ofa peptide substrate in sucha way that the
relative concentrations of bulk free cellularMgADP and MgATP become lessimportant (Table 2). In fact,evenwhen ATPwasin
extreme excess over ADP, the peptide substrate kemptide
en-hanced the dephosphorylation of P-RII(cAMP)2 (Figure 1). Kemptide also efficiently blocked the phosphorylation of RII (Figure 5), thus shifting the equilibrium between RII and P-RII in favour ofdephosphorylation. The blocking of phosphorylation ofRII(cAMP)2 by kemptide is easily explained bythelatter out-competing the former for complex formation with C(MgATP) (illustrated in Table 2), since kemptide was present at
concen-trations well above its apparent Km value of about 5,uM (Whitehouse et al., 1983). The enhanced dephosphorylation in the presenceofkemptide is explained (1) by the production of
C*(MgADP) from C(MgATP) during kemptidephosphorylation and (2) by the fact that P-RII competes successfully with P-kemptide for C*(MgADP). This ispossible because P-kemptide bindsto C withabout 1000times loweraffinity thankemptide, theapparentKdofP-kemptide bindingtoCbeing about2-4mM (Whitehouseetal., 1983; Quamaretal., 1992). In contrast, the C subunit binds P-RII(cAMP)2 with only moderately lower affinity than it binds
RII(cAMP)2
(Granotetal., 1980;D0skelandetal., 1993). Thismeansthatattheconcentrationsofkemptide
used in thepresentstudy (andpresumablyattheconcentrations of most exogenous substrates encountered in the intact cell)
phosphorylated substrates do not compete efficiently with P-RII(cAMP)2 for binding toC*(MgADP).
The mechanism described in the present study may be
im-portant for other kinases releasing phosphorylated product before MgADP. One possible candidate reaction is auto-dephosphorylation of the Ca2+/calmodulin-dependent multi-functional protein kinase II. A sequential release of phosphoproduct and MgADP has been suggested for thisenzyme
(Kwiatkowskietal., 1990), but isnotgenerally accepted (Hanson and Schulman, 1992). It is of interest that the presence of exogenoussubstratewasshowntodecrease therateaswellasthe
steady-state level ofautophosphorylation of thisenzyme(Katoh
andFujisawa, 1991b).
This workwas supported by the Medical Research Council of Norway (NAVF) and the Committee of the Nordic Insulin Foundation.
REFERENCES
Asz6di, A., MUller, U., Friedrich,P.and Spatz, H.-C. (1991) Proc. Natl. Acad. Sci. U.S.A. 88,5832-5836
Beebe, S.J. andCorbin,J. G. (1986) Enzymes3rd Ed. 17A,43-111
Bollen, M. andStalmans,W. (1992)Crit. Rev. Biochem. Mol. Biol. 27,227-281
Bradford, M. M.(1976)Anal. Biochem.72,248-254
Buxbaum, J. D.and Dudai,Y.(1989)J. Biol. Chem.264,9344-9351
Cohen,P. (1989)Annu.Rev. Biochem.58,453-508
Doskeland,A.P., Schworer,C.M., Doskeland, S. O., Chrisman, T. D., Soderling,T.R., Corbin,J.D.and Flatmark, T. (1984)Eur. J.Biochem. 145,31-37
D0skeland,A.P., Vintermyr,O., Flatmark, T., Cotton,R. G. H. and Doskeland, S.0. (1992)
Eur. J. Biochem.206,161-170
D0skeland,S.O., Vintermyr,0.K., Corbin,J.D.and0greid,D.(1987)J. Biol.Chem.262,
3534-3540
Doskeland, S.O., Maronde, E. andGjertsen,B.T. (1993)Biochim. Biophys. Acta,inthe press
Edelman,A.M., Blumenthal, D.K.and Krebs,E.G. (1987)Annu. Rev. Biochem.56,
567-613
Ekanger, R.,Vintermyr,0.K., Houge,G., Sand, T.-E., Scott,J.D., Krebs, E.G., Eikhom, T., Christoffersen, T., 0greid,D. and D0skeland,S.0.(1989)J. Biol. Chem.264,
4374-4382
Geuss, U., Mayr,G. W. andHeilmeyer, M. G.(1985) Eur. J. Biochem. 153,327-334
Granot, J., Mildvan,A.S., Hiyama, K., Kondo, H.and Kaiser,E. T. (1980)J.Biol. Chem.
255,4569-4573
Hanson,P.I.andSchulman, H.(1992)Annu. Rev. Biochem.61,559-601 Hidaka,H.andKobayashi, R.(1992)Annu. Rev.Pharmacol. Toxicol.32,377-397
Hofmann, F., Bechtel, P.J. and Krebs, E.G.(1977)J. Biol. Chem.252,1441-1447
Katoh, T.andFujisawa, H. (1991a)Biochim. Biophiys.Acta1091,205-212
Katoh,T. andFujisawa, H. (1991b)J. Biol. Chem.266,3039-3044
Kong, C.-T.andCook, P. F. (1988) Biochemistry 27,4795-4799
Kwiatkowski, A.P.,Huang,C. Y. andKing, M.M. (1990) Biochemistry 29,153-159
Laemmli, U. K.(1970) Nature(London) 227, 680-685
McKnight,G.S. (1991)Curr.Opin.Cell Biol.3,213-217
Ogreid,D.and D0skeland,S.0.(1980) FEBSLett.121,340-344 Quamar, R., Yoon, M.-Y. andCook,P. F.(1992) Biochemistry 31,9986-9992
Rangel-Aldao,R.andRosen,0.M. (1976)J. Biol. Chem.251,7526-7529 Rosen,0.M. and Erlichmann,J.(1975)J. Biol.Chem.250,7788-7794 Schwartz,J. H. andGreenberg, S.M.(1987)Annu.Rev. Neurosci. 10,459-476
Scott,C. W. and Mumby,M. C.(1989) Mol.Endocrinol. 3,1815-1822
Scott,J.D.,Glaccum,M.B., Fischer, E.M.and Krebs,E.G.(1986) Proc. Natl. Acad.Sci. U.S.A.83,1613-1616
Shizuta, S.,Beavo, J.A., Bechtel,P.J., Hofmann,F.andKrebs, E. G. (1975)J.Biol. Chem.
250,6891-6896
Sugden,P.H., Holladay, L.A., Reimann, E.R.andCorbin,J.D.(1976) Biochem.J.159,
409-422
Taylor, S., Buechler,J. A. andYonemoto,W. (1990)Annu. Rev. Biochem.59,971-1005
Whitehouse, S., Feramisco,J.R., Casnellie,J.E., Krebs, E. G. andWalsh, D.A.(1983)
J. Biol. Chem.258,3693-3701