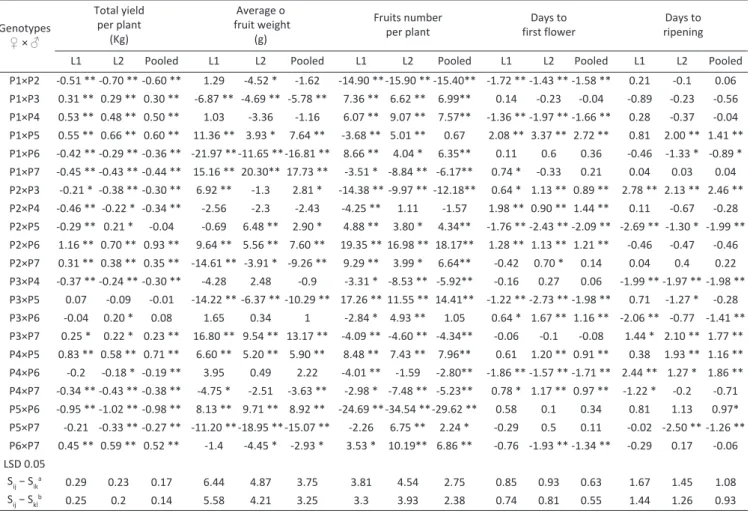
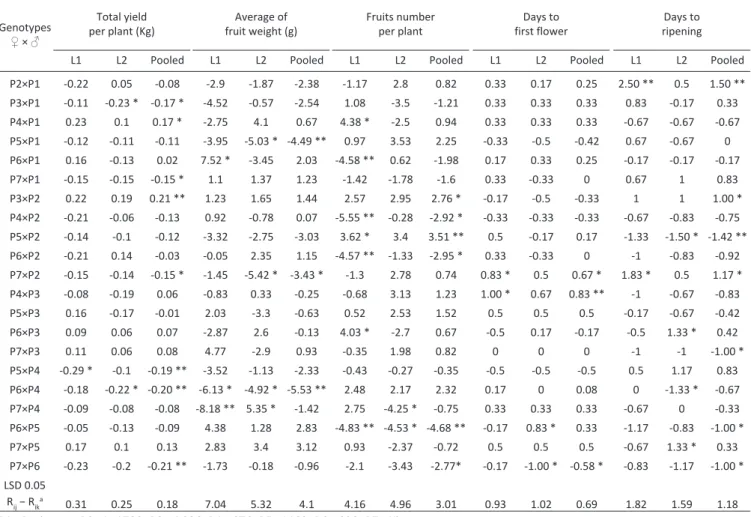
Combining ability and gene action of some tomato genotypes under low light condition
Full text
Figure

Related documents
Perform integration test based on various scenarios Tester Test Designer Test System Administrator Client test code Test scripts Test results System test
Head large as wide as width of anterior pronotal margin; anterior margin of frontoclypeus almost straight; frons regularly, rather strongly convex, vertex weakly convex, 0.9 times
We believe the most charitable thing that can be said of such statements is that they are naive in the extreme and betray a complete lack of understanding of history, of the
The projected gains over the years 2000 to 2040 in life and active life expectancies, and expected years of dependency at age 65for males and females, for alternatives I, II, and
PLOS One, Information Sciences, Journal of Economic Psychology, Journal of Economic Behavior and Organization, International Journal of Industrial Organization,
In Saudi Arabia, higher education institutions are growing rapidly and the need for quality assurance is also generating, prompting the development of the National
If a source-destination path does not exist in the routing table, a network configuration packet will be sent to request for routes to the destination node.. Figure
This paper briefs Members on the existing regulatory framework of private healthcare services, the review on the regulation of private healthcare facilities to enhance regulation
