Abstract:
Plant-microbe and microbe-microbe interactions play an important role in the modulation of plant development and productivity. Certain bacterial strains from the plant microbiome have proven to induce root growth inhibition (RGI) through pathways related to auxin, a plant regulatory hormone. However, this RGI phenotype is not induced when the plant and these strains are co-cultured with Variovorax sp. CL14. We hypothesize Variovorax sp. CL14 is able to degrade the auxin, indole-3-acetic
acid, to modulate this phenotype. Herein, we describe the construction of a genomic library of
Variovorax sp. CL14 DNA in order to identify the genes responsible for auxin degradation in this
plant-microbe-microbe interaction. Variovorax sp. CL14 genomic DNA was partially digested and the DNA fragments >10kb were inserted into a broad host range vector and transformed into E. coli 10β cells. Clones were screened for large inserts, and approximately 5000 clones were picked as the genomic library. Subsequently, the library was transformed into Burkholderia sp. CL11 and Acidovorax sp. Root219, two closely related genera to Variovorax, via triparental mating. Various screening approaches to elucidate the genes involved in auxin degradation are underway. Determining the genetic basis for this auxin degradation will enable broader understanding of how members of the genus Variovorax contribute to auxin homeostasis in the plant microbiome system.
Introduction and Background
Microbial communities play a vital role in the life of plants by influencing their physiology and development, by modulating the plant’s nutrient exchange with the surrounding environment, and by contributing to overall plant growth, health, and productivity [1]. Specifically, the rhizosphere, or the soil-root interface, is a dynamic region in which numerous biogeochemical processes take place [2]. The beneficial and competitive relationships among microbial community members and the resulting changes in host phenotype require improved definition [3]. Furthermore, increased characterization of plant-microbe and plant-microbe-plant-microbe interactions is vital to be able to promote beneficial interactions and control detrimental interactions for positive plant outcomes in agricultural settings.
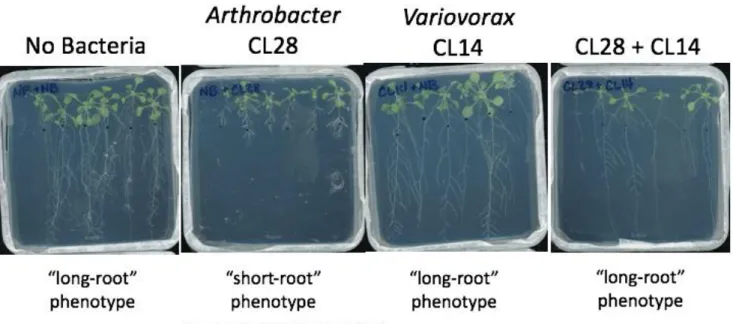
Previously, the Dangl Lab has identified a relationship between specific bacterial isolates from the Arabidopsis thaliana root microbiome, and the effect of these bacteria on Arabidopsis thaliana root length
Figure 1. Representative Agar Plate Scans Showing the Interaction between Arthrobacter sp. CL28 and Variovorax sp. CL14. Arthrobacter sp. CL28 induces a Root Growth Inhibition (RGI), or “short-root” phenotype, while when co-cultured with Variovorax sp. CL14, the phenotype is reverted.
Plant growth and development involves the integration of many environmental and endogenous signals. Plant hormones, or phytohormones, are fundamental to plant development [9]. Auxin, specifically indole-3-acetic acid (IAA), is an important mobile growth regulator in plants [4]. Both plants and plant associated microorganisms can produce IAA which modulate plant growth [5], and too much auxin is deleterious to plant growth and development [7]. Furthermore, IAA in solution is able to be quantified utilizing a colorimetric assay with Salkowski reagent [12].
Genomic libraries can be used to physically map and sequence entire genomes, clone agriculturally important genes, and to investigate gene expression patterns [14]. Constructing a genomic library involves cleaving the genome of a strain of interest and inserting a section of genomic DNA (gDNA) into a broad host range vector to create a clone [12]. Well-characterized broad host range vectors such as pBBR1-MCS2 are commonly used for gDNA insertion [10]. The entire collection of clones comprises the genomic library, which can contain up to millions of clones that can be easily sequenced [12]. The purpose of this “shotgun cloning” is to create clones that contain large samples of fragments in which at least one clone will contain the gene of interest. [13]. Genetic material within clones is able to be transferred to other strains via conjugation by triparental mating, by using helper plasmids such as pRK2013 [15]. Transferring the library to other strains may improve expression of inserted genes.
Materials & Methods
Partial Digestion of Variovorax sp. CL14 gDNA
A partial restriction digest of 100 ng/µL of Variovorax sp. CL14 genomic DNA was performed with Sau3A1 (New England Biolabs), according to the manufacturers protocol. Fifty µL digestion reactions, along with a control of gDNA without Sau3A1, were placed in a 37C water bath. Every ~5 minutes for a total of 120 minutes, a test tube was taken out and 50μL of phenol:chloroform:isoamyl alcohol (25:24:1) was added to stop the reaction, and samples were placed on ice. Each reaction was centrifuged for one minute at 14000xG and 40μL of the top aqueous phase containing partially digested gDNA removed to a new tube. The partially digested gDNA samples were run on a 0.5% agarose gel. The partial digestion was repeated using similar methodology, with larger reaction volumes. The reaction was only allowed to proceed between times 7.5 and 17.5 minutes, stopping the reaction every 2.5 minutes (Supplementary Figure 1).
Variovorax gDNA from the second digestion reaction (time points 7.5, 10, 12.5, and 15 minutes)
were pooled and separated via the BluePippin (Sage Science). Five μg of partially Sau3AI digested gDNA was run on the BluePippin in two separate runs with retention cutoffs of 15kb and 12.5kb. DNA from both runs were pooled and precipitated by adding 40μl of 3M Sodium Acetate and 900μL of Isopropanol. The samples were spun at 14000xG for 15 minutes. The supernatant was removed, and the pellet was washed twice with 70% ethanol, allowed to dry, and resuspended in water.
pBBR-1MCS2 Preparation
Cutsmart buffer, 13μL BamHI-HF, and 4μL DpnI (New England Biolabs). The digestion reaction proceeded at 37C for 3 hours and was cleaned up using the PCR cleanup kit (Qiagen). The BamHI digested pBBR1-MCS2 DNA which resulted from the restriction digest was dephosphorylated via Quick Cip (New England Biolabs), according to the manufacturer’s protocol. The dephosphorylated vector backbone was run on a 0.7% agarose gel (Supplementary Figure 2). The resulting 5kb vector backbone was isolated via gel extraction using the Qiagen Gel Extraction Kit and eluted in water.
Ligation and Transformation of plasmids
Ligation reactions were performed using ElectroLigase (New England Biolabs) with varied ratios of Blue Pippin purified SauAI partially digested Variovorax sp. CL14 gDNA and the linearized dephosphorylated BamHI cut vector backbone (1:1, 1:2, 1:3, molar ratios of plasmid: gDNA). One microliter of ligation was transformed into 50μL of NEB 10β electrocompetent E.coli cells via electroporation using the MicroPulser (BioRad) at 1.8 kV in 0.1 cm gap cuvettes. One mL of NEB 10 SOC outgrowth media was added to the electroporation cuvette and cells were transferred to 1.7mL tubes and incubated at 37C for one hour with shaking. 100μL of recovery was plated on selected on 150mm Petri dishes containing LB medium with 50 µg/mL kanamycin, 40 µg/mL X-gal, and 1 mM IPTG. White colonies were selected for further screening.
Colony PCR
Fifty μL of dH2O was added to a 96-well PCR plate and 48 random E. coli colonies were spotted
within the sterile water. This water was spotted on LB kanamycin plates to create a replicate of the E. coli clones and then the PCR plate was heated at 95C for 10 minutes to lyse the E. coli. Twenty µL PCR reactions were performed using Taq DNA polymerase, primers JMC247
PCR cycling conditions were: 95°C for 30s; 32 cycles of: 95°C for 30s, 55°C for 30s, and 68°C for 2min 30s; 68°C for 5 mins; 10°C hold. Similar methodology was used to screen the entirety of the library (~6000 clones). All PCR reactions were run on 1.5% Agarose gels.
Vector Insert Validation:
Sanger Sequencing of 23 pBBR1-MCS2 based plasmids was performed using primers JMC247 and JMC270 to obtain sequences from the pBBR1-MCS2 vector backbone into the Variovorax sp. CL14 genomic DNA insert. BLAST was used to identify the location of each end of the insert in the CL14 genome and used to calculate the size of the insert. To map the vector insert size, a double digest using EcoRV-HF and SacI-HF (New England Biolabs) was performed and run on 0.5% agarose gel. A virtual digest was performed (restrictionmapper.org) to determine the expected restriction digest pattern. The expected restriction fragment pattern was compared to the results of the double digest fragments, and to the Sanger Sequencing results.
Conjugation via Tri-parental mating
Bacterial Culture
E.coli library clones were grown in Axygen 1.1mL 96-well plates in LB media with 50 µg/mL kanamycin at 37°C. Triparental mating E. coli helper strain pRK2013 was grown at 37°C in a 100mL culture of LB with 50µL kanamycin. A rifamycin resistant strain of Burkholderia sp. CL11 and ampicillin resistant Acidovorax sp. Root219 were grown at 28°C in a 100mL 2xYT media culture with 50µg/mL rifamycin or 100µg/mL ampicillin, respectively.
Tri-parental MatingConjugation Reaction
Root219) were mixed in 96-well plates, and 4µL of the mixture was spotted onto LB plates containing no antibiotics and incubated at 28C for 24-48 hours. The mixed bacteria conjugations were streaked onto plates containing 50µg/mL rifamycin and 50µg/mL kanamycin for Burkholderia conjugations to select only rifamycin resistant Burkholderia sp. CL11 containing the pBBR1-MCS2 library plasmid, or plates containing 100µg/mL ampicillin and 50µg/mL kanamycin for Acidovorax conjugations to select only Acidovorax sp. Root219 containing the pBBR1-MCS2 library plasmid. Plates with ex-conjugants were
grown at 28°C for 2 days. Colonies were picked into LB media with the same concentrations of antibiotics used on plates and grown at 28°C. Conjugations were performed for 480 clones from the E.coli library into both Burkholderia sp. CL11 and Acidovorax sp. Root219. Insert sizes were analyzed via Sanger Sequencing and BLASTing into the Variovorax genome.
IAA Screening Protocols
Media Preparation
Condition 1 Media: To 500mL of 1X M9 Media, 1mL 1M MgSO4, 50µL 1M CaCl2, 500µL 10mM FeSO4,
2.5mL 20 mg/mL of leucine, and 1mL of 50mg/mL IAA was added. Kanamycin was added to reach a concentration of 50µg/mL. Two, 75mL aliquots were taken out.
Condition 1A Media: 75mL of Condition 1 Media.
Condition 1B Media: 75mL of Condition 1 Media, with the addition of 1.5mL of 0.75 M
Succinate.
Condition 2 Media: 500mL of Condition 1 media was made without the addition of leucine or kanamycin. Ampicillin was added to reach a final concentration of 100μg/mL. Two, 75mL aliquots were taken out.
Condition 2A Media: 75mL of Condition 2 Media with the addition of kanamycin (50 μg/µL)
Condition 2B Media: 75mL of Condition 2 Media, with the addition of kanamycin (50 μg/µL) and
Acidovorax sp. Root219 Screening Preparation
Five plates of 480 individual Acidovorax sp. Root219 conjugants were pooled into one, 96-well plate. 600µL of the pooled clones were washed twice (addition of 900µL of 1X M9 Salts, spun at 4000xG for 10 minutes, and re-suspended in 600µL of M9 media).
In a sterile, 96-well plate (Plate 1), 600µL of Condition 2 Media was added to wells A1-A6. 600µL of Condition 2A Media was added to wells A7-H12. In a second sterile plate (Plate 2), 600µL of Condition 2 Media was added to wells A1-A6, and 600µL of Condition 2B Media was added to wells A7-H12. 10µL of washed Root219 culture was added to both Plate 1 and Plate 2 (wells A9-H12). Ten µL of Variovorax sp. CL14 culture was added to wells A1-A3, 10µL of Root219 with an inserted vector was added to wells
A4-A6, and no bacteria was added to wells A6-A9 in both Plate 1 and 2. Both plates were placed in a shaking 28C incubator for 9 days.
E.coli Screening Preparation
Cultures of thirteen 96-well plates of E.coli clones were pooled into one 96-well plate containing 13 clones per well and washed as described above. In a third sterile plate (Plate 3), 600µL of Condition 1A media was added to each well. To Plate 4, 600µL of Condition 1B media was added to each well. 10µL of washed E.coli was added to each well in Plates 3 and 4. Both plates were placed in a shaking 37C incubator for 9 days.
Quantifying IAA in Clones
Fifty µL of culture from Plates 1-4 was removed and centrifuged at 4000xG for 10 minutes to remove the cells. Forty µL of supernatant was removed and added to a sterile 96-well plate. Salkowski reagent was prepared by adding 10mM FeCl3 into 35% perchloric acid. Eighty µL of Salkowski reagent
with concentrations ranging from 500µg/mL to 0µg/mL were taken, and a calibration curve was generated to convert absorbance measurements of clones with Salkowski reagent to concentrations of IAA in solution.
Results:
Construction of Variovorax genomic library
High molecular weight Variovorax sp. CL14 genomic DNA was partially digested over selected time intervals (Figure 2) to obtain varied sized DNA fragments with compatible ends for ligation in the BamHI site of the designated pBBR1-MCS2 vector backbone.
Figure 2: Gel Results from Partial Digestion of Variovorax sp. CL14 Genomic DNA. Digestion reactions started at 5 minutes (left) and ran until 120 minutes (right) in time intervals shown.
Since the amount of gDNA fragments being inserted into the vector backbone are much larger than the vector, different molar ratios of vector to gDNA were tested to see which would allow for a more effective ligation. Thus, three ligation reactions were performed, and subsequently transformed into E.coli cells. To eliminate clones with empty vectors, a blue-white screen was performed, and white colonies were selected for further screening. Through analysis of colony PCR of 96 white colonies, the ligation reaction that was found to be most successful in producing recombinant colonies with large inserts was the ligation with 1:3 molar ratio of plasmid: gDNA (Supplementary Table 3). Therefore, this ligation reaction was performed for subsequent transformations for the remainder of the experiment.
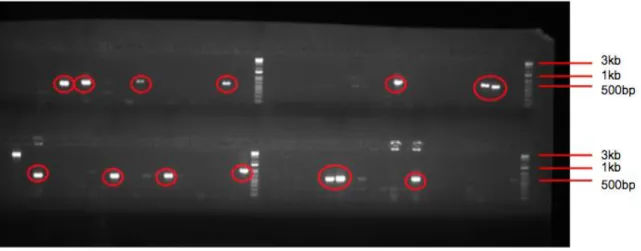
Following additional transformations and blue-white screenings, approximately 6000 white colonies were selected for the library. Based on the insert size of Variovorax gDNA in each clone, we aimed to pick enough colonies to encompass the Variovorax genome (Supplementary Figure 4). Colony PCR screening for empty vectors was performed and any sample with approximately a 500bp amplicon was eliminated (Figure 3). Following colony PCR screening of all ~6000 clones, approximately 5000 clones were found to have large inserts.
Figure 3. Gel Image of PCR Screen to Eliminate Clones with Small Inserts. Clones with ~500bp amplicons (highlighted in red) were eliminated from the library, as these clones contain empty vector.
Confirmation of Large Inserts in Library
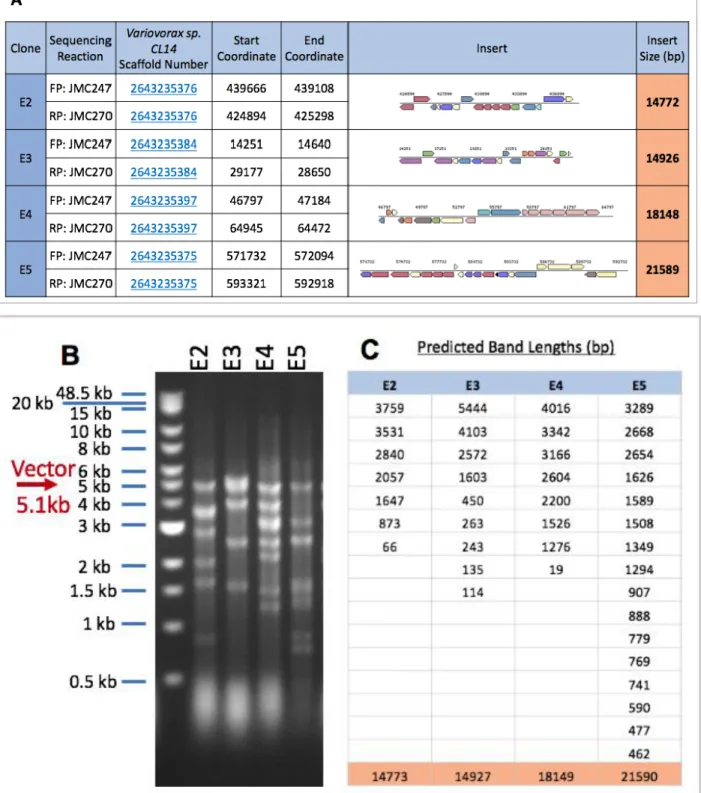
In addition to colony PCR, we sought to confirm the presence of large inserts in clones via restriction mapping and sequencing. A sample of 23 clones were randomly selected for further screening. Through Sanger sequencing into the insert, we performed BLAST analysis on the insert sequence to Variovorax sp. CL14 gDNA, and obtained a length of the insert size, and what genes were contained within the insert (Figure 4A). In addition to sequencing results, a double restriction digest was performed. Two restriction enzyme sites on pBBR-1MCS2 (EcoRV-HF and SacI-HF) flank the
BAMHI-HF site used to digest the backbone. Through a restriction digest of selected clones with these two enzymes, we were able see how much gDNA was inserted into the backbone (Figure 4B).
Figure 4: Comprehensive Data from E. coli Vector Insert Validation. (A) Sanger Sequencing Results from Sequencing into the Insert and BLASTing into CL14 genome. FP = Forward Primer and RP = Reverse Primer. Sequences for JMC247 and JMC270 can be found in Methods. Orange cells in the right-most row are the predicted insert size. (B) Cropped Gel Image from a Double Digest of pBBR-IMCS2 Backbone with Variovorax sp. CL14 gDNA inserted. (C) Predicted Band Lengths from
Transferring the E.coli vector library into Burkholderia sp. CL11 and Acidovorax sp. Root219
While we were successful in creating a large genomic library of Variovorax genes in E.coli, we were skeptical of how well these genes would be expressed with the cellular machinery in E. coli. To facilitate favorable expression of Variovorax genes during screening, we moved the genomic library into strains from two more closely related genera to Variovorax: Burkholderia sp. CL11 and Acidovorax sp. Root219 through conjugation via tri-parental mating. E. coli strain pRK2013 is necessary as the helper strain to conjugate the vectors into these recipients.
Only 136 of the 480 conjugation reactions (28%) from E. coli into Burkholderia sp. CL11
Figure 5: Sanger Sequencing Results of 4 out of 23 Selected Acidovorax sp. Root219 Clones Containing Variovorax gDNA. Uncropped data for the remaining clones is shown in Supplementary Table 4. Clones were found to have large inserts.
Screening of Library in Root219 and E.coli
Acidovorax sp. Root219 conjugations were pooled, so that multiple clones could be screened
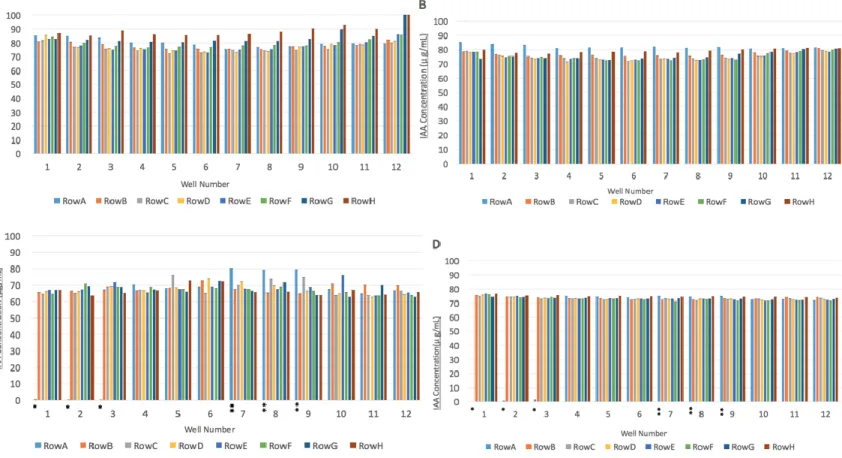
The library of E.coli was screened using similar methodology, with the exception of different media conditions (Condition 1A and 1B). Leucine was added to Condition 1 Media as E.coli 10β is a leucine auxotroph. Absorbance measures for each E.coli clone in a solution with Salkowksi’s reagent are presented in Figure 6A and 6B. No clones in the E.coli library were found to significantly degrade IAA in solution.
Figure 6. Concentration of IAA (µg/mL) in Solution of Library Clones and Salkowski Reagent.
Concentration of IAA in E.coli clones grown in Condition 1B Media (A) and Condition 1A Media (B). Concentration of IAA in Acidovorax sp. Root219 clones grown in Condition 2B Media (C) and
Discussion:
We were successful in creating a large genomic library of Variovorax sp. CL14 gDNA in E.coli. Not only is our library approximately 5000 clones, through screening procedures, we determined that clones contained large (>10kb) inserts of Variovorax gDNA. We also were successful in transferring this large library into a closely related strain of Variovorax, Acidovorax sp. Root219, where large inserts of Variovorax gDNA were retained in Root219 clones. We also characterized a preliminary screening
method to test if clones contained genes that were responsible for IAA degradation. While we did not report specific genes related to auxin degradation in Variovorax sp. CL14 due to time limitations, further experimentation would be required to do so. Large genomic libraries constructed with random insertions as described herein are limited in that it is possible clones within our initial screen in M9 media did not contain any genes which degrade auxin. Screening a larger number of clones would be necessary to encompass the entirety of the Variovorax genome, and potentially identify genes of interest.
By utilizing the construction of a genomic library to elucidate the genes responsible for IAA in Variovorax, we are limited in that we will only obtain results if the genes needed to facilitate this
metabolism are adjacent on CL14 chromosomes. While we did analyze how large the library needed to be to encompass the entirety of the CL14 genome (Supplementary Figure 4), if the genes of interest are involved in a pathway and are not next to each other on the chromosome, they would be on separate clones in the library and the functional pathway would not be reconstituted in E. coli or Acidovorax sp. Root219. The potential complexity of the auxin degradation mechanism could explain why we were unable to isolate particular genes that metabolized auxin. Furthermore, we were testing clones via a colorimetric assay in vitro, when the mechanism related to auxin degradation may require Variovorax to be present in the
Acknowledgements:
I would like to thank my thesis advisor, Dr. Jonathan Conway, for his continued mentorship and support during my time in the Dangl Lab. Thank you to Dr. Jeff Dangl for providing funding for this research. Finally, many thanks to Chloe Lindberg and Nicole Del Risco for their help in the picking and screening of the 6000 E. coli library clones.
References:
1. Mendes, Rodrigo, et al. (2013) The Rhizosphere Microbiome: Significance of Plant Beneficial, Plant Pathogenic, and Human Pathogenic Microorganisms. FEMS Microbiology Reviews, 37: 634–663. 2. McNear Jr., D. H. (2013) The Rhizosphere - Roots, Soil and Everything In Between. Nature Education Knowledge 4(3):1.
3. Aufrecht, Jayde A., et al. (2018) Quantifying the Spatiotemporal Dynamics of Plant Root Colonization by Beneficial Bacteria in a Microfluidic Habitat. Advanced Biosystems, 2(6).
4. Ottoline Leyser. (2010) The Power of Auxin in Plants. Plant Physiology 154: 501-505.
5. Yunde Zhao (2010) Auxin Biosynthesis and Its Role in Plant Development. Annual Review of Plant Biology 61: 49-64.
6. Gilbert, S., Xu, J., Acosta, K., Poulev, A., Lebeis, S., & Lam, E. (2018). Bacterial Production of Indole Related Compounds Reveals Their Role in Association Between Duckweeds and Endophytes. Frontiers in Chemistry, 6, 265.
7. Gretchen Vogel (2006) Auxin Begins to Give Up Its Secrets. Science 313: 1230-1231.
8. Atiqur Rahman, Irnayuli R. Sitepu, Sui-Yan Tang & Yasuyuki Hashidoko (2010) Salkowski’s Reagent Test as a Primary Screening Index for Functionalities of Rhizobacteria Isolated from Wild Dipterocarp Saplings Growing Naturally on Medium-Strongly Acidic Tropical Peat Soil. Bioscience, Biotechnology, and Biochemistry, 74:11, 2202-2208.
9. Gray William. M. (2004). Hormonal regulation of plant growth and development. PLoS biology, 2(9), E311.
10. Kovach et. al. (1995) Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 166: 175-176.
11. Jong-In Han, Hong-Kyu Choi, Seung-Won Lee, Paul M. Orwin, Jina Kim, Sarah L. LaRoe, Tae-gyu Kim, Jennifer O'Neil, Jared R. Leadbetter, Sang Yup Lee, Cheol-Goo Hur, Jim C. Spain, Galina Ovchinnikova, Lynne Goodwin, Cliff Han (2011) Complete Genome Sequence of the
Metabolically Versatile Plant Growth-Promoting Endophyte Variovorax paradoxus S110. Journal of Bacteriology 193(5): 1183-1190.
12. Alberts B, Johnson A, Lewis J, et al. (2002) Chapter 8: Isolating, Cloning, and Sequencing DNA. Molecular Biology of the Cell. 4th edition. New York: Garland Science.
13. Griffiths AJF, Gelbart WM, Miller JH, et al. (1999) Chapter 10: Cloning a Specific Gene Modern Genetic Analysis. New York: W. H. Freeman.
Supplementary Figures
Supplementary Figure 1: Repeated Restriction Digestion for Times Between Five minutes and 15 minutes. The first four digestion reactions (highlighted in red) were chosen to transform into the vector backbone. All times shown in minutes.
Supplementary Figure 3: Uncropped Gel from EcoRV/SACI Restriction Mapping.
Supplementary Table 3: Comprehensive Data Table Displaying Number of Clones with Small Inserts for Various Ligation Ratios. Clones were indicated as having small inserts by analyzing a gel of Colony PCR, with a “small insert” being defined as an amplicon of ~500bp.
Ligation Ratio (molar ratio plasmid: gDNA)
Ratio of Clones with Small Inserts:
Total Clones
Percentage of Clones with Small Inserts
1:1 37/96 38.54%
1:2 38/96 39.58%