LEABHARLANN CHOLAISTE NA TRIONOIDE, BAILE ATHA CLIATH TRINITY COLLEGE LIBRARY DUBLIN
OUscoil Atha Cliath
The University of Dublin
Terms and Conditions of Use of Digitised Theses from Trinity College Library Dublin
Copyright statement
All material supplied by Trinity College Library is protected by copyright (under the Copyright and
Related Rights Act, 2000 as amended) and other relevant Intellectual Property Rights. By accessing
and using a Digitised Thesis from Trinity College Library you acknowledge that all Intellectual Property
Rights in any Works supplied are the sole and exclusive property of the copyright and/or other I PR
holder. Specific copyright holders may not be explicitly identified. Use of materials from other sources
within a thesis should not be construed as a claim over them.
A non-exclusive, non-transferable licence is hereby granted to those using or reproducing, in whole or in
part, the material for valid purposes, providing the copyright owners are acknowledged using the normal
conventions. Where specific permission to use material is required, this is identified and such
permission must be sought from the copyright holder or agency cited.
Liability statement
By using a Digitised Thesis, I accept that Trinity College Dublin bears no legal responsibility for the
accuracy, legality or comprehensiveness of materials contained within the thesis, and that Trinity
College Dublin accepts no liability for indirect, consequential, or incidental, damages or losses arising
from use of the thesis for whatever reason. Information located in a thesis may be subject to specific
use constraints, details of which may not be explicitly described. It is the responsibility of potential and
actual users to be aware of such constraints and to abide by them. By making use of material from a
digitised thesis, you accept these copyright and disclaimer provisions. Where it is brought to the
attention of Trinity College Library that there may be a breach of copyright or other restraint, it is the
policy to withdraw or take down access to a thesis while the issue is being resolved.
Access Agreement
By using a Digitised Thesis from Trinity College Library you are bound by the following Terms &
Conditions. Please read them carefully.
The Signalling and Function of EBI2 in the Central
Nervous System
This thesis submitted to University of Dublin, Trinity College
for the degree of
Doctor of Philosophy
Aleksandra Rutkowska
O cto ber 201 4)
Supervisor:
Prof. Kumlesh K. Dev
Molecular Neuropharmacology,
D epartm ent of Physiology, School o f Medicine,
D ec la ratio n and s ta te m e n t o f plagi.irism
I declare that this thesis has not been submitted as an exercise for a degree a t this or any other university and it is entirely my own work.
I agree to deposit this thesis in the University's open access institutional repository or allow the library to do so on my behalf, subject to Irish Copyright Legislation and Trinity College Library conditions of use and acknowledgement.
Signed Date
TRWITY COLLEGE
LIBRARY DUBLIN
Acknowledgments
I am grateful to my supervisors, Prof. Kumlesh Dev and Dr. Andreas Sailer fo r th e ir continuous support and much valued advice and guidance. I am particularly thankful to Dr. Sailer fo r giving me
the opportun ity to spend the last three summers in Novartis. Thanks to his easy-going nature and passion fo r science he made my tim e spent in Novartis very productive as w/ell as enjoyable. I have learned a great deal o f techniques and had a unique opportunity to experience how science is done
in a big pharma.
M y gratitude also goes to prof. Dev fo r his ongoing guidance and support in solving everyday problems encountered in the lab. I am also grateful fo r his innovative ideas and enthusiasm w/hen
em barking new/ concepts. I am particularly thankful fo r giving me the opportun ity to contribute to several publications and also fo r the tim e spent teaching me how to w rite papers.
I would also like to thank the current and form er lab members who provided advice and experim ental support: Dr. Adam Pritchard, Sinead O'Sullivan, Cat O'Sullivan, Dr. Graham Sheridan, Dr. Debadutta Deb, Dr. Marika Doucet and Dr. Luke Healy. My thanks also go to colleagues in
T a b le o f c o n te n ts
D e c la ra tio n a n d s ta t e m e n t o f p la g ia r is m ...ii
A c k n o w le d g m e n ts ... iii
T a b le o f c o n te n ts ... iv
List o f fig u re s a n d t a b le s ... v iii T a b le o f c o m m o n a b b r e v ia tio n s ... x
S c ie n tific a b s t r a c t ...xi
Lay a b s tr a c t... x ii A im s a n d h y p o th e s is ... x iii V a lu e o f r e s e a r c h ... x iv O u tp u ts ...XV C h a p te r 1: In t r o d u c t io n ...1 7 O v e rv ie w ... 18
1 EBI2 Receptor...20
1.1 Structure and expression of EBI2... 20
1.2 Signalling o f EBI2... 20
1.3 Oxysterols...21
1.4 EBI2 function in th e im m une sy s te m ... 22
2 A stro cytes...30
2.1 Astrocyte origins and typ es... 30
2.2 Astrocyte fu n ctio n s...30
2.3 Similarities and differences b etw een rodent and human astrocytes... 31
2.4 Astrocyte involvem ent in disease... 31
3 Oligodendrocytes... 37
3.1 Oligodendrocyte m orphology and fu nction... 37
3.2 Myelin developm ent and m aintenan ce... 37
3.3 Myelin-specific and o th er proteins expressed in oligodendrocytes...38
3.4 Diseases o f o ligo d en drocytes... 39
4 EBI2 and oxysterols in disease... 43
4.1 EBI2 in disease... 43
4.2 Hereditary spastic paraplegias type 5 ...43
4.3 Alzheimer's disease... 43
4 .5 Parkinson's disease...45
4 .6 A ge-related m acular d e g e n e ra tio n ... 45
5 Closing s ta te m e n t... 47
C h a p te r 2 : M a t e r ia ls a n d m e t h o d s ...4 8 1 M a te r ia ls ... 49
1.1 C o m po u nd s... 49
1.2 A n tib o d ie s ... 49
1.3 S tains... 50
2 Anim al h usb an d ry... 50
3 Cell C u ltu re ...50
3.1 Hum an astrocyte c u ltu re ...50
3.2 M u rin e astrocyte c u ltu re ...50
3 3 Culture o f mouse organotypic slices... 51
3.4 H eterologous cell line c u ltu re ... 51
4 M o lecu lar B iology... 52
4 .1 Preparation o f m R N A ...52
4.2 Real tim e q u an titative polymerase chain reaction (RT-qPCR)... 52
5 B io ch em istry... 52
5.1 Proteom e profiler antibody a rra y s ... 52
5.2 Hom ogeneous tim e resolved fluorescence (HTRF)... 53
5.3 The enzym e-linked im m unosorbent assays (ELISAs)...53
5.4 W estern blot analysis...54
6 Cellular staining s tu d ies... 55
6.1 Fluorescence-activated cell sorting (FACS)...55
6.2 Im m u no cyto ch em istry... 55
6.3 M TT assay... 56
7 Functional Studies... 56
7.1 Calcium signalling... 56
7.2 xCELLigence im pedance assay...56
7.3 M igration and w/ound healing assays...57
8 Statistical analysis... 57
C h a p te r 3: EBI2 re g u la te s in tr a c e llu la r s ig n a llin g a n d m ig r a tio n in a s tr o c y t e s ...6 1 C hapter a im s ... 62
1 Introduction 64
2 Results... 66
2.1 EBI2 Is expressed in human and mouse astrocytes... 66
2.2 Human and mouse astrocytes express enzymes necessary fo r 7a25H C synthesis and d egradatio n ... 66
2.3 EBI2 activation induces pERK 1 /2 signalling in human astrocytes... 66
2 .4 Agonism of EBI2 induces Ca^^ signalling in human astrocytes...67
2.5 EBI2 regulates wound healing/m igration in human astrocytes... 67
2 .6 7a25H C does not induce migration in EBI2 knock-out a s tro c y te s... 68
3 Discussion... 75
3.1 Astrocytes are involved in norm al and pathophysiological processes in th e CNS...75
3.2 Astrocytes express EBI2 and 7a25H C synthesising and degrading e n z y m e s ... 75
3.3 EBI2 induces m igration/w ound healing in astrocytes... 76
C h a p te r 4: EBI2 re g u la te s lev e ls o f IL6 a n d T N F a in a s tro c y te s a n d t h e ir s ig n a llin g w ith m a c r o p h a g e s ... 7 7 Chapter a im s...78
A bstract... 79
1 In tro d u c tio n ...80
2 Results... 82
2.1 LPS does not induce cytokine release in human astrocytes... 82
2.2 7a25H C attenuates IL17/TNFa-induced levels of IL6 and NFkB translocation to th e nucleus in human astrocytes...82
2.3 EBI2 agonism inhibits IL17/TN Fa signalling in human astrocytes...83
2.4 7a25H C attenuates LPS-induced levels o f IL6 and TNFa in rodent astrocytes...83
2.5 LPS induces changes in mRNA levels of EBI2 and 7a25H C related enzymes in mouse astrocytes... 83
2.6 Mouse astrocyte conditioned m edia induces macrophage (R AW 264.7) m igration...84
3 Discussion... 91
3.1 EBI2 Inhibits pro-inflam m atory cytokine release and NFkB signalling in astrocytes...91
3.2 EBI2 regulates inter-cellular com m unication betw een astrocytes and m acrophages... 92
C h a p te r 5: EB I2 re g u la te s m y e lin d e v e lo p m e n t a n d in h ib its L P C -in d u c e d d e m y e lin a tio n . 9 3 C hapter a im s ...94
A bstract... 95
1 In tro d u c tio n ...96
2.1 Organotypic cerebellar slices maintain host tissue cytoarchitecture and physiology...99
2.2 EBI2 deficiency results in transient delay in MBP expression... 99
2.3 O lig l, CNPase and MOG are not differentially expressed in EBI2 KO and WT m ic e ...100
2.4 Downregulation o r antagonism o f EBI2 signalling staggers m ye lin a tio n ... 100
2.5 7a25HC attenuates LPC-induced demyelination in mouse cerebellar slices...100
2.6 EBI2 attenuates LPC-induced IL6 and IL1(3 release in mouse cerebellar slices... 101
3 Discussion... 109
3.1 EBI2 is involved in myelin develop m e nt... 109
3.2 Inhibition o f EBI2 signalling leads to delay in MBP expression... 110
3.3 EBI2 protects from LPC-induced demyelination via inhibition o f pro-inflam m atory cytokine release...110
Chapter 6: Discussion... 112
6.1 Study overview/...113
6.2 The role of astrocytes in CNS physiology... 113
6.3 Expression of EBI2 and 7a25HC synthesising and degrading enzymes in astrocyte biology 114 6.4 The role of EBI2 in astrocyte m ig ra tio n ...114
6.5 Regulatory role o f EBI2 and oxysterols in in fla m m a tio n ...117
6.6 The role of EBI2 in inflam m atory response in astrocytes... 117
6.7 The role of EBI2 in astrocyte-mediated cellular cross-talk... 117
6.8 EBI2 signalling in myelin developm ent... 120
6.9 The role of EBI2 in protection from chemically induced dem yelination... 120
6.10 Limitations and future directions... 123
6.11 Closing remarks... 123
List of figures and tables
C hapter 1: In troduction
Figure 1.1 Hum an, mouse, and rat EBI2 sequences 24
Figure 1.2 EBI2 phylogenetic tree 25
Figure 1.3 EBI2 expression in cells and tissue 26
Figure 1.4 EBI2 signalling pathways 27
Figure 1.5 Chemical structure and synthetic pathways of cholesterol, related oxysterols and
EBI2 antagonists
28
Figure 1.6 Function of EBI2 in th e im m une system 29
Table 1.1 Astrocyte functions 33
Figure 1.7 Receptors and ligands expressed in astrocytes 34
Table 1.2 Differences betw een human and ro d ent astrocytes 35
Figure 1.8 Cellular processes involved in m ultiple sclerosis 36
Figure 1.9 O ligodendrocyte markers expressed during developm ent 41
Table 1.3 M yelin-specific proteins and th e ir function 42
Table 1.4 EBI2 and oxysterols implicated in diseases 46
C hapter 2: M a te ria l and M etho d s
Table 2.1 List of prim ary and secondary antibodies 58
Figure 2.1 Experim ental setup using EClS and xCELLigence platforms 59
C hapter 3: EBI2 regulates intracellular signalling and m igratio n in hum an astrocytes
Figure 3.1 EBI2 is expressed in human and m ouse astrocytes 68
Figure 3.2 Human and mouse astrocytes express enzymes necessary fo r 7a25H C synthesis 69
and degradation
Figure 3.3 EBI2 activation induces ERK signalling in hum an astrocytes 70
Figure 3.4 EBI2 activation induces Ca^* signalling in hum an astrocytes 71
Figure 3.5 7a25H C induces human astrocyte w ound healing/m igration 72
Figure 3.6 7a25H C does not induces m igration in EBI2 knock-out astrocytes 73
C hapter 4: EBI2 regulates levels o f IL6 and T N F a in astrocytes and th e ir signalling w ith
___________ macrophages_____________________________________________________________
Figure 4.1 IPS does not induce release o f cytokines and chemokines in human astrocytes 84
Figure 4.2 7a25H C attenuates IL17/TN Fa-induced levels of IL6 and NFkB nuclear 85
translocation in human astrocytes
Figure 4.3 7 a25H C inhibits IL17/TN Fa-induced signalling in human astrocytes 86
Figure 4 .4 7 a25H C attenuates LPS-induced levels o f IL6 and TNFa in rodent astrocytes 87
Figure 4.5 LPS induces changes in mRNA levels o f EBI2 and 7 a25H C synthesizing and 88
degrading enzymes in mouse astrocytes
Figure 4 .6 M ouse astrocyte conditioned m edia induces m acrophage (R AW 264.7) migration 89
C hapter 5: EBI2 regulates m yelin d e v e lo p m e n t and inhibits LPC-induced d e m y e lin a tio n
Figure 5.1 Organotypic cerebellar slices m aintain host tissue cytoarchitecture and physiology 102
Figure 5.2 EBI2 deficiency results in transient delay in MBP expression 103
Figure 5.3 O lig l, CNPase and MO G are not differentially expressed in W T and EBI2 KO mice 104
Figure 5.5 7 a25H C protects from LPC-induced dem yelination
Figure 5 .6 7 a25H C attenuates LPC-induced levels o f IL ip and IL6 in cerebellar slices
106 107
C hapter 6: Discussion
Figure 6.1 EBI2 regulates intracellular signalling and m igration in astrocytes 115 Figure 6.2 EBI2 regulates levels o f IL6 and TNFa in astrocytes and th e ir signalling w ith 118
macrophages
T ab le o f c o m m o n a b b re viatio n s
Abbreviation Definition Abbreviation Definition
7a25HC
la, 25-dihydroxycholesterol
HPRTlHypoxanthine-guanine
phosphoribosyltransferase
Ab
Antibody
HSD3B7Cholest-5-ene-3P,7a-diol 3(3-
dehydrogenase
AD
Alzheimer disease
IFNInterferon
APC
Antigen presenting cell
ILInterleukin
BBB
Blood brain barrier
KOKnock-out
BCR
B cell receptor
LPCLysophosphatidylcholine
BDNF
Brain derived neurotropic factor
LPSLipopolysaccharide
BSA
Bovine serum albumin
LXRLiver X receptor
CCL/CXCL/...
Chemokine ligand
MAPMitogen-activated protein
CCR/CXCR/...
Chemokine receptors
MBPMyelin basic protein
CH25H
Cholesterol-25-hydroxylase
MOGMyelin oligodendrocyte
glycoprotein
CNS
Central nervous system
MRCMarginal reticular cell
CHO
Chinese hamster ovary cell
MSMultiple sclerosis
Cl
Cell index
MTT3-(4,5-dimethylthiazol-2-yl)-2,5-
diphenyltetrazolium bromide
CSF
Cerebrospinal fluid
NFATNuclear factor of activated I
cells
CYP7B1
Hydroxycholesterol 7-alpha-
hydroxylase
NFkB
Nuclear factor kappa B
DC
Dendritic cell
NKNatural killer cell
DMSO
Dimethyl sulfoxide
NONitric oxide
EAE
Experimental autoimmune
encephalomyelitis
OPC
Oligodendrocyte progenitor cell
EBI2
Epstein-Barr virus-induced gene
PDParkinson's disease
EBV
Z
Epstein-Barr virus
PBSPhosphate buffered saline
EClS
Electric Cell-Substrate
Impedance Sensing (EClS)
PBMC
Peripheral blood mononuclear
cell
ERK
Extracellular-signal -regulated
kinase
PLP
Proteolipid protein
FACS
Fluorescence-activated cell
sorter
Ptx
Pertussis toxin
FBS
Fetal bovine serum
ROSReactive oxygen species
FDC
Follicular dendritic cell
SIPSphingosine-l-phosphate
FGF
Fibroblast growth factor
S lP R lSIP receptor 1
GC
Germinal centre
TBSTris-buffered saline
GFAP
Glial fibrillary acidic protein
TLRToll-like receptor
GPCR
G-protein coupled receptor
TNFaTumor necrosis factor alpha
[image:11.534.16.501.70.767.2]Scientific abstract
T h e G p ro te in -c o u p le d re c e p to r EBI2 (E p s te in -B a rr viru s -in d u c e d g e n e 2) is a c tiv a te d by 7 a , 2 5 - d ih y d ro x y c h o le s te ro ! (7 a 2 5 H C ) an d plays a ro le in T c e ll-d e p e n d e n t a n tib o d y resp on se an d B cell m ig ra tio n . A b e r ra n t EBI2 signalling is im p lic a te d in a ra n g e o f a u to im m u n e disorders h o w e v e r, its
ro le in th e CNS re m a in s u n k n o w n . H e re w e c h a ra c te rize th e fu n c tio n a l ro le o f EBI2 in th e brain using p rim a ry h u m a n an d m o u se astro cy tes, o rg a n o ty p ic c e re b e lla r slice cu ltu res and EBI2 kn o ck o u t an im als. W e fin d th a t h u m a n an d m o u se astro cytes express EBI2 and th e en zym es necessary fo r synthesis an d d e g ra d a tio n o f 7 a 2 5 H C . In astro cy tes, EBI2 a c tiv a tio n s tim u la te s ERK p h o s p h o ry la tio n ,
sig nalling an d induces c e llu la r m ig ra tio n (R esu lts 1), in a d d itio n to a tte n u a tin g LPS an d IL 1 7 /T N F a -m e d ia te d re lease o f IL6 an d T N F a in h u m an and m o u se astro cytes. A c tiv a tio n o f EBI2 also inhibits IL 1 7 /T N F a -in d u c e d signalling an d N FkB tra n s lo c a tio n to th e nucleus. Im p o rta n tly , analysis o f
m R N A levels o f EBI2 an d its signalling p a th w a y in m o u se astro cy tes tr e a te d w ith LPS s h o w e d th a t th e s e cells re s p o n d to im m u n e c h allen g e in p a rt by re g u la tin g th e expression p a tte rn o f EBI2 an d
o xys tero l re la te d genes. T h e d a ta also s h o w e d th a t as tro cy tes s tim u la te d w ith LPS can in d u ce m a c ro p h a g e m ig ra tio n and th a t EBI2 can re g u la te th is c e llu la r crosstalk (R esu lts 2 ). In a d d itio n to
playing im p o r ta n t roles in a s tro c y te b iology and im m u n e system re g u la tio n in th e CNS, th e d a ta s h o w e d t h a t EBI2 signalling is involved in m y elin d e v e lo p m e n t and th a t in h ib itio n o f EBI2 signalling induces a d e la y in m y elin d e v e lo p m e n t. W e also s h o w th a t EBI2 signalling is p ro te c tiv e in LPC- ind u ced d e m y e lin a tio n in o rg a n o ty p ic c e re b e lla r slice c u ltu re s and th a t th e p ro te c tio n is, a t least
p artia lly , m e d ia te d via in h ib itio n o f p ro -in fla m m a to ry c y to k in e re lease (R esu lts 3 ). These results, fo r th e first tim e , d e m o n s tra te a ro le fo r EBI2 in a s tro c y te fu n c tio n and m yelin d e v e lo p m e n t and p ro te c tio n ag ain st d e m y e lin a tin g insults and suggest t h a t m o d u la tio n o f th is re c e p to r m a y be
Lay a bstract
The four major brain-specific cell types in the brain are; (i) neurons, which transm it and process
inform ation via electrical signals, (ii) oligodendrocytes, which insulate neuronal axons w ith fatty
sheets called myelin to ensure fast and complete electrical signal transduction, (iii) microglia, which
provide defence against potential infections in the brain and, (iv) astrocytes, th a t serve many
im portant roles such as immune defence, cleaning up debris, ensuring proper neuronal functioning
and also communication w ith other cells. These cells, and indeed all living cells, have a wide range of
proteins called receptors on th e ir surface, which are activated by signalling molecules. EBI2 is one
such receptor that plays very im portant roles in the biology of immune cells, where it regulates
antibody production to fight infections. In this thesis, we investigated w hether EBI2 is also present
on the cell surface of brain cells, namely astrocytes. We asked w hether EBI2 receptor is involved in
regulation of astrocyte physiology and fighting o f infections in the brain, and also if it can regulate
oligodendrocyte myelination o f neurons. Our research has shown fo r the first tim e that EBI2 is
present on the surface o f astrocytes and that it does play a role in immune function in the brain.
When EBI2 is in contact w ith its signalling molecule (called oxysterol 7a25HC) it causes the
astrocytes to migrate and also stops these cells from releasing harmful inflam m atory signals. The
EBI2 receptor is also capable of sensing infections and responding to them by releasing molecules
that attract immune cells into the CNS, to help fight infection. In addition, we found that the EBI2
receptor is necessary fo r appropriate and tim ely myelination of neurons and that activation o f the
EBI2 receptor can protect the cells from chemically induced loss of myelin. These findings are
im portant because they inform us about the role of the EBI2 receptor in the brain, and about how
astrocytes and oligodendrocytes are modulated by this receptor. These findings also allow us to
develop new ways to control brain cell function and lim it pro-inflam m atory processes. We believe this w ork is im portant fo r development of new drugs that might be beneficial in treating diseases
A im s an d hypothesis
The aims o f th e study are as fo llo w s:
• To investigate EBI2 expression in th e cells o f th e central nervous system, specifically
astrocytes (Results 1).
• To id e n tify w h e th e r astrocytes express enzymes necessary fo r th e EBI2 ligand, oxysterol
7a25HC, synthesis and degradation (Results 1).
• To e xplore EBI2 signalling pathw ays in astrocytes (Results 1).
• To study w h e th e r EBI2 agonism induces astrocyte m igration (Results 1).
• To exam ine if EBI2 in h ibits p ro -in fla m m a to ry cytokine release (Results 2).
• To exam ine w h e th e r activation o f EBI2 attenuates p ro -in fla m m a to ry signalling in astrocytes
(Results 2).
• To investigate w h e th e r EBI2 and 7a25HC related enzymes are regulated in response to
im m une challenge (Results 2).
• To e xplore the role o f EBI2 in astrocytes in cellular cross-talk (Results 2).
• To study th e role o f EBI2 signalling in m yelin d eve lo pm e nt in vivo and in vitro using w ild typ e and EBI2 knock-out mice (Results 3).
• To id e n tify th e role o f EBI2 signalling in m yelin deve lo pm e nt (Results 3).
• To investigate w h e th e r EBI2 protects fro m chem ically-induced d em yelination (Results 3).
• To exam ine th e role o f EBI2 in p ro -in fla m m a to ry cytokine signalling during chem ically-
induced dem yelina tion (Results 3).
W e hypothesise th a t EBI2 is expressed in astrocytes and th a t it regulates astrocyte signalling and
fu n ctio n . Specifically, we hypothesise th a t astrocytes express EBI2 and enzymes necessary fo r
7a25HC synthesis and degradation, EBI2 agonism w ith 7a25HC activates dow nstream signalling
pathways, EBI2 stim ulates m igration o f astrocytes, EBI2 alters cyto kine /che m okine levels in
astrocytes, EBI2 and 7a25H C -related enzymes are regulated in astrocytes upon im m une challenge,
and th a t EBI2 in astrocytes is involved in in te r cellular com m unication. We also hypothesise th a t EBI2
is involved in m yelin d evelopm ent, th a t EBI2 a ctivation has p ro tective effects on LPC-induced
d em yelina tion in cerebellar slices and th a t it m ight e xhibit its p ro tective effects via in h ib itio n o f pro-
V a lu e o f research
EBI2 is a fairly recently deorphanised receptor that has attracted much interest over the last 4-5
years. EBI2 is mainly expressed in the lymphoid tissue and the cells o f the immune system such as B
cells. For these reasons its function and expression in the immune system have been the main focus
of research. Astrocytes, among other functions, also serve immune functions in the CNS. They
present antigens to peripheral immune cells and release cytokines and chemokines that activate and
drive immune cell influx into the CNS. Even though astrocytes are necessary fo r a proper CNS
function and physiology, they have also been implicated in a range of neurodegenerative diseases
w ith an immune component such as m ultiple sclerosis, Alzheimer's disease or Parkinson's disease.
Moreover, cholesterol is a major component of myelin and the EBI2 agonists, oxysterols, are
cholesterol derivatives involved in its homeostasis. It is therefore warranted to investigate the
expression and function of EBI2 in the CNS and its involvement in astrocyte biology and myelination.
Therapies that control astrocyte activation may prove useful in managing neuroinflam matory or
neurodegenerative diseases and EBI2 might be a valuable tool via which inflam m atory signalling and
myelination can be controlled. This study explored the expression and function o f EBI2 in human and
mouse astrocytes and its role in myelination. The data showed th a t EBI2 is expressed in astrocytes
and th a t it plays im portant roles in the cell physiology. Activation o f EBI2 induced intracellular
signalling and cellular migration and dampened pro-inflam m atory signalling. Im portantly, this study
showed that astrocytes respond to immune challenge by regulating EBI2 and oxysterol expression.
The data also suggests that astrocytes communicate w ith the cells of the immune system, such as macrophages, via the EBI2 signalling system (the receptor and 7a25HC synthesising enzymes).
Im portantly, the data showed that EBI2 is involved in myelin development and can protect from LPC-
induced demyelination most likely via inhibition of pro-inflam m atory cytokine signalling. Taken
together, these studies support the possibility that EBI2 in astrocytes regulates inflammatory
responses and astrocyte migration. These results provide initial evidence fo r the use of EBI2 as a
novel drug target to altering astrocyte function in neuroinflam matory and neurodegenerative
Outputs
Papers
Rutkowska A., Sailer A.W. and Dev K.K. EBI2 regulates myelination state and protects from LPC-
induced dem yelination in organotypic cerebellar slice cultures. In preparation.
Preuss I., Rutkowska A., Dev K.K., and Sailer A.W. Signalling o f the oxysterol / EBI2 system in immune
cells. In preparation.
Rutkowska A., Sailer A.W. and Dev K.K. EBI2 regulates levels o f IL6 and TNFa in astrocytes and th e ir
signalling w ith macrophages. EMBO Reports, to be submitted.
Rutkowska A., Preuss I., Gessier P., Sailer A.W. and Dev K.K. EBI2 regulates intracellular signalling
and migration in human astrocytes. Glia, 10.1002/glia.22757.
Rutkowska, A., Elain, G., Jeanneau, K., Healy, L.M., M ir, A.K., Dev, K.K. SIP receptors regulate IL17-
induced levels o f IL6 in human astrocytes. Glia, in preparation.
Elain, G., Jeanneau, K., Rutkowska, A., Mir, A.K., Dev. K.K. The selective anti IL17A monoclonal
antibody Secukinumab (AIN457) attenuates IL17A-induced levels o f IL6 in human astrocytes.
Glia, 2014; 62(5):1098-1136.
Healy, L.M., Sheridan, G.K., Pritchard, A.J., Rutkowska, A., Mullershausen, F., Dev, K.K. Pathway
specific m odulation o f S lP l receptor signalling in rat and human astrocytes. British Journal o f Pharmacology, 2013; 169:1114-1129.
Doyle, P., Rohde, D., Rutkowska, A., Morgan, K., Cousins, G., McGee, H. Systematic review and m eta
analysis of the impact of depression on subsequent smoking cessation in patients w ith
coronary heart disease: 1990-2013. Psychosomatic Medicine, 2014; 76:44-57.
Posters
Rutkowska, A., Preuss, I., Sailer, A.W., Dev, K.K. EBI2 regulates intracellular signalling and migration
in human astrocytes. 9th PENS Porum o f Neuroscience, Milan, Italy, July 5-9 2014.
Rutkowska, A., Sailer, A.W., Dev, K.K. Oxysterol-EBI2 signalling in astrocytes and th e ir role in
demyelination. Astrocytes in Health and Neurodegenerative disease, a jo in t Biochemical
Society/British Neuroscience Association Pocused Meeting, London, April 28-29 2014.
Rutkowska, A., Sailer, A.W., Dev, K.K. EBI2 directs astrocyte migration and protects from
demyelination. Poster presented at 3rd Annual Meeting Frontiers in Neurology Ireland,
Dublin, Ireland, 2013.
Preuss, I., Voshol H., Abraham Y., Lasbennes MC., Laurent S., Rutkowska A., Baumgarten B., Zhang J.,
Gessier P., Dev K.K., Seuwen K., Sailer A.W. Signalling o f the oxysterol/EBI2 system in immune
cells. Poster presented at 13th International Conference on Bioactive Lipids in Cancer,
Inflammation, and Related Diseases, Puerto Rico, 2013.
Elain, G., Jeanneau, K., Rutkowska, A., Dev, K.K., M ir, A.K. IL17A and TNFa regulate levels of
chemokines and cytokines in human astrocytes via MAP kinase and NFkB signalling pathways.
Poster presented at European Committee fo r Treatment and Research in M ultiple Sclerosis
(ECTRIMS), Copenhagen, Denmark, 2013.
Rutkowska, A., Dev, K.K., Sailer, A.W. Oxysterols stimulate astrocyte migration via the EBI2 receptor.
Poster presented at the 5th School o f Medicine Postgraduate Research Day, TCD, Dublin,
Rutkowska, A., Dev, K.K., Sailer, A.W. EBI2 receptor directs human astrocyte migration. Poster presented at the NSI Conference, RCSI, Dublin, 2012.
Rutkowska, A., Dev, K.K., Mosbacher, J., Deymier, C., & Muehlemann, A. The role of SIP receptors in LPS-induced cellular stress in microglia. Poster presented at the NSI Conference, Maynooth,
2011.
Awards
Best poster aw/ard fo r a poster presented at the 3rd Annual Meeting Frontiers in Neurology Ireland, Dublin, Ireland, 2013. Rutkowska, A., Sailer, A.W., Dev, K.K. EBI2 directs astrocyte migration and protects from demyelination.
Biochemical Society Student Travel Bursary (£300) to attend Astrocytes in Health and Neurodegenerative disease conference taking place in London on April 28-29 2014.
O verview
Epstein-Barr virus (EBV)-induced gene 2 (EBI2), also known as GPR1S3, is a G protein-coupled receptor (GPCR) which is highly expressed in peripheral blood mononuclear cells (PBMCs) (Birkenbach et a!., 1993, Benned-Jensen et al., 2012). EBI2 regulates B cell positioning w ith in the
lymphoid tissue and is crucial fo r launching appropriate T cell-dependent antibody response (Gatto
et al., 2009, Gatto et al., 2011, Kelly et al., 2011, Pereira et al., 2010). In addition to being crucial fo r launching correct humoral response, EBI2 is also involved in inflam m atory responses (Gatto et al., 2011, Gatto et al., 2009, Pereira et al., 2009). EBI2 is activated by oxysterols and signals through the pertussis toxin (PTx)-sensitive heterotrim eric G proteins of the Gi/o fam ily leading to a decrease in cyclic adenosine monophosphate (cAMP) production, calcium mobilization and stim ulation of the
extracellular-signal-regulated kinase (ERK) pathway (Hannedouche et al., 2011, Liu et al., 2011,
Benned-Jensen et al., 2013, Benned-Jensen et al., 2012).
Oxysterol 7a, 25-dihydroxycholesterol (7a25HC) and closely related compounds have been identified as high affinity agonists fo r EBI2 (Liu et al., 2011, Hannedouche et al., 2011). Oxysterols are oxygenated derivatives o f cholesterol and play im portant roles in metabolism o f vitamins and lipids, cholesterol trafficking, activation of nuclear hormone receptors, regulation o f inflam m atory
signalling pathways, and many more (Schroepfer, 2000, Leoni and Caccia, 2011, Russell, 2000).
Oxysterols have chemotactic properties inducing cell migration in vitro and in vivo (Liu et al., 2011, Hannedouche et al., 2011). Moreover, oxysterols have im m unom odulatory properties having both,
pro- and anti-inflam m atory effects depending on the oxysterol and cell type (Koarai et al., 2012, Aye
et al., 2012, Diczfalusy et al., 2009, Rosklint et al., 2002, Lemaire-Ewing et al., 2009, Moog et al., 1988).
W hile the role of EBI2 in the immune system is the main focus of research its expression and function in the cells of the central nervous system (CNS) have not been investigated. Astrocytes are one o f the four resident brain cells th a t serve a range o f physiological functions in the CNS. Among other roles, they are involved in CNS immune defence. They communicate w ith cells of the immune system via signalling molecules (for example by cytokines and chemokines) and cell-cell interactions
(for example by antigen presentation) (Kimelberg and Nedergaard, 2010, Ransom et a!., 2003).
Astrocytes also isolate inflamed or injured areas by form ing glial scars. They express a wide range of receptors and release various signalling molecules allowing them to be involved in both, normal and
pathophysiological processes in the CNS (Kimelberg and Nedergaard, 2010, Ransom et al., 2003).
Im portantly, these glial cells have been implicated in many neurodegenerative diseases such as m ultiple sclerosis (MS), Parkinson's disease (PD) and Alzheimer's disease (AD) (Lin et al., 1993, Knott et al., 2002, Sidoryk-Wegrzynowicz et al., 2010, Kimelberg, 2010, Sriram, 2011).
A quarter of the to ta l body cholesterol is located in the brain o f in oligodendrocytes, neurons and astrocytes (Leoni and Caccia, 2011). Cholesterol is the main building block o f myelin and is crucial for correct and tim ely myelin form ation (Nelissen et al., 2011). Astrocytes participate in myelination
during development by upregulating cholesterol synthesis (Pfrieger and Lingerer, 2011). Oxysterols
are also involved in cholesterol homeostasis and turnover and have been shown to play im portant
roles in myelination (Nelissen et al., 2011). In addition to being involved in normal myelin
development, oxysterols can have detrim ental effects on myelin and oligodendrocytes (Makoukji et
n eurod eg en erative diseases such as Alzheim er's disease or m ultiple sclerosis (Lutjohann e t a!., 20 0 0 , Leoni e t al., 2 0 0 2 ). U pregulated levels of oxysterols have been d etected in th e brains o f EAE animals and MS patients (D iestel e t al., 2 0 0 3 ). In spastic paraplegia gene 5 (SPG5) m utations in CYP7B1 gene lead to n europ ath y o f upper m o to r neurons as w ell as p eriventricular and subcortical w h ite m a tte r lesions (Schule and Schols, 20 1 1 , Biancheri e t a!., 2009).
1 EBI2 R e c e p to r
1.1 Structure and expression o f EBI2
EBV-induced gene 2 (EBI2, also known as GPR183) is a G protein-coupled receptor (GPCR) which belongs to th e largest and most studied fam ily of GPCRs called rhodopsin-like receptors (also known as fam ily A) (Birkenbach e t a!., 1993). EBI2 is located in chrom osom e 13 in region q 32.3 very close to GPR18 which is its closest hom ologue (Rosenkilde e t a!., 2 0 0 6 ). EBI2 also has some hom ology w ith GPR17, cysteinyl leukotriene receptors and purinergic receptors (e.g. PSY2, PSY6) (N o rreg aard e t a!., 2 0 1 1 ). Am ong the most prom inent characteristics o f EBI2 structure is th e lack o f a signal peptide. M o re interestingly, th e sequence encoding human EBI2 has tw o m ethionine codons in th e N-term inus, and it is currently unclear which one is responsible fo r initiation o f translation. The m urine sequences lack th e first m ethionine but th e y do not seem to have any functional consequences in term s of expression (R osenkilde e t a l., 20 0 6 , Birkenbach e t a!., 1993, N orregaard e t a l., 20 1 1 ). Figure 1.1 shows aligned hum an, rat and mouse sequences o f EBI2 and Figure 1.2 shows EBI2 phylogenetic tree.
EBI2 is expressed at th e cell surface m ainly in lymphoid tissue (spleen and lymph nodes) and peripheral blood m ononuclear cells (PBMC) mainly on B and T cells (Rosenkilde e t al., 2 0 0 6 ). EBI2 is upregulated during B cell m aturation and is expressed in recirculating B cells in th e spleen, lymph node and bone m arrow (Pereira e t a l., 20 0 9 ). EBI2 expression is highest in naive B cells and a fte r interaction w ith T helper cells, and it is alm ost undetectable in germ inal centre (GC) B cells (Rosenkilde e t al., 2006, N orregaard e t al., 2 0 1 1 ). EBI2 expression has also been found in granulocytes, dendritic cells, macrophages, monocytes, neutrophils, human platelets, tonsils and pancreas (Birkenbach e t al., 1993, Pereira e t al., 2 0 09, Am isten e t al., 2 0 08, Chan e t a l., 2010b, H annedouche e t al., 2011, Liu e t al., 2 0 1 1 , N orregaard e t al., 2 0 1 1 ). Figure 1.3 illustrates EBI2 expression in d ifferen t tissue and cells (adapted from Rosenkilde e t al., 2006).
1.2 Signalling o f EBI2
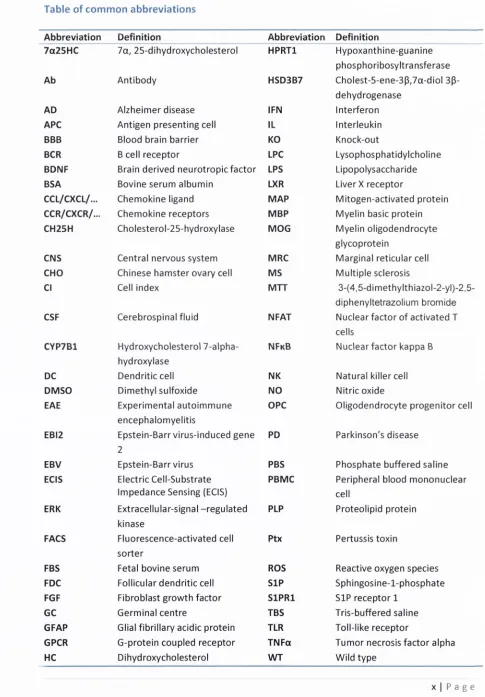
EBI2 is coupled exclusively to th e G| protein and w hen activated signals via tw o m itogen-activated protein (M AP) kinases, p38 and extracellular-signal-regulated kinase (ERK), as well as calcium and serum response e lem ent (SRE) transcription factor in a pertussis toxin (Ptx)-sensitive m anner. Notably, it does not signal via nuclear fa c to r o f activated T cells (NFAT) or nuclear factor kappa B (N FkB) (Liu e t al., 2 0 1 1 , Rosenkilde e t al., 2 0 0 6 , Benned-Jensen e t a l., 20 1 1 , Benned-Jensen e t al., 2 0 1 3 , H annedouche e t al., 2 0 11, D aug vilaite e t a!., 20 1 4 ). Figure 1.4 illustrates EBI2 signalling pathways. EBI2 is activated by several oxysterols and am ong th e m , 7 a , 25-dihydroxycholesterol (7a25H C ) has th e highest affinity and is th e most potent endogenous agonist fo r EBI2 w ith affinity ranging from 2 + /- 2 nM (H an ned o uch e e t a l., 2 0 11, Liu e t al., 20 1 1 ). O ther related oxysterols activate EBI2 w ith low er potency ranging fro m 5 + /- 4 nM fo r 7a27H C , 121 + /- 54 nM fo r 7P25HC, 3 1 0 + /- 261 nM fo r 7PHC, 1453 + /- 1163 nM fo r 7aHC, 3032 + /- 2645 nM fo r 25HC, to above 1 0,000 n M fo r cholesterol (H annedouche e t al., 2 0 1 1 ). In addition to agonists, th e re are currently tw o characterised EBI2 com petitive antagonists, NIBR189 and GSK682753A (Benned-Jensen e t a l., 20 1 3 , Gassier e t al., 20 1 4 ). Figure 1.5 shows th e chemical structure of EBI2 agonists and antagonists.
c h em o attractan t properties; indicating a ligand-induced receptor desensitization, most likely due to receptor internalization (Liu e t al., 20 1 1 , H annedouche e t aL, 2 0 1 1 ). In fact, 7a25H C induces EBI2 internalization a fte r as little as 10 m inutes o f tre a tm e n t w ith 7 a 2 5 H C (H annedouche e t a!., 2 0 1 1 , Liu e t al., 2 0 1 1 , Yi e t al., 2 0 1 2 ). O th e r related oxysterols such as 7a 2 7 H C , 7P25HC, 7aH C and 25HC also induce EBI2 expressing cell m igration although w ith much lo w er potency than 7a25H C (H an ned o uch e e t a l., 2011).
1.3 Oxysterols
Oxysterols are oxygenated derivatives o f cholesterol, which are involved in m etabolism o f vitamins and lipids, cholesterol homeostasis and trafficking, bile acid synthesis, apoptosis, necrosis, in flam m ation, im munosuppression, and many others (O taeg u i-A rrazo la e t a!., 2 0 1 0 , Bjorkhem and Diczfalusy, 2 0 02, S chroepfer, 2 0 0 0 , Leoni and Caccia, 2 0 11, Russell, 2 0 0 0 ). Oxysterols, o th e r than 7a25H C , are also agonists fo r nuclear horm one receptors such as liver x receptors (LXR) (Spann and Glass, 2 0 1 3 ). Agonism of LXR has effects on gene expression in brain cells such as astrocytes, for instance, by inducing synthesis o f ATP-binding cassette (ABC) transporters (W h itn e y e t al., 20 0 2 ). Oxysterols are fo rm ed via several pathways such as autoxidation, action of monooxygenase or by enzym atic or non-enzym atic lipid peroxidation (B jorkhem and Diczfalusy, 20 0 2 ). Oxysterols form ed by oxidation in th e ring have been found to have cytotoxic effects w hile oxysterols fo rm ed via enzym atic reaction ten d to have a range o f beneficial biological functions (O taegui-A rrazola e t al., 20 1 0 ). 7a25H C is synthesised from cholesterol through enzym atic action o f cholesteroi-25-hydroxylase (CH25H) follow ed by a cytochrom e P450 hydroxycholesterol 7-alpha-cholesteroi-25-hydroxylase (CYP7B1) and is degraded by c h olest-5-ene-3p,7a-d io l 3p-dehydrogenase (HSD3B7) (Yi e t al., 2012). Figure 1.5 shows th e synthetic pathways o f 7a25HC .
1.4 EBI2 function in the imm une system
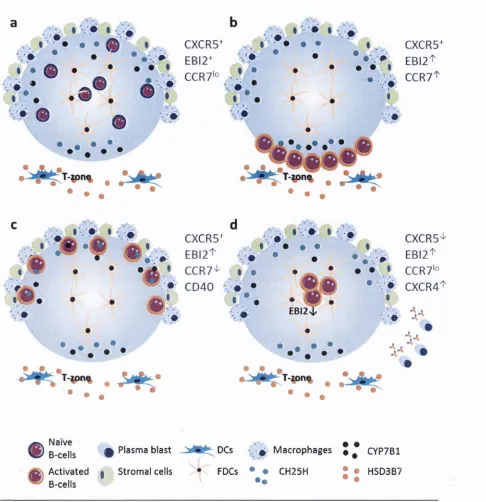
EBI2 plays very im p o rta n t roles in th e im m une system by being involved in th e correct positioning o f
activated B cells in th e lym phoid organs and launching a pp ro priate a ntib od y response (Gatto et al., 2013, Kelly et al., 2011). In brief, chem okine re ce p to r typ e 5 (CXCR5) guides naive B cells to the follicles in th e lym phoid tissue. During norm al hum oral im m une response, s h o rtly a fte r B cell
rece pto r (BCR) engagem ent, activated B cells upregulate EB12 w hich tra n sie n tly guides th e m to the
o u te r regions o f the fo llicle, possibly to e n co un te r m ore antigens e ntering fro m circulation (Kelly et al., 2011). This effe ct is counteracted by strong signals fro m chem okine receptor typ e 7 (CCR7) (also know n as E B Il) w hich is upregulate a bo ut 6 to 10 hours a fte r BCR engagem ent. CCR7 directs B cells
to th e B zone - T zone (B-T) boundary w here th e y interact w ith cognate T helper cells (Kelly et al., 2011, Pereira et al., 2010, Pereira et al., 2009, Gatto et al., 2009, Gatto et al., 2011). CXCR5 and EB12 co ntin ue to signal c o n trib u tin g to equal d is trib u tio n o f B cells along th e B-T boundary ensuring
co rrect B cell to T cell interactio ns (Kelly et al., 2011). Subsequently, fo llo w in g CCR7 d ow nre gu la tio n and cluster o f d iffe re n tia tio n 40 (CD40) engagem ent, EBI2 fu n c tio n takes over and directs cells away
fro m th e B-T boundary and to w a rd s th e o u te r- and in te r- areas o f th e fo llicle (Kelly et al., 2011). From th ere, some cells m ove to th e extra fo llic u la r areas in CXCR4- and sphing osin e-l-p ho sp ha te
(S lP l)-d e p e n d e n t m anner, d iffe re n tia te into plasmablasts and m o u n t a rapid a ntib od y response.
Some cells how ever, m ove to th e fo llicle centre, d iffe re n tia te into GC B cells and undergo antibody
a ffin ity m atu ratio n, necessary fo r a lon g-te rm a ntib od y response (Kelly et al., 2011, Pereira et al., 2010, Pereira et al., 2009, Gatto et al., 2009, Gatto et al., 2011). The signals th a t are involved in d irectin g B cells to th e GC and stim u la tin g th em to leave th e fo llicle to m o u n t a s h o rt-te rm antibody
response are still unclear. Figure 1.6 illustrates th e sequence o f events described above.
EBI2 deficiency does not a ffe ct norm al GC response, fo llic u la r organization, lym phocyte num bers or
B cell m igration to th e B-T boundary th a t occurs w ith in hours o f BCR stim ulatio n (Pereira et al., 2009, Gatto et al., 2009). However, in th e subsequent stages o f th e hum oral response EBI2 d eficient B cells fa il to move to th e o u te r- and in te r-fo llic u la r regions and instead fa vo u r th e fo llic le centre
(Gatto et al., 2009, Pereira e t al., 2009). In add itio n, EBI2 deficiency results in a decreased early phase T cell d ependent a ntib od y defence show ing th a t EBI2 is crucial fo r a pp ro priate hum oral
response before th e involvem ent o f h ig h -a ffin ity GC plasma cells occurs (Gatto et al., 2009). On the o th e r hand, EBI2 overexpression o r suppression o f its d ow nre gu la tio n a fte r T cell in te ra c tio n results
in a dim inished GC response and increased B cell m igration to th e o u te r- and in te r-fo llic u la r regions
fo llo w e d by gre ate r plasmablast fo rm a tio n and increased conce ntra tion o f antibodies in th e early
phases o f T cell dependent a ntib od y response (Gatto et al., 2009, Pereira et al., 2009).
In CH25H (cholesterol-25-hydroxylase, a 7a25HC synthesising enzyme) d eficien t mice, activated B
cells fa il to m igrate to th e o u te r- and in te r-fo llic u la r regions and fa v o u r B-T boundary and central
areas o f th e fo llicle (Hannedouche et al., 2011). These CH25H-deficient m ice also show a th re e-fo ld decrease in th e e xten t o f th e ir a ntib od y response. On th e o th e r hand, saturating the
m icro e n viro n m e n t w ith 7a25HC o r in h ib itio n o f CYP7B1 (hydroxycholesterol 7-alpha-hydroxylase,
th e second 7a25HC synthesising enzyme) results in equal d is trib u tio n o f B cells in th e lym phoid
tissue suggesting th a t 7a25HC g ra die nt is crucial fo r app ro priate localization o f B cells in the
lym phoid tissue (Hannedouche et al., 2011, Liu et al., 2011). Thus, to ensure co rrect fu n c tio n in g o f th e hum oral a ntib od y response th e g ra die nt o f 7a25HC has to be tig h tly co ntrolle d. Since B cells do
n o t them selves express the 7a25HC synthesising (CH25H and CYP7B1) nor th e degrading enzyme
lym phoid strom al upregulate th e levels of 7 a25H C via CH25H, CYP7B1 enzymes and follicular
dendritic cells dow nregulate th e levels via HSD3B7 enzym e (Yi e t al., 2012).
EBI2 is involved in various stages o f th e hum oral response exerting a w ide range of effects (Kelly e t
al., 2 0 1 1 ). As G atto and colleagues (2009) suggested, regulation of EBI2 expression may be a
p ow erfu l tool to control B cell localization w ithin secondary lymphoid tissue. This m ay be achieved
via th e B cell lym phom a 6 (BCL-6) transcriptional repressor and th e non-canonical N F-kB pathw ay
which have been found to regulate EBI2 expression (G a tto e t al., 2 0 0 9 ). It has been shown th a t
BCR-triggered NF-kB activation transiently induces upregulation o f EBI2 in naive B cells and th a t BCL-6
triggers dow nregulation of EBI2 in GC B cells (S haffer e t a l., 2 0 0 0 , G lynne e t a l., 2 0 0 0 , G atto e t al.,
2 0 0 9 ). EBI2 expression has also been found to be regulated by cytokines such as interleukin 4 (IL4),
IL6 and ILIO (S haffer e t al., 20 0 0 , Lam e t al., 2 0 0 8 , G a tto e t al., 20 0 9 ).
EBI2 expression and function have so fa r only been investigated w ith regards to th e cells o f the
im m une system and th e receptor's expression and function in th e central nervous system remains
currently unknown. H ow ever, studies suggest th a t dysregulated expression o f EBI2 or o f its related
oxysterols have been im plicated in a n um ber o f CNS diseases such as Alzheim er's disease or m ultiple
sclerosis (Papassotiropoulos e t al., 20 0 0 , B jorkhem , 20 0 6 , Fuku m o to e t al., 2 0 0 2 , Koldam ova e t al.,
2 0 0 3 , Lutjohann e t a l., 2 0 0 0 , V aya and Schipper, 2 0 0 7 , Vega e t a l., 20 0 3 ). It is th e re fo re w arranted
to investigate th e role o f EBI2 in th e CNS as w ell as th e im m une system. EBI2 involvem ent in disease
Human MDIOMANNFTPPSATPOGNDCDLYAHHSTARIVMPLHYSLVFIIGLVGNLLALVVIVONRKKIN 64
Mouse
--- T-L— SH-N--- VL--- 60
Rat
--- T-L-ASH— N--- L--- 60
Human
STTLYSTNLVISDILFTTALPTRIAYYAMGFDWRIGDALCRITALVFYINTYAGVNFMTC 124
Mouse
M---L---V--- 120
Rat
M---V
L---L--- 120
Human
LSIDRFIAVVHPLRYNKIKRIEHAKGVCIFVWILVFAOTLPLLINPMSKOEAERITCMEY 184
Mouse
F---Y---LS---LT----E-GDKT---180
Rat
F---Y---I-V--- LK---DKT---- 180
Human
PNFEETKSLPWILLGACFIGYVLPLIIILICYSOICCKLFRTAKONPLTEKSGVNKKALN 244
Mouse
G-A---LL--- ITV— L--- 240
Rat
G-A--- LL---A
L--- 240
Human
TIILIIVVFVLCFTPYHVAIIQHMIKKLRFSNFLECSORHSFOISLHFTVCLMNFNCCMD 304
Mouse
1--- M-CSPGA-- GA--- 300
Rat
G---M -- V-T-YAPGA-G-GV--- 300
Human
PFIYFFACKGYKRKVMRMLKRQVSVSISSAVKSAPEENSREMTETOMMIHSKSSNGK
361
Mouse
--- K--- R--- S---A --- R
357
Rat
--- K--- R--- S---A --- R
357
Figure 1.1 H um an, m ouse, and ra t EBI2 sequences. Alignm ent of hum an, mouse, and rat EBI2
sequences (h ttp ://b la s t.n c b i.n lm .n ih .g o v /B la s tA lig n .c g i). Amino acid residues th a t are identical in
hum an EBI2 are indicated by dashes; differences are shown. The transm em brane dom ains are
[image:25.534.55.495.71.365.2]Monkey (0.0003)
Human (0.0024)
Bovine (0.0413)
Chicken (0.1644)
--- Zebra fish (0.1792)
■Tetraodon (0.1337)
- Mouse (0.0351)
Rat (0.0293)
[image:26.534.31.511.51.641.2]Lym phoid organs: spleen, lymph node; PBMCs: B cells, T cells (Including CD4*, CD8^), natural killer (NK) cells, monocytes H e a rt (ao rta, atriu m )
Lung
G astro in testin al tra c t (appendix)
U ro g e n ita l system (placenta, bladder)
Glands (salivary gland)
Brain (corpus callosum, medulla oblongata)
7a25HC
Arresi
p38 cAMP
E R K l/2 1
NFkI SRE
[image:28.534.35.520.75.465.2]a CH25H CYP7B1 HSD367
Cholesterol ---^ 25HC ► 7a25H C 4-cholesten-7a25^
ol-3-one
OH
Cholesterol 25HC
HO'
HO' HO' 'OH
b
7aHC ■'OH
HO'
OH
7a27H C
'OH HO'
7P25HC
HO' ’OH
OH
7P27HC
HO' 'OH
N BR189
Cl
i
O ^ NCl
O GSK682753A^'
Figure 1.5 Chemical structures and synthetic pathways of (a) cholesterol, 25HC and 7a25HC and (b)
other EBI2 activating oxysterols. (c) Chemical structure of EBI2 antagonists, NIBR189 and
[image:29.534.22.506.49.567.2]. . • % • * 9 '
I ,
CXCR5+
CXCR5"
■ > • • * EBI2^ E B I 2 ^
I • • • • • • r l * • • • • •
a®
• •
•-• • • W ^ •
•
•
•
•
*
.
•
' ■■ I ' i 1 . '■ ■ ■ i " i . ■ ■■
.
.
CXCR5^
■
•
•
'•■■•
CXCR5^
vaJ
« • >
EB,2t
• ;
eb
,2^
« W
•
•
• ^ . *
CCR7^
• •
• _
•
• •
•
CCR7'°
• CD40 V # •
> CXCR4^
EBI2T
.
1
4
®
N aive « «B-cells 0 Plasma blast DCs 0. M a c ro p h a g e s CYP7B1
©
A c tiv a te d | S tro m a l cells • FDCs • • C H 25H J • H SD 3B 7B-cells • • •
[image:30.534.31.517.30.532.2]2 Astrocytes
2.1 Astrocyte origins and types
Astrocytes are one o f the three well known types of resident CNS glial cells alongside microglia and
oligodendrocytes (and not withstanding others such as glial antigen 2 (NG2) positive glia). Astrocytes
are homeostatic cells of the CNS characterised by a star-shaped morphology and spreading
numerous extended processes that surround neurons and blood vessels (Wang and Bordey, 2008,
KImelberg, 2010, Butt et a!., 2002). They are non-excitable and are identified by interm ediate
filaments called glial fibrillary acidic proteins (GFAP) (Wang and Bordey, 2008, KImelberg, 2010). Of
importance, it has to be noted that not all astrocytes have a star-shaped morphology or express
GFAP (Kimelberg, 2004).
Astrocytes originate from neural progentitors that give rise to both, macroglia (astrocytes and
oligodendrocytes) and neurons (Verkhratsky and Butt, 2013). The neural progenitors differentiate
into radial glial cells in the ventricular zone and can give rise to astrocytes, neurons and some
oligodendrocytes. Most oligodendrocytes, however, are generated in distinct areas of the CNS from
specific glial precursors. Astrocytes, on the other hand, originate from both, radial glial cells and glial
precursors (Verkhratsky and Butt, 2013).
Astrocytes are an exceptionally varied cell type and many subtypes that distinguish one type from the other can be named based on th e ir function, cellular presence, morphology, gene expression,
physiological properties and response to stim uli (Verkhratsky and Butt, 2013). One classification
divides these cells into type 1 and type 2 astrocytes. Type 1 astrocytes are defined as fibroblast-type cells that originate early in development and secrete growth factors that, in turn, can stim ulate the
growth o f type 2 astrocytes (Raff et al., 1983, Armstrong et al., 1990). Type 2 astrocytes can
differentiate into 0-2A progenitor cells and give rise to oligodendrocytes (Raff et al., 1983,
Armstrong et a!., 1990). Other classification divides astrocytes into protoplasmic astrocytes, found in
the grey m atter of the brain and the spinal cord, and fibrous astrocytes, found in the w hite m atter in
the brain, the spinal cord, optic nerve and retina (Verkhratsky and Butt, 2013). These are the most
numerous subtypes o f astrocytes, however, there are other types which include radial glia and other
radial glia-type cells such as M uller glia in the retina and Bergmann glia in the cerebellum
(Verkhratsky and Butt, 2013). In addition, there are other, much less abundant subtypes which
include: perivascular and marginal astrocytes, velate astrocytes o f the cerebellum and olfactory bulb,
interlam inar and polarised astrocytes found in the primate cortex, and varicose projection astrocytes
present only in human brains (Verkhratsky and Butt, 2013). Other subtypes include tanacytes found
in the periventricular organs, hypothalamus and the spinal cord, pituicytes in the pituitary gland, and
retinal and ventrical cells such as ependymocytes, choroid plexus cells and retinal pigment epithelial
cells (Verkhratsky and Butt, 2013).
2.2 Astrocyte functions
Astrocytes are a very abundant cell type in the brain and serve a range o f physiological functions. For
instance, they control neuronal m aturation, form and maintain synapses, uptake neurotransmitters,
release antioxidants and are responsible fo r extracellular ion buffering (Kimelberg and Nedergaard,
2010, Wang and Bordey, 2008, Ransom et al., 2003, Kimelberg, 2010). Astrocytes prom ote
myelination and speed up myelin wrapping around axons and form ation o f compact myelin during
expression is necessary fo r correct nnyelination and blood-brain barrier form ation (Liedtke e t al., 19 9 6 ). In addition to supporting neurons, astrocytes also play a role in m aintenance of th e BBB and com m unication w ith th e im m u ne system via antigen presentation and cytokine/chem okine production (K im elberg and N e d erg aard , 2 0 1 0 , W an g and Bordey, 2 0 0 8 , Ransom e t al., 2003, K im elberg, 20 1 0 ). They also regulate synaptic transmission by processing and integrating neuronal activity and coordinating calcium waves, suggesting astrocyte involvem ent in intercellular com m unication (K im elberg and N edergaard , 2 0 1 0 , W an g and Bordey, 2 0 0 8 , Ransom e t al., 2003, K im elberg, 20 1 0 ). Table 1.1 lists various types o f astrocyte functions.
Astrocytes also express a w id e range o f receptors such as G protein coupled receptors (GPCRs), ionotropic receptors as well as receptors fo r grow th factors, chemokines, cytokines, steroids, toll-like receptors (TLRs), and, them selves, release neurotransm itters, grow th factors and o th e r signalling m olecules allowing th em to be involved in both normal and pathophysiological processes taking place in th e CNS (K im elberg and N edergaard , 2 0 1 0 , W an g and Bordey, 2 0 0 8 , Ransom e t al., 2003, K im elberg, 20 1 0 ). Figure 1.7 illustrates some o f th e large rep ertoire o f proteins expressed by astrocytes.
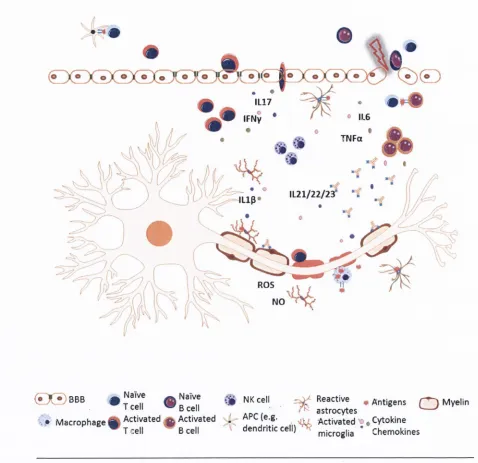
During CNS dam age or injury astrocytes facilitate healing of th e injured tissue, form glial scars to lim it th e exten t o f dam age and release pro-inflam m atory cytokines and chemokines leading to activation of microglia and influx o f leukocytes into th e CNS. Astrocytes have also been found to present myelin antigens to T cells in MS (Fontana e t al., 1984). Although others have shown th a t only microglia, and not astrocytes, present myelin basic protein (M BP) antigens to T cells and astrocytes are involved in suppression of T cell proliferation and release of an ti-inflam m atory cytokines but not in phagocytosis (M a ts u m o to e t al., 1992).
2.3 S im ilarities and differences b e tw e e n ro d e n t and hum an astrocytes
Part of o ur knowledge about astrocyte biology and function comes from in vitro studies th a t use mouse or rat prim ary cells. W h ile hum an and rodent brain cells share many similarities th e re are also differences b etw een species th a t need to be taken into account w hen conducting research (O b e rh e im e t al., 20 0 9 ). For instance, both rodent and human astrocytes respond to m etabotropic agonists by intracellular Ca^* changes. On th e o th e r hand, hum an astrocytes are m ore varied in m orphology and are approxim ately 2.6-fold larger than rodent astrocytes (O berheim e t al., 2009). M u rin e and hum an astrocytes also express d ifferen t receptors. For instance, mouse astrocytes express a w ide range o f to ll-like receptors (TLRs) while human astrocytes preferentially express TLRS and do not express TLR4 receptors (Farina e t al., 2 0 0 5 ). These differences in cell biology may account fo r th e fact th a t m any compounds fail w hen tested in humans in spite o f excellent th e ra p e u tic effects in animals. Table 1.2 lists th e similarities and differences b etw een rodent and hum an astrocytes (O b e rh e im e t al., 20 0 9 ).
2 .4 Astrocyte in v o lv e m e n t in disease
inflam m atory infiltrates (D'Amelio et al., 1990, Morcos et al., 2003, Smith and Sommer, 1992). Figure 1.8 illustrates how astrocytes exacerbate the pathophysiology of m ultiple sclerosis. Astrocytes also contribute to neurodegeneration in Parkinson's disease via release of toxic molecules such as l-M ethyl-4-phenylpyridinium (MPP^) that is taken up by dopaminergic neurons and causes neurotoxicity (Rappold and Tieu, 2010). In Alzheimer's disease, (3-amyloid plaques activate astrocytes, which in turn secrete pro-inflam m atory cytokines that exacerbate the neuronal injury taking place during Alzheimer's disease (Johnstone et a!., 1999).
There is a bi-directional communication between astrocytes and peripheral immune cells via cytokine/chemokine system and other signalling molecules. At the site of injury, astrocytes release cytokines and chemoklnes to activate resident immune cells and induce influx o f leukocytes from the periphery (Miljkovic et al., 2011, Miljkovic et al., 2007). They stimulate interferon gamma (IFN-y) and IL17 production by T helper 1 (T h l) and T h l7 cells in m ultiple sclerosis plaques and respond to the stim ulation w ith these cytokines by increasing expression of inducible nitric oxide (iNOS), NO and other pro-inflam m atory mediators such as IL6 implicated in m ultiple sclerosis pathogenesis (Miljkovic et al., 2011, Bo et al., 1994, Miljkovic et al., 2007). Increased levels of IL6 enhance its own production in a positive feedback loop and also drive IL17 synthesis and release (Ma et al., 2010, Meares et al., 2012, McGeachy et al., 2007). Some studies have suggested that astrocytes also drive the development o f EAE by phagocytosing myelin and presenting myelin basic protein (MBP), myelin oligodendrocyte glycoprotein (MOG) and proteolipid protein (PLP) antigens to CD4* T cells. However, the role o f astrocytes and th e ir phagocytosing capabilities remain a topic of debate in the field (Tan et al., 1998, Kort et al., 2006, Lee et al., 1990, Fontana et al., 1984).