Review
Evaluating
‘
Plasticity-First
’
Evolution
in
Nature:
Key
Criteria
and
Empirical
Approaches
Nicholas
A.
Levis
1,*
and
David
W.
Pfennig
1Many biologists are asking whether environmentally initiated phenotypic
change(i.e.,‘phenotypicplasticity’)precedes,andevenfacilitates,evolutionary
adaptation. However, this ‘plasticity-first’ hypothesis remains controversial,
primarilybecausecomprehensivetestsfromnaturalpopulationsaregenerally
lacking.We brieflydescribe theplasticity-firsthypothesis andpresent
much-neededkeycriteriatoallowtestsindiverse,naturalsystems.Furthermore,we
offeraframeworkforhowthesecriteriacanbeevaluatedanddiscussexamples
wheretheplasticity-firsthypothesis has been investigated in natural
popula-tions.Ourgoalistoprovideameansbywhichtheroleofplasticityinadaptive
evolutioncanbe assessed.
NeedforaPredictiveFramework
Amongtheenduringproblemsofevolutionarybiologyisexplaininghowcomplex,adaptivetraits originate[1,2].Althoughitiswidelyassumedthatnewtraitsarisesolelyfromgeneticfactors[3], manyresearchersareaskingwhetherenvironmentallyinitiatedphenotypicchange–inother wordsphenotypicplasticity(seeGlossary)–precedesandfacilitatesadaptation[4–12].
Thisalternativeroute,dubbedtheplasticity-firsthypothesis[4,13],restsontheobservation
thatphenotypicplasticityoftenproducesdevelopmentalvariantsthatcanenhancefitnessunder
stressfulconditions[4,5,14].Ifunderlyinggeneticvariationexistsinthetendencyormannerin whichindividualsproducesuchvariants(asisoftenthecase[15]),thenselectioncanrefinethe
traitfromaninitial,potentiallysuboptimalversionthroughquantitativegeneticchangesovertime; inotherwordsgeneticaccommodationoccurs[4].Furthermore,dependingonwhetheror notplasticityisfavored[16],thisselectioncanrespectivelypromoteeitherincreased environ-mental sensitivity –which maintains the trait as a polyphenism(sensu [17]) ordecreased environmentalsensitivity–whichcanresultintheconstitutiveexpressionofthetrait;inother wordsgeneticassimilation(sensu[18]).By‘jump-starting’phenotypicchangeinanadaptive direction[19],environmentallyinducedphenotypicchangeprecedes,andpromotes,the evo-lutionaryoriginsofacomplex,adaptivetrait(Figure1;Box1).
Wheninitiatedbyplasticity,refinementofadevelopmentalvariantintoanadaptivetrait(whether
novelornot)doesnotrequirenewgenes.Instead,environmentallyinducedphenotypicchange setsinmotionanevolutionarysequenceinwhichselectionpromotesadaptationbyactingon existinggeneticvariation(e.g.,[15,20–22]).Inessence,suchselectionrefinesatraitthrough
evolutionaryadjustmentsinboththeformandregulationoftraitexpression.Theoutcomeofthis processisanadaptivephenotypethat,relativetoitsinitialstate,hasbeenmodifiedbothinits
Trends
Phenotypic plasticity has long been
proposed to precede and possibly
facilitateadaptiveevolution.
This‘plasticity-firsthypothesis’ is
con-troversialbecauseskepticsarguethat
itlackscompellingevidencefrom
nat-uralpopulations.
A chief difficulty with demonstrating
plasticity-firstevolutioninnatural
popu-lationsisthat,onceatraithasevolved,
itsevolutioncannotbestudiedinsitu.
Togetaroundthisdifficulty,
research-erscanstudyextantlineagesthatact
asancestral-proxiestothelineage
pos-sessingthefocaltrait.
Usingsuchanapproach,keycriteriaof
the plasticity-first hypothesis can be
evaluated using a relatively simple
experimentaldesign.
Applyingthesecriteriatovarious
sys-tems,theplasticity-firsthypothesishas
some empirical support. However,
morestudiesareneededto
conclu-sivelydeterminethe roleof plasticity
inadaptiveevolution.
1DepartmentofBiology,CampusBox
3280,UniversityofNorthCarolina, ChapelHill,NC27599,USA
*Correspondence:
morphologicalandphysiologicalattributesaswellasinitsenvironmentalsensitivity.Ofcourse, other evolutionary forces (e.g., genetic drift, mutation) could alter the degree of plasticity. However,theplasticity-firsthypothesisassumes thatanysuchchangeoccurredvia genetic
accommodation,which(bydefinition)isdrivenbyselection.
Althoughlabstudieshavedemonstratedthattheplasticity-firsthypothesiscanpromote
adap-tation(e.g.,[18,23–25]),andtherearesuggestivefieldstudies(e.g.,[26–30],reviewedin[31]), whetherplasticity,followedbygeneticaccommodation,hasactuallycontributedtotheevolution ofanycomplextraitinanynaturalpopulationiscontroversial[32–35].Partofthedifficultyisthat
thekeycriteriaoftheplasticity-firsthypothesishavenotbeenmadeclear.However,ifasstatedin
tworecentprominentreviews:‘whatremainstobedoneistogeneratecreativeapproachesto collectingempiricaldatafromnaturalpopulationstotestpredictions...’[11],andif‘thebestway toelevatetheprominenceofgenuinelyinterestingphenomenasuchasphenotypicplasticity...is tostrengthentheevidencefortheirimportance’[35],thenthesecriteriaandpredictionsmustbe madeclearandrigorouslytested.
Wedescribeherekeycriteriafortestingtheplasticity-firsthypothesis.Wealsopresentageneral
frameworkinwhichthesecriteriacouldbeevaluatedinnaturalpopulations,andwediscusscase
Glossary
Crypticgeneticvariation:genetic
variationthatnormallyhaslittleorno
effectonphenotypicvariationexcept
underatypicalconditions.
Geneticaccommodation:a
mechanismofevolutionwhereina
phenotype,generatedbyeithera
mutationorenvironmentalchange,is
refinedintoanadaptivephenotype
throughselectiondrivingquantitative
geneticchanges.Accommodation
canalsopromoteeitherincreasedor
decreasedenvironmentalsensitivityof
thefocalphenotype;when
environmentallyinducedphenotypes
loseenvironmentalsensitivity,they
undergo‘geneticassimilation’.
Geneticassimilation:anextreme
formof‘geneticaccommodation’ that
occurswhenselectioncauses
environmentallyinduced(i.e.,plastic)
phenotypestolosetheir
environmentalsensitivityover
evolutionarytime.
Noveltrait:broadly,anymajor
developmentalinnovation;sometimes
definedasabodypartthatisneither
homologoustoanybodypartinthe
ancestrallineagenorserially
homologoustoanyotherbodypart
ofthesameorganism;adifficult
concepttodefine.
Phenotypicaccommodation:the
maintenanceofanovel,inducedtrait
orphenotypethatisanautomatic
consequenceofmultidimensional
adaptivephysiological,morphological,
and/orbehavioralplasticityintheface
ofadevelopmentalchange.
Phenotypicplasticity:theabilityof
anorganismtoalteritsbehavior,
morphology,and/orphysiologyin
responsetochangesin
environmentalconditions;sometimes
usedsynonymouslywith
developmentalplasticity.
Plasticity-firsthypothesis:a
mechanismofadaptiveevolutionin
whichenvironmentalperturbation
leads,viaphenotypicplasticity,to
developmentalreorganization(via,e.
g.,alteredgeneexpression)and
uncovers‘crypticgeneticvariation’
for,andultimatelyproductionof,a
noveldevelopmentalvariant(i.e.,trait)
thatimmediatelyundergoes
‘phenotypicaccommodation’andis
subsequentlyrefinedthrough‘genetic
accommodation’ somedefinitions
includecasesinwhichmutation
initiatestraitorigin(notdiscussed
here;seeBox1).
(A)
(B) (C)
(E) (D)
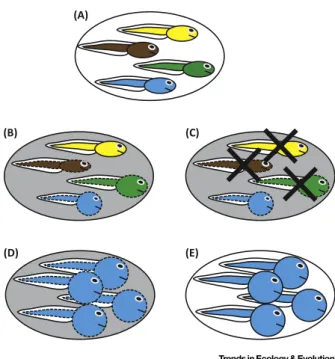
Figure1.AnIdealizedRepresentationofHowPlasticity-FirstEvolutionLeadstoaNovel,Adaptive,Complex Trait.(A)Ageneticallyvariablepopulation(here,different-coloredtadpolesrepresentdifferentgenotypes)(B)experiencesa
novelenvironment(shading),whichimmediatelyinducesnoveldevelopmentalvariants(dashedlines).However,different
genotypesvaryinthemannerinwhichtheyrespond(orindeed,iftheyrespondatall).(C)Selectionactsonthisformerly
crypticgeneticvariation(revealedbythechangeinenvironment)bydisfavoringthosegenotypesthatproducephenotypes
thatarepoorlyadaptedinthenewenvironment(here,theround-bodiedphenotypeisfavored,whereasallothersare
disfavored,asindicatedbythe‘X’).(D)Thisselectionalsoleadstotheadaptiverefinementofthefavoredphenotype
(depictedherebytheenlargementofthebluetadpole).Iftheancestralphenotype(i.e.,narrow-bodiedtadpole)isstill
maintainedintheancestralenvironment(seeA),thentheresultisanovelpolyphenism.(E)However,selectionmightinstead
favorthelossofplasticity(i.e.,geneticassimilation),resultinginanovelphenotypethatisnowproducedconstitutively,even
whenthepopulationexperiencestheancestralenvironment(indicatedherebythelossofshadinganddashedlines).Note
thatobservationsfromnaturalsystemslikelywillnotbeasclear-cutastheprocessdescribedhere.Furthermore,although
wehaveshownhowaplasticity-firstprocesspromotesnovelty,thisprocesscouldalsoexplaintheevolutionoftraitsthatare
Polyphenism:environmentally
inducedalternativephenotypes.
Reactionnorm:agraphical
representationofthesetof
phenotypesthatasinglegenotype
producesinresponsetosome
specificenvironmentalvariable(s);
individualsshowplasticityiftheir
reactionnormisnon-horizontal.
studiesthathaveutilizedthisframeworktofindsupportforthesecriteria.Ourgoalistoprovidea
roadmapfor testingthe plasticity-first hypothesisandtherebyclarify the roleofplasticity in
adaptiveevolution.
KeyCriteriaforthePlasticity-FirstHypothesisof AdaptiveEvolution
Beforeoutliningcriteriafordemonstratingplasticity-firstevolutioninnaturalpopulations,wenote
thatthemostcompellingevidenceforthisprocesswouldbetoactuallyobserveittakingplace. Indeed,plasticity-firstevolutioncouldpotentiallybeobservedinreal-timein:(i)casesinwhich
naturallyoccurringpopulationsexperiencerapidenvironmentalchange(e.g.,climatechangeor introduced species[26]), or(ii) resurrectionstudies;for example,usingseeds from datable sedimentasancestralcontraststomodern,derivedindividualsfromthesamepopulation.In bothcontexts,theancestralconditionwouldbeknown,andenvironmentalchangecanoccur swiftlyenoughtoobservean evolutionaryresponse.Notethatathird context,studyingthe plasticity-firsthypothesisinlabpopulations of rapidlyevolving organisms [36,37],wouldbe
worthwhilebutwouldnotclarifywhetherplasticityhascontributedtoadaptationinanynatural population[32–35].
Formostsystems,however,itislikelythatonlythefinalproductsofanyputativeplasticity-first
processwillbepresentinmodern-daypopulations.Insuchsituationsthechiefdifficultywith
demonstratingplasticity-firstevolutionisthat,onceatraithasevolved,itsevolutioncannotbe
studiedinsitu.Togetaroundthisdifficultyonecouldeitherstudylineages(i.e.,speciesand/or
populations)thatareancestral-proxiestothelineagepossessingthefocaltrait[5]or, alterna-tively,evaluatewhetherenvironmentallyinduceddifferenceswithintaxareflectadaptive(fixed)
differencesamongthesesame(orrelated)taxa[31].Oftheseapproaches,ancestral-derived comparisonsproducestrongerevidenceofgeneticaccommodation[31].Althoughsuch com-parisonsareonlyfeasibleinsystemswithwell-understoodnaturalhistoriesandreadily acces-sibleancestral-proxyandderivedlineages(Box2),wefocusonthisapproachinsuggestingfour keycriteriaoftheplasticity-firsthypothesis.
Importantly,validationofanyonecriterion,byitself,isinsufficienttoestablishthatplasticity-first
evolutionhasoccurred.Forinstance,manysystemsappeartosatisfycriterion1below–thata trait is present in ancestral-proxy lineages as an environmentally induced variant [13,31]. However, themere existenceof suchancestralplasticity isinsufficient to demonstrate that
thetraitevolvedviaaplasticity-firstprocess.Asnotedabove,severalevolutionarymechanisms Box 1. Evolutionary Potential of Environmentally Initiated Versus Mutationally Initiated Phenotypic Change
Stresses– suchasmutationorenvironmentalchange– cangeneratenoveldevelopmentalvariants[4,5].The
sub-sequentgeneticaccommodationofsuchvariantscanoccurwhethertheyareinducedbymutationorachangeinthe
environment[4],andthesetwomodesofinductionareofteninterchangeable[4,12,13].However,environmentally
triggerednoveltieslikelyhavefargreaterevolutionarypotentialthanmutationallyinducedones,foratleastthreereasons
[4].First,changesintheenvironmentoftenaffectmanyindividualssimultaneously,incontrasttoageneticmutation,
whichinitiallyaffectsonlyoneindividualanditsimmediatedescendants(also,thevastmajorityof mutationsare
deleterious[58,59]).Thiswidespreadimpactofenvironmentalchangeenablesanewlyinducedtraittobetestedamong
diversegenotypes, thereby providingfertilegroundforselectiontoactandincreasingthechances thatgenetic
accommodationwilloccur[4].Second,althoughthechancethataparticularmutationwilloccurisnotinfluenced
bywhetherornottheorganismisinanenvironmentinwhichthatmutationwillbeadvantageous–inotherwords,
adaptivelydirectedmutationdoesnotoccur[60]– thesituationisdifferentforenvironmentallytriggeredtraits.Suchtraits
arealwaysassociatedwithaparticularenvironment– theonethattriggeredit.Therefore,environmentallyinducedtraits
aremorelikelythanmutationallyinducednoveltiestoexperienceconsistentselectionanddirectionalmodification[4].This
allowsnewenvironmentstoimmediatelyproduceandselectamongnewphenotypesandrapidlyrefinetheirexpression
[5].Third,plasticitypromotesthestorageandreleaseofcrypticgeneticvariation–inotherwordsvariationthatis
expressedonlyunderatypicalconditions(e.g.,[41,45,46]).Thereleaseofsuchvariationultimatelymakesgenetic
accommodationpossible[9,44].Forthesereasons,environmentalinitiationmighthavegreaterevolutionarypotential
canleadtothesubsequentchangeinthedegreeofplasticity,buttheplasticity-firsthypothesis
requiresthatselectionfavoredanysuchchanges.Thus,confirmationofseveral(ideally,allfour)
criteriaincreasessupportforthismodeofevolution.
Criteria1and2focusondetectingpre-existingplasticityanddevelopmentalreorganization (i.e., phenotypic accommodation), whereas criteria 3 and 4 focus on the subsequent refinement ofexpression and formof the focal phenotype (i.e., genetic accommodation).
Notethatwehaveomittedcriteriaspecificallydealingwithhowdevelopmentalreorganization
canproduceanovelphenotypeinancestral-proxylineagesbecausesuchreorganizationhas extensivesupport(reviewedin[4])andiswidelyaccepted.However,weexpectanychanges inthemeasuredtraitbetweenancestral-proxyandderivedlineagestobeaccompaniedby concurrentmechanisticchanges(e.g.,hormones,alternativesplicing,transcription factors, cis-regulatory elements, etc.) [4,9,38–40] underlying production and/or regulation of that trait.
Belowwelisteachcriterionfollowedbytherationalebehindit.Althougheachofthesecriteriahas beendiscussedpreviously(e.g.,[4,5,9,22,29]),theyhavenotbeencollectivelyassembled,and clearmethodsfortestingthemhavenotbeenprovidedbeforenow.
Criterion1.TheFocalTraitWillBeEnvironmentallyInducedinAncestral-ProxyLineages
Themostfundamentalcriterionoftheplasticity-firsthypothesisisthatthetraitofinterestshould
exhibitancestralplasticity.By‘ancestralplasticity’wemeanthatadevelopmentalvariant(or character state), similar to the derived (possibly fixed) trait, should be expressed among
individualsfromancestral-proxylineageswhentheseindividualsexperiencethederived envi-ronment; in other words the environmental conditions in which the focal trait is normally expressedinderivedlineages[4].Inaddition,ifthereisphylogeneticsupportthatataxonwith aninducibletraitresemblestheancestralcondition,and/orifthedevelopmentalmechanismsfor producingthetraitareconservedinrelatedspecieswherethetraitdoesnotregularlyoccur,then thiswouldsuggestthatthetraitstartedoutasanenvironmentallyinduceddevelopmentalvariant
[4].Itisimportanttonotethatthedevelopmentalvariantneednotbeexpressedtothesame degreeasisseeninderivedlineages.
Box2.ConsiderationsWhenChoosingaStudySystem
Asnoted,onlythefinalproductsofplasticity-firstevolutionmightstillexistformostsystems,makinginsituinvestigation
difficult.Onewaytocircumventthisdifficultyistouseotherlineages(i.e.,speciesorpopulations)asproxiesforthe
ancestralconditionandcomparethemtoderivedlineagesthatpossessthefocaltrait.Todoso,onemustknowthe
phylogeneticrelationshipsbetweenlineagesthatdifferintheformof,orpropensitytoproduce,aninducedphenotype.
Thisisimportantbecauseitisessentialtoknowwhichlineagescanrepresenttheancestralandderivedstates.However,
thismeansthatsomesystemsthatlackphylogeneticinformationmightnotbesuitableforevaluatingtheplasticity-first
hypothesisusingtheapproacheswehaveoutlinedhere.
Inaddition,ourcriteriadependonknowingtheenvironmentalstimulusthatmighthaveinitiallyledtoproductionofthetrait
aswellastheselectivepressuresleadingtoitsrefinementorchangeinfrequencyofexpressioninderivedlineages.
Withoutthisinformation,evaluatingtheplasticity-firsthypothesisisnotpossible.
Whileknowledgeofthephylogeneticrelationshipsandrelevantenvironmentalfactorsisrequired,othernon-essential
characteristicscouldimprovetheutilityofasysteminstudyingtheplasticity-firsthypothesis[61],including:
(i) Multiple,parallelderivedlineageswithvaryingdivergencetimesfromtheancestral-proxylineage.
(ii) Knowledgeoftheecologicalcircumstancesexperiencedby,andtheselectiveagentsimpingingon,both
ancestral-proxyandderivedlineages.
(iii) Aquantifiabletraitthatisreadilyinducedunderlaboratoryconditions.
(iv) Adequategenomicresourcesforinvestigationofmolecularunderpinnings.
(v) Otherfeaturesof‘typical’ modelorganisms(e.g.,fastgenerationtime,easytorearinlargenumbersinthelab,
Criterion2.CrypticGeneticVariationWillBeUncoveredWhenAncestral-ProxyLineages
ExperiencetheDerivedEnvironment
Ifatraitonlyexistsasanenvironmentallyinducedvariant,andthereforeisinfrequently(ornever) exposedtoselection,thengeneticvariationshouldaccumulateintheresponseofthetraittoa novel environment (or in components that make up the trait) that can be revealed when environmentalconditionschange (Figure 1C).In a novelenvironment, thiscrypticgenetic variationwouldbeuncovered–manifestasanincreaseinheritabilityorgreaterphenotypic variation resultingfromperturbation of anevolutionary capacitor[30,41–43]–and actas a selectable substrate (Figure 1D) [4,9,44–46]. Subsequently, however, lineages inthe novel (derived)environmentshouldexperienceaselectivesweepandlosethisvariationasthetrait undergoes genetic accommodation [4,9,47]. Thus, once derived lineages have undergone genetic accommodation, theyshould exhibitreduced heritability and/orgenetic variation in thetraitofinterest.
Criterion3,TheFocalTraitWillExhibitEvidenceofHavingUndergoneanEvolutionary
ChangeinitsRegulation,Form,orBothinDerivedLineages
Thiscriterioncanbemanifestasachangeintheslopeorelevation(orboth)ofthereaction norm inderivedversus ancestral-proxy lineages. Because achange in sloperepresents a changeintheregulationofatraitandachangeinelevationindicatesachangeinitsform[30,48],
finding either in derived lineages would suggest genetic accommodation [4]. Furthermore, findingthatreactionnormsarefixedacrossdifferentenvironmentswouldsuggestthatthetrait
has beengenetically assimilated.Finally, changesin reactionnorms should bemirrored by changesinthemechanismsunderlyingthetrait(e.g.,hormones,alternativesplicing, transcrip-tionfactors,cis-regulatoryelements,etc.)[4,9,38–40].
Criterion4.TheFocalTraitWillExhibitEvidenceofHavingUndergoneAdaptiveRefinement
inDerivedLineages
Undertheplasticity-firsthypothesis,thefrequencythatatraitisexpressedwilldeterminethe
degree to which it is refined by selection [4]. Therefore, compared with individuals from
ancestral-proxylineages,individualsfromderivedlineagesshouldproducesuperiorversions ofthe trait when bothtypes of lineages experience environmentsthat inducethe trait. For example,derivedindividualsshouldproduceaversionofthetraitwithimprovedfunctionality, fewer side effects, or a lower threshold for expression [4]. This criterion is based on two assumptions:(i)individualsinderivedlineagesshouldexpressthe traitmorefrequentlythan individualsinancestral-proxylineages(whichproducethetraitinfrequently),and(ii)atraitina populationinwhichitisexpressed(andexposedtoselection)morefrequentlyshouldevolve greaterandmorerapidrefinement[4].Asacorollary,thefitnessconsequencesofatrait–in otherwordsitscontributiontoindividualfitness–shouldbehigherinderivedlineagesthanin ancestral-proxylineageswhenbothareinthederivedenvironment[4,49].
EvaluatingthePlasticity-FirstHypothesisin Nature:General Framework
Ratherthandescribehoweachcriterioncouldbetestedindividually,wepresentaschemefor testingallcriteriasimultaneously(Figure2).Ourschemeconsistsoftwophases:(i)identification
ofancestral-proxyandderivedlineages,and(ii)common-gardenexperimentationinwhichboth typesoflineagesarerearedunderenvironmentalconditionssimilartothosethattheywould experienceinthewild.
Beforestarting,itisessentialthatbackgroundinformationaboutthestudysystemisknown (Box 2).Assuming this informationis available, the first phaserequires inferring ancestral
becloselyrelatedtothederivedlineages,buttheyshouldnotpossessthederived(potentially
fixed)traitwhenintheirnatural(ancestral)environmentthatlackstheinducingstimulus.
In the second phase of testing the plasticity-firsthypothesis, all key criteriacan be tested
simultaneously.Thisrequiresrearingmultiplesibshipsfromancestral-proxyandderivedlineages (Figure2A)inconditionsrepresentingancestralandderivedenvironments(e.g.,withoutandwith theinducingstimulus,respectively).Thiscouldbedonewithacommon-gardenorreciprocal transplantdesign.Anadvantageofthisapproachisthatitallowsonetoestimatehowmuch evolutionarydistance(fromancestraltoderivedtrait) wascoveredwhenanancestrallineage experiencedanovelenvironment(indeed,suchenvironmentallyinducedvariantstypicallycover onlypartofthisevolutionarydistance;e.g.,[22,27,29,50]).Multiplesibshipsarenecessary,both becausetheexpressionofphenotypicplasticityoftenvariesamonggenotypes[15]andbecause criterion 2 specifically involves comparing genetic variability across ancestral and derived
environments.
Fromsuch an experiment, observations and measurementsthat could validate the criteria include:awidevarietyofphenotypesproducedbyancestral-proxysibshipsinderived environ-ments,someofwhicharealongthesameaxisofvariationasthefocalphenotypepossessedby derived individuals(criteria 1 and2; Figure 2B); an increase inheritability ofthe trait(s) (or componentsofthetrait)amongancestral-proxysibshipswhenrearedinderivedenvironments (A)
Lineage 1 Lineage 2 Lineage 3 Ancestral phenotype
Induced novel phenotype
Canalized novel phenotype
(C) Time 1 Time 2
Compare fitness of induced versus canalized novel phenotypes
(B)
Phenotype
Ancestral environment
Novel environment
Environment (A, ancestral; N, novel) N N
A A A N
Pre-exis!ng plas!city?
Compare reac!on norms and heritabili!es Lineage 1 Lineage 2 Lineage 3
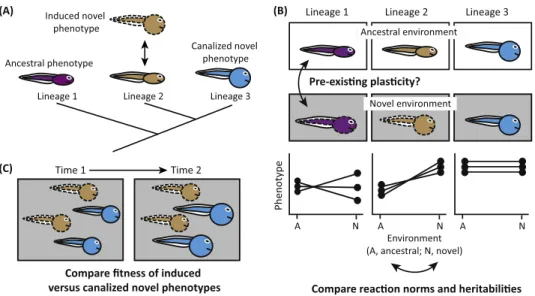
Figure2.ApproachesforTestingtheKeyCriteriaofthePlasticity-FirstHypothesis.(A)Asaninitialstep,identify
lineagestoserveasancestral-proxiestothelineage(s)thatproduce(s)thefocaltrait.Inthisexample,lineage1doesnot
produceanoveltraitduringnormaldevelopment;lineage2producesthenoveltraitaspartofnormaldevelopment
dependingonenvironmentalconditions(i.e.,itexhibitsadaptiveplasticityintraitexpression);lineage3producesthenovel
traitregardlessofenvironmentalconditions(i.e.,itiscanalized,meaningthatitexhibitsnoplasticityinthetrait).(B)Next,use
acommon-gardenapproachtodetermineif:(i)ancestral-proxylineagesthatnormallylackthetraitofinterest(lineage1)
producedevelopmentalvariantsofthetraitthroughphenotypicplasticitywhenexperiencingthenovelenvironmental
stimulus(criterion1);(ii)thenovelenvironmentuncoverscrypticgeneticvariationintheseancestral-proxylineages(criterion
2);(iii)phenotypicvariationrevealedintheancestral-proxylineagesisgreaterthaninderivedlineages(andmightberandom
withrespecttoitsadaptivevalueinthenovelenvironment);and(iv)reactionnormshaveevolvedinamannersuggestingthat
selectionhasrefinedtraitexpressioninthenovelenvironment(indicatedbydirectionalreactionnormsinlineage2versusflat
reactionnormsinlineage3;criterion3).(C)Finally,useacommon-gardenapproachtodetermineifthetraithasindeed
undergoneadaptiverefinementbycomparingthefitnessofindividualsfromlineagesthatproducethenovelphenotype
facultativelyversusconstitutively(criterion4).Ifgeneticaccommodationhasoccurred,thelattershouldoutperformthe
formerinthenovelenvironment[indicatedbyincreasedgrowth(largersize)oftheblue(canalized)phenotypeattime2],but
(criterion2;Figure2B);achangefromreactionnormslackingconsistentdirectionalityamong ancestral-proxysibshipstohavingarelativelyconsistentslopeamongderivedsibships(criteria 2and3;Figure2B);achangeintheslopeorelevationofthereactionnorminderivedsibships relativetoancestral-proxysibships(criterion3;Figure2B);greatervarianceintraitvaluesfor ancestral-proxysibshipscomparedtoderivedsibshipswhenrearedinderivedenvironments (criteria2and3;Figure2B);greaterfitness(e.g.,survival,growth,size,development,fecundity,
etc.)ofderivedsibshipsthanancestral-proxysibshipswhenrearedinderivedenvironments (criterion4;Figure2C);andastrongercorrelationbetweenfitnessandtraitvalueinderived
sibshipsthanancestral-proxysibshipswhenrearedinderivedenvironments(criterion4;see
Box3forothersuggestionsfortestingcriterion4).
EvaluatingthePlasticity-FirstHypothesisin Nature:IllustrativeCaseStudies
Toillustratetheaboveframeworkfortestingthekeycriteriaoftheplasticity-firsthypothesis,we
describetwo casestudies.Althoughthesestudies arenot necessarilythe mostcompelling examplesofplasticity-firstevolution,theyillustratehowtotesttheplasticity-firsthypothesisina
naturalpopulation.
SpadefootToads
Speatadpoles exhibita novel polyphenismnot seenin otherspecies[22] consistingofan omnivore ecomorph, which eats detritus primarily, and a morphologically and behaviorally distinctivecarnivoreecomorph,whichspecializes onshrimpandwhichexpressesasuiteof unique,complextraits[51].Omnivoresarethedefaultmorph;carnivoresareinducedwhena young omnivoreeats shrimp orothertadpoles [28,52]. However, most populations harbor heritablevariationinthepropensitytoproducecarnivores[51].Carnivoresarisedevelopmentally fromanomnivore-likeformviaacceleratedgrowthoffeatures[53],andfrequency-dependent, disruptiveselection–stemmingfromresourcecompetition–maintainsbothecomorphswithin most populations [54]. Several studies, taken together, suggest that this novel carnivore ecomorpharosethroughplasticity-firstevolution[22,41,55].
Onestudyfoundsupportforcriteria1,3,and4 [22].Using ancestralcharacterstate recon-struction, Scaphiopus couchii was chosen as a proxy for non-plastic Spea ancestors
Box3.AlternativeMethodsforEvaluatingCriterion4ofthePlasticity-FirstHypothesis
Ourcriteriamightbevalidatedusingmethodsotherthanthosepresentedabove.Thisisespeciallylikelytobetruefor
criterion4becausewhatconstitutesasuperiortraitissystem-specific.Wehighlightheresomealternativeapproachesfor
testingcriterion4.
(i) Compareindifferentlineagestheamountofstimulusrequiredtoinducethefocaltrait(e.g.,[62]).Forexample,inthe
caseoflight-inducedresponseinplants,onecouldmeasuretheamountoflightneededtoinducethetraitand,inthe
caseofpredator-induced responsesineitherplantsoranimals,onecouldmeasuretheintensityofpredationor
abundanceofpredatorsneededtoinducethetrait.Moreover,iftheunderlyingendocrinesignalsareknown,onecould
manipulateconcentrationsofhormonestodeterminetheamountneededtoinducethetrait.Generally,theamountof
stimulusrequiredtoelicitinductionofatraitshouldbeinverselyproportionaltotheamountofevolutionarytimea
populationhasbeenexposedtothatstimulus.Thus,thethresholdofinductionshouldbelower(orzero)inderived
sibshipsthaninancestral-proxyones.
(ii) Performexperimentsinwhichindividualsfromancestral-proxyandderivedlineagesaredirectlysetagainstone
another(Figure2C)[28].Forexample,ifderivedindividualshaveanovelresource-acquisitiontrait,thentheycouldbe
directlycompetedwithancestral-proxyindividualsinobtainingthatresource.Onecouldthendetermineif(aspredicted)
derivedindividualsobtainmoreresourceandexhibitimprovedsurvivalorgrowththanancestral-proxyindividuals.
(iii) Measureselectiononthetraitinancestral-proxy,derivedpolyphenic,andderivedconstitutiveexpressionlineagesin
thewild[54].Ideally,onewouldidentifyindividualswithdifferenttraitvaluesandmeasuretheirfitness.Theexpectationis
thatderivedlineageswithconstitutiveexpressionshouldexhibitthestrongestselectionfavoringthetrait.Inpractice,
however,itmightonlybepossibletomeasureacomponentoffitness,suchassurvival,matingsuccess,orfecundity,or
(evenlessdirectly)atraitthatcorrelateswiththesefitnesscomponents,suchasbodysize.Regardless,fromtheslope
andshapeoftheregressionlinerelatingphenotypetofitness(orsomeproxyoffitness),onecoulddeterminethe
(Sc.couchiiproduceonlyomnivores).Gutlength(carnivoresproduceshortergutsthan omni-vores)andgutcellproliferation(ameasureofgutperformance)werecomparedinSc.couchii, Speamultiplicata,andSp.bombifrons.Sc.couchiiproducedawiderrangeofgutlengthswhen fedshrimp[anoveldietforthisspecies,butrepresentingthederiveddiet(environment)forSpea] thanwhenfeddetritus,andthevariationwasnotdirectional:bothshorterandlongergutlengths wereproducedonshrimp.Bycontrast,bothSpeaspeciesconsistentlyproducedshorterguts whenfedshrimpthanwhenfeddetritus.Furthermore,whereasSc.couchiitadpolesdidnot exhibitincreasedgutcellproliferationwhenfedshrimp,bothSpeaspeciesdid,suggestingthat shrimpdigestionhasundergonegeneticaccommodationinSpea.
Twosubsequentstudiesfurthersupportcriteria2,3,and4[41,55].Crypticgeneticvariationin Sc.couchii was detectedwhenfed differentdiets andexposed to corticosterone(a stress hormone). These Sc. couchii tadpoles developed and grew more slowly, had increased corticosteronelevels, andexhibited greater heritability insize, development,andgut length whenfedshrimpthanwhenfeddetritus.Inaddition,Sc.couchiitadpolesexposedto cortico-steronehadgreaterheritabilityindevelopmentandgutlength.Therefore,thesestudies dem-onstratedareleaseofcrypticgeneticvariationintheancestralconditionwhentadpoleswere exposedtothederivedstimulus,andtheyalsoidentifiedapossiblehormonalmediatorofthe
carnivoreecomorph.
Moreover, this novelecomorph appearsto have undergone genetic assimilation insome derivedpopulationsofSpea.Inancestralpopulationscontainingonlyasinglespecies,both Sp.multiplicataandSp.bombifronsproducesimilar,intermediatefrequenciesofboth eco-morphs.Bycontrast,inderivedpopulationswherethesespeciesco-occur,eachbecomes nearlymonomorphic,withSp.multiplicataproducingmostlyomnivores,andSp.bombifrons producingmostlycarnivores,regardlessofresourceavailability[28].Thisnearfixationofone
ecomorphisadaptivebecauseitminimizescompetitionbetweenthetwospecies[28].More generally, this selection-driven shift from plastic to fixed ecomorph production supports
criterion3.
Note,however,thatSc.couchiimightnotrepresenttheancestralcondition(theymighthave evolvedtheomnivorefeedingstrategysecondarilyasanadaptiveresponsetocompetitionwith, orpredation by, sympatricSpea tadpoles),and supportfor criteria1–3 could therefore be questioned.Inaddition,supportforcriterion4islimited.Nevertheless,thissystemillustrateshow plasticitymight have contributedto the evolution ofa novel,complexphenotype innatural populations.
Cavefish
Eyelossincave-dwellingpopulationsofMexicantetra(Astyanaxmexicanus)alsoprovidesan excellentsettingfortestingtheplasticity-firsthypothesis.Thecaveenvironmentisan
evolution-arily novel environment, and it is known that cave populations are derived from surface populations[56].
The greatest abiotic difference between surface and cave environments (other than light availability)islowerconductivityofcavewater[43].WhensurfaceA.mexicanuswerereared underlowconductivity,theydisplayedgreatervariationineyeandorbitsize,andthey upregu-latedHSP90[43].Moreover,whenHSP90wasmanipulatedtomimicenvironmentalstress(i.e., itschaperoneabilitywasreduced),surfacefishdisplayedgreatervariationineyeandorbitsize
beyondtherangeobservedincontrols.CavefishdidnotincreasetraitvariationunderHSP90
manipulation.Inaddition,whenHSP90-manipulatedfishwiththesmallesteyeswerecrossed,
theiruntreatedF2progenyhadeyesandorbitsizesatthelowerendoftherangeinparentalfish,
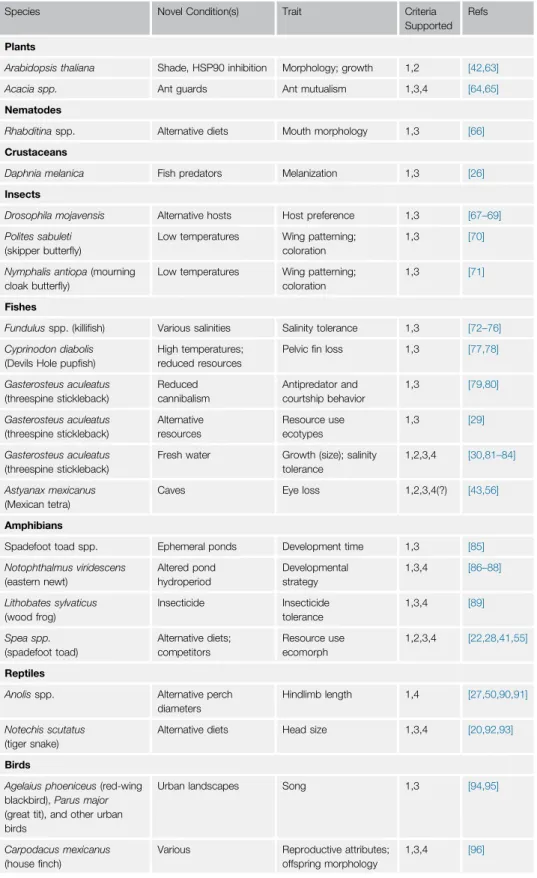
Table1.ExamplesofSpecies,Conditions,andTraitsforwhichTwoorMoreKeyCriteriaofthePlasticity-First HypothesisAreSupportedinNaturallyOccurringSystems
Species NovelCondition(s) Trait Criteria
Supported Refs
Plants
Arabidopsisthaliana Shade,HSP90inhibition Morphology;growth 1,2 [42,63]
Acaciaspp. Antguards Antmutualism 1,3,4 [64,65]
Nematodes
Rhabditinaspp. Alternativediets Mouthmorphology 1,3 [66]
Crustaceans
Daphniamelanica Fishpredators Melanization 1,3 [26]
Insects
Drosophilamojavensis Alternativehosts Hostpreference 1,3 [67–69]
Politessabuleti
(skipperbutterfly)
Lowtemperatures Wingpatterning;
coloration
1,3 [70]
Nymphalisantiopa(mourning
cloakbutterfly)
Lowtemperatures Wingpatterning;
coloration
1,3 [71]
Fishes
Fundulusspp.(killifish) Varioussalinities Salinitytolerance 1,3 [72–76]
Cyprinodondiabolis
(DevilsHolepupfish)
Hightemperatures;
reducedresources
Pelvicfinloss 1,3 [77,78]
Gasterosteusaculeatus
(threespinestickleback)
Reduced cannibalism
Antipredatorand
courtshipbehavior
1,3 [79,80]
Gasterosteusaculeatus
(threespinestickleback)
Alternative resources
Resourceuse
ecotypes
1,3 [29]
Gasterosteusaculeatus
(threespinestickleback)
Freshwater Growth(size);salinity
tolerance
1,2,3,4 [30,81–84]
Astyanaxmexicanus
(Mexicantetra)
Caves Eyeloss 1,2,3,4(?) [43,56]
Amphibians
Spadefoottoadspp. Ephemeralponds Developmenttime 1,3 [85]
Notophthalmusviridescens
(easternnewt)
Alteredpond
hydroperiod
Developmental strategy
1,3,4 [86–88]
Lithobatessylvaticus
(woodfrog)
Insecticide Insecticide
tolerance
1,3,4 [89]
Speaspp.
(spadefoottoad)
Alternativediets;
competitors
Resourceuse
ecomorph
1,2,3,4 [22,28,41,55]
Reptiles
Anolisspp. Alternativeperch diameters
Hindlimblength 1,4 [27,50,90,91]
Notechisscutatus
(tigersnake)
Alternativediets Headsize 1,3,4 [20,92,93]
Birds
Agelaiusphoeniceus(red-wing
blackbird),Parusmajor
(greattit),andotherurban
birds
Urbanlandscapes Song 1,3 [94,95]
Carpodacusmexicanus
(housefinch)
Various Reproductiveattributes;
offspringmorphology
suggest that: stressful conditions (i.e., low conductivity) induce similar changes in HSP90 functionasinlaboratorymanipulationofHSP90;bothstressfulconditionsandHSP90 manipu-lationresultinuncoveringofcrypticgeneticvariationforeyesize;andindividualsthatdevelop smalleyeswhenHSP90isinhibitedcontainallelesthatcontributetotheinheritanceofreduced eyes,evenintheabsenceoftreatment(i.e.,becomegeneticallyassimilated).
Subsequentrefinementofthisinducedeyelessphenotypeisassociatedwithimproved
func-tionality in cave conditions. When competed directly, cavefish forage better than
surface-dwellingfishinthedark[56].
Thus,the transitionfrom surfaceto cave likelyinvolved theproduction ofa range ofnovel phenotypes(satisfyingcriterion 1),whichwasfacilitatedbytheuncoveringofcrypticgenetic variation(satisfyingcriterion2).Thiswasfollowedbyselectionfavoringfixationoftheeyeless
phenotype(satisfyingcriterion3)andpossiblerefinementofthisphenotype,suchthatcavefish
outcompetesurfacefishforfoodinthedark(potentiallysatisfyingcriterion4).Note,however,
thatitisunclearifderived(cave-dwelling)populationsexhibitenhancedresourceacquisition becauseofrefinementofthefocaltraitperse(reducedeyesize;asrequiredbycriterion4)as
opposedtosomeotheraspectofthephenotype(e.g.,olfaction),whichcouldhavearisenvia newmutations.Thus,furtherstudieswillbenecessarytodetermineifcriterion4issatisfiedinthis
system.Nevertheless,thissystemagainillustrateshowplasticitymighthavecontributedtothe evolutionofacomplextrait.
EvaluatingthePlasticity-FirstHypothesisin Nature:General Assessmentof
theEvidence
Beyondthesecasestudies,researchershave(intentionallyornot)demonstratedportionsofthe plasticity-first hypothesis in numerous natural systems. Indeed, two recent reviews have
evaluatedtheempiricalsupportforplasticity-firstevolutionandfoundmanysystemsinwhich
an adaptive trait is present inancestral (orancestral-proxy) lineages as an environmentally inducedvariant[13,31],therebysatisfyingcriterion1above.However,asnotedpreviously,the mereexistenceofsuchancestralplasticityisnotsufficienttodemonstratethatatraitevolvedvia
aplasticity-firstprocess;demonstratingthat plasticity-firstevolution haslikely occurredalso
requiresevidence that any evolutionary changesin expressionof plasticity reflectselection
(criteria3and4).
InTable1 weprovide examplesofnaturallyoccurringsystemsinwhichourliteraturesurvey revealedthattwoormorecriteriawerevalidated.Ourgeneralassessmentisthatfewsystems havefulfilledallfourcriteria.Inparticular,althoughmanysystemshavesatisfiedcriteria1and3,
fewsatisfycriteria2(accumulationandrelease ofcrypticgenetic variationinancestral-proxy lineages)and4(increasedrefinementinderivedlineages).Whilecriterion2istheleastcrucialof
thefourcriteria(andamongthemostdifficult toevaluate),criterion4 isessentialtoruleout
alternativeevolutionaryexplanations(seeabove).
Wehastentoadd,however,thatalthoughfewsystemssupportallfourcriteria,takentogether thebodyofevidenceisreminiscentofDarwin'sapproachtosupporting evolutionbynatural selection[57]:multiple linesofpartial evidencepointtowardthesameprocess operatingin many,diversetaxa.
ConcludingRemarks
Wehavedescribedkeycriteriaforevaluatingtheplasticity-firsthypothesisinnaturalpopulations,
providedaroadmapfortestingthesecriteria(Figure2andBox3),andpresentedexamplesthat serveasaguidefortestingthecriteria(Table1).Moretestsareneededbeforetheplasticity-first
hypothesiscanberegardedasageneralexplanationforhowcomplex,adaptivetraitsoriginate.
OutstandingQuestions
Howpervasiveisplasticity-first
evolu-tioninnature?
Canweobserveplasticity-first
evolu-tioninreal-timeinnaturalpopulations?
Whatarethemolecularsignaturesof
theplasticity-firsthypothesis?
Areparticulartaxonomicgroups and
traitsmorelikelytoexperience
plastic-ity-firstevolutionthanothers,and,ifso,
why?
Whataretheselectiveandproximate
bases (e.g., molecular mechanisms)
of genetic accommodation and
Acknowledgments
WethankK.Pfennig,C.Ledón-Rettig,C.Martin,P.Durst,andtwoanonymousreviewersforvaluablediscussionand
comments.
References
1. Mayr,E.(1959)Theemergenceofevolutionarynovelties.In Evo-lution after Darwin (Tax,S.,ed.),pp.349–380,Universityof Chi-cagoPress
2. Wagner,G.P.andLynch,V.J.(2010)Evolutionarynovelties.Curr. Biol.20,R48–R52
3. Carroll,S.B.(2008)Evo-devoandan expandingevolutionary synthesis:agenetictheoryofmorphologicalevolution.Cell134, 25–36
4. West-Eberhard,M.J.(2003)Developmental Plasticity and Evolu-tion, OxfordUniversityPress
5. Badyaev,A.V.(2005)Stress-inducedvariationinevolution:from behaviouralplasticitytogeneticassimilation.Proc. R. Soc. B 272, 877–886
6. Pigliucci,M.et al. (2006)Phenotypicplasticityandevolutionby geneticassimilation.J. Exp. Biol. 209,2362–2367
7. Lande,R.(2009)Adaptationtoanextraordinaryenvironmentby evolutionofphenotypicplasticityandgeneticassimilation.J. Evol. Biol. 22,1435–1446
8. Pfennig,D.W.et al. (2010)Phenotypicplasticity'simpactson diversificationandspeciation.Trends Ecol. Evol. 25,459–467
9. Moczek,A.P.et al. (2011)Theroleofdevelopmentalplasticityin evolutionaryinnovation.Proc. R. Soc. B 278,2705–2713
10.Laland,K.N.etal.(2015)Theextendedevolutionarysynthesis:its structure,assumptionsandpredictions.Proc.R.Soc.B282, 20151019
11.Gilbert,S.F.etal.(2015)Eco-Evo-Devo:developmentalsymbiosis anddevelopmentalplasticityasevolutionaryagents.Nat.Rev. Genet.16,611–622
12.Whitman,D.W.andAgrawal,A.A.(2009)Whatisphenotypic plasticityand whyisitimportant?InPhenotypic Plasticity of Insects: Mechanisms and Consequences (Whitman,D.W.and Ananthakrishnan,T.N.,eds),pp.1–63,SciencePublishers
13.Schwander,T. andLeimar,O.(2011)Genesasleadersand followersinevolution.Trends Ecol. Evol. 26,143–151
14.Ghalambor,C.K.et al. (2015)Non-adaptiveplasticitypotentiates rapidadaptiveevolutionofgeneexpressioninnature.Nature 525, 372–375
15.Schlichting,C.D.andPigliucci,M.(1998)Phenotypic Evolution: A Reaction Norm Perspective, SinauerAssociates
16.Moran,N.A.(1992)Theevolutionarymaintenanceofalternative phenotypes.Am. Nat. 139,971–989
17.Mayr,E.(1963)Animal Species and Evolution, BelknapPress
18.Waddington,C.H.(1953)Geneticassimilationofanacquired character.Evolution7,118–126
19.Pfennig,D.W.andPfennig,K.S.(2012)Evolution'sWedge: Com-petitionandtheOriginsofDiversity,UniversityofCaliforniaPress
20.Aubret,F. et al.(2004)Adaptive developmentalplasticityin snakes.Nature431,261–262
21.Emlen,D.J.et al. (2007)Ontheoriginandevolutionary diver-sificationofbeetlehorns.Proc. Natl. Acad. Sci. U.S.A. 104,
8661–8668
22.Ledón-Rettig,C.C.et al. (2008)Ancestralvariationandthe potential for genetic accommodationin larval amphibians: Implicationsfortheevolutionofnovelfeedingstrategies.Evol. Dev. 10,316–325
23.Sollars,V.et al. (2003)Evidenceforanepigeneticmechanismby whichHsp90actsasacapacitorformorphologicalevolution.Nat. Genet. 33,70–74
24.Fischer,M.et al. (2004)Experimentallife-historyevolution: selec-tionongrowthformanditsplasticityinaclonalplant.J. Evol. Biol.
17,331–341
25.Suzuki,Y.andNijhout,H.F.(2006)Evolutionofapolyphenismby geneticaccommodation.Science 311,650–652
26.Scoville,A.G.andPfrender,M.E.(2010)Phenotypicplasticity facilitatesrecurrentrapidadaptation tointroducedpredators.
Proc.Natl.Acad.Sci.U.S.A.107,4260–4263
27.Losos,J.B.etal.(2000)Evolutionaryimplicationsofphenotypic plasticityinthehindlimbofthelizardAnolissagrei.Evolution54, 301–305
28.Pfennig,D.W.andMurphy,P.J.(2000)Characterdisplacementin polyphenictadpoles.Evolution54,1738–1749
29.Wund,M.A.et al. (2008)Atestofthe‘flexiblestem’modelof evolution:ancestralplasticity,geneticaccommodation,and mor-phologicaldivergenceinthethreespinesticklebackradiation.Am. Nat. 172,449–462
30.Robinson,B.W.(2013)Evolutionofgrowthbygenetic accommo-dationinIcelandicfreshwaterstickleback.Proc. R. Soc. B 280, 20132197
31.Schlichting,C.D.andWund,M.A.(2014)Phenotypicplasticityand epigenetic marking:an assessmentof evidence for genetic accommodation.Evolution 68,656–672
32.Via,S.et al. (1995)Adaptivephenotypicplasticity:consensusand controversy.Trends Ecol. Evol. 10,212–217
33.DeJong,G.(2005)Evolutionofphenotypicplasticity:patternsof plasticityandtheemergenceofecotypes.New Phytol. 166,101–
118
34.Futuyma,D.J.(2013)Evolution.(3rdedn),SinauerAssociates
35.Wray,G.A.etal.(2014)Doesevolutionarytheoryneedarethink? No,alliswell.Nature514,161–164
36.Garland,T.andKelly,S.A.(2006)Phenotypicplasticityand experi-mentalevolution.J.Exp.Biol.209,2344–2361
37.Kassen,R.(2014)Experimental Evolution and the Nature of Bio-diversity, RobertsandCompany
38.Williams,T.M.andCarroll,S.B.(2009)Geneticandmolecular insightsintothedevelopmentandevolutionofsexualdimorphism.
Nat. Rev. Genet. 10,797–804
39.Dall,S.R.X.et al. (2015)Genesascues:phenotypicintegrationof geneticandepigeneticinformationfromaDarwinianperspective.
Trends Ecol. Evol. 30,327–333
40.Ehrenreich,I.M.andPfennig,D.W.(2015)Geneticassimilation:a reviewofitspotentialproximatecausesandevolutionary conse-quences.Ann.Bot.PublishedonlineSeptember10,2015.http:// dx.doi.org/10.1093/aob/mcv130
41.Ledón-Rettig,C.C.et al. (2010)Dietandhormonalmanipulation revealcrypticgeneticvariation:implicationsfortheevolutionof novelfeedingstrategies.Proc. R. Soc. B 277,3569–3578
42.Queitsch,C.etal.(2002)Hsp90asacapacitorofphenotypic variation.Nature417,618–624
43.Rohner,N.etal.(2013)Crypticvariationinmorphological evolu-tion:HSP90asacapacitorforlossofeyesincavefish.Science
342,1372–1375
44.Moczek,A.P.(2007)Developmentalcapacitance,genetic accom-odation,andadaptiveevolution.Evol. Dev. 9,299–305
45.Paaby,A.B.andRockman,M.V.(2014)Crypticgeneticvariation: evolution'shiddensubstrate.Nat. Rev. Genet. 15,247–258
46.Ledón-Rettig,C.C.et al. (2014)Crypticgeneticvariationinnatural populations:Apredictiveframework.Integr. Comp. Biol. 54,1–11
47.Nielsen,R.(2005)Molecularsignaturesofnaturalselection.Annu. Rev. Genet. 39,197–218
48.Crispo,E.(2007)TheBaldwineffectandgeneticassimilation: revisitingtwomechanismsofevolutionarychangemediatedby phenotypicplasticity.Evolution 61,2469–2479
49.Lloyd,E.A.(1988)The Structure and Confirmation of Evolutionary Theory, GreenwoodPress
50.Kolbe,J.J.andLosos,J.B.(2005)Hind-limblengthplasticityin
51.Martin,R.A.andPfennig,D.W.(2011)Evaluatingthetargetsof selection during character displacement. Evolution 65, 2946–2958
52.Levis,N.A.et al. (2015)Aninducibleoffense:carnivoremorph tadpolesinducedbytadpolecarnivory.Ecol. Evol. 5,1405–1411
53.Pfennig,D.W.(1992)Proximateandfunctionalcausesof poly-phenisminananurantadpole.Funct. Ecol. 6,167–174
54.Pfennig,D.W.et al. (2007)Fieldandexperimentalevidencefor competition's role in phenotypic divergence. Evolution 61, 257–271
55.Ledón-Rettig,C.C.et al. (2009)Stresshormonesandthefitness consequencesassociatedwiththetransitiontoanoveldietin larvalamphibians.J. Exp. Biol. 212,3743–3750
56.Jeffery,W.R.(2008)Emergingmodelsystemsinevo-devo:
cave-fishandmicroevolutionofdevelopment.Evol. Dev. 10,265–272
57.Lloyd,E.A.(1983)ThenatureofDarwin'ssupportforthetheoryof naturalselection.Philos.Sci.50,112–129
58.Halligan,D.L.andKeightley,P.D.(2009)Spontaneousmutation accumulationstudiesinevolutionarygenetics.Annu.Rev.Ecol. Evol.Syst.40,151–172
59.Kassen,R.andBataillon,T.M.(2006)Distributionoffitnesseffects
amongbeneficialmutationsbeforeselectioninexperimental
pop-ulationsofbacteria.Nat. Genet. 38,484–488
60.Sniegowski,P.D.andLenski,R.E.(1995)Mutationand adapta-tion:thedirectedmutationcontroversyinevolutionaryperspective.
Annu. Rev. Ecol. Syst. 26,553–578
61.Renn,S.C.P.andSchumer,M.E.(2013)Geneticaccommodation andbehaviouralevolution:insightsfromgenomicstudies.Anim. Behav. 85,1012–1022
62.Sikkink,K.L.et al. (2014)Rapidevolutionofphenotypicplasticity andshiftingthresholdsofgeneticassimilationinthenematode
Caenorhabditis remanei.G3 4,1103–1112
63.Pigliucci,M.et al. (1999)Evolutionofphenotypicplasticitya comparativeapproachinthephylogeneticneighbourhoodof Ara-bidopsis thaliana.J. Evol. Biol. 12,779–791
64.Janzen,D.H.(1966)Coevolutionofmutualismbetweenantsand acaciasinCentralAmerica.Evolution 20,249–275
65.Heil,M.etal.(2004)Evolutionarychangefrominducedto constitutiveexpressionofanindirectplantresistance.Nature
430,205–208
66.Susoy,V.etal.(2015)Rapiddiversificationassociatedwitha
macroevolutionary pulse of developmental plasticity. Elife 4, e05463
67.Ruiz,A.et al. (1990)Evolutionofthemojavensis clusterof cacto-philicDrosophila withdescriptionsoftwonewspecies.J. Hered.
81,30–42
68.Matzkin,L.M.et al. (2006)Functionalgenomicsofcactushost shiftsinDrosophila mojavensis.Mol. Ecol. 15,4635–4643
69.Matzkin,L.M.(2012)Populationtranscriptomicsofcactushost shiftsinDrosophila mojavensis.Mol. Ecol. 21,2428–2439
70.Shapiro,A.M.(1975)Genetics,environment,andsubspecies differences:thecaseof Polites sabuleti (Lepidoptera:Hesperiidae).
Gt. Basin Nat. 35,33–38
71.Shapiro,A.M.(1981)Phenotypicplasticityintemperateand sub-arcticNymphalis antiopa (Nymphalidae):evidenceforadaptive canalization.J. Lepid. Soc. 35,124–131
72.Griffith,R.W.(1974)Environmentandsalinitytoleranceinthe genusFundulus.Copeia1974,319–331
73.Whitehead, A. (2010)Theevolutionary radiationof diverse osmotolerantphysiologiesinkillifish(Fundulussp.).Evolution
64,2070–2085
74.Whitehead,A.etal.(2012)Salinity-andpopulation-dependent genomeregulatory response duringosmoticacclimation in the killifish (Fundulus heteroclitus) gill. J. Exp. Biol. 215,
1293–1305
75.Whitehead,A.et al. (2011)Genomicmechanismsofevolved physiologicalplasticityinkillifishdistributedalonganenvironmental salinitygradient.Proc. Natl. Acad. Sci. U.S.A. 108,6193–6198
76.Whitehead,A.(2012)Comparativegenomicsinecological physi-ology:towardamorenuancedunderstandingofacclimationand adaptation.J. Exp. Biol. 215,884–891
77.Lema,S.C.andNevitt,G.A.(2006)Testinganecophysiological mechanismofmorphologicalplasticityinpupfishanditsrelevance toconservationeffortsforendangeredDevilsHolepupfish.J. Exp. Biol. 209,3499–3509
78.Martin,C.H.et al. (2016)DiabolicalsurvivalinDeathValley: recentpupfishcolonization,geneflowandgeneticassimilation inthesmallestspeciesrangeonearth.Proc. R. Soc. B 283, 20152334
79.Foster,S.A.(1994)Inferenceofevolutionarypattern: diversion-arydisplaysofthree-spinedsticklebacks.Behav. Ecol. 5,114–
121
80.Shaw,K.A.et al. (2007)Ancestralplasticityandtheevolutionary diversificationofcourtshipbehaviourinthreespinesticklebacks.
Anim. Behav. 73,415–422
81.Mäkinen,H.S.etal.(2006)Geneticrelationshipsamongmarine andfreshwaterpopulationsoftheEuropeanthree-spined stickle-back(Gasterosteusaculeatus)revealedbymicrosatellites.Mol. Ecol.15,1519–1534
82.McCairns,R.J.S.andBernatchez,L.(2010)Adaptivedivergence betweenfreshwaterandmarinesticklebacks:insightsintotherole ofphenotypicplasticityfromanintegratedanalysisofcandidate geneexpression.Evolution 64,1029–1047
83.Mcguigan,K.et al. (2011)Crypticgeneticvariationandbodysize evolutioninthreespinestickleback.Evolution 65,1203–1211
84.Ólafsdóttir,G.Á.et al. (2007)Postglacialintra-lacustrine diver-genceofIcelandicthreespinesticklebackmorphsinthree neo-volcaniclakes.J. Evol. Biol. 20,1870–1881
85.Gomez-Mestre,I.andBuchholz,D.R.(2006)Developmental plas-ticitymirrorsdifferencesamongtaxainspadefoottoadslinking plasticityanddiversity.Proc. Natl. Acad. Sci. U.S.A. 103,19021–
19026
86.Gabor,C.R.andNice,C.C.(2004)Geneticvariationamong populationsofeasternnewts,Notophthalmus viridescens:a preliminaryanalysisbased onallozymes.Herpetologica 60, 373–386
87.Takahashi,M.K.andParris,M.J.(2008)Lifecyclepolyphenismas afactoraffectingecologicaldivergencewithinNotophthalmus viridescens.Oecologia158,23–34
88.Takahashi,M.K.etal.(2011)Rapidchangeinlife-cycle polyphen-ismacrossasubspeciesboundaryoftheeasternnewt, Notoph-thalmus viridescens.J. Herpetol. 45,379–384
89.Hua,J.et al. (2015)Thecontributionofphenotypicplasticitytothe evolutionofinsecticidetoleranceinamphibianpopulations.Evol. Appl. 8,586–596
90.Losos,J.B.(2009)Lizards in an Evolutionary Tree: Ecology and Adaptive Radiation of Anoles, UniversityofCaliforniaPress
91.Sanger,T.J.et al. (2012)Repeatedmodificationofearlylimb morphogenesisprogrammesunderliestheconvergenceofrelative limblengthinAnolis lizards.Proc. R. Soc. B: Biol. Sci. 279,739–
748
92.Aubret,F.andShine,R.(2009)Geneticassimilationandthe postcolonizationerosionofphenotypicplasticityinislandtiger snakes.Curr. Biol. 19,1932–1936
93.Aubret,F.andShine,R.(2010)Fitnesscostsmayexplainthe post-colonisationerosionofphenotypicplasticity.J.Exp.Biol.213, 735–739
94.Hanna,D.etal.(2011)Anthropogenicnoiseaffectssongstructure inred-wingedblackbirds(Agelaiusphoeniceus).J.Exp.Biol.214, 3549–3556
95.Slabbekoorn,H.(2013)Songsofthecity:Noise-dependent spec-tralplasticityintheacousticphenotypeofurbanbirds.Anim. Behav. 85,1089–1099
96.Badyaev, A.V.(2009)Evolutionarysignificance ofphenotypic