0095-1137/08/$08.00⫹0 doi:10.1128/JCM.00343-08
Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Characterization of Small-Colony Variants of
Enterococcus faecalis
Isolated from Chickens with Amyloid Arthropathy
䌤
Andreas Petersen,* Mark S. Chadfield, Jens P. Christensen, Henrik Christensen, and Magne Bisgaard
University of Copenhagen, Faculty of Life Sciences, Department of Veterinary Pathobiology, Stigbøjlen 4, 1870 Frederiksberg C, Denmark
Received 19 February 2008/Returned for modification 14 April 2008/Accepted 19 June 2008
In this study we report the isolation and characterization of normal-sized and small-colony variants of
Enterococcus faecalisfrom outbreaks of amyloid arthropathy in chickens. Postmortem examinations of 59
chickens revealed orange deposits in the knee joints, typical for amyloid arthropathy. Bacterial cultures from 102 joints and 43 spleens exhibited pure (nⴝ88) and mixed (nⴝ11) cultures of normal (nⴝ60) and pinpoint (nⴝ28) colonies ofE.faecalis. Pulsed-field gel electrophoresis of 62 isolates demonstrated seven different band patterns with at most two band size variations, and multilocus sequence typing demonstrated two different sequence types, sharing six out of seven alleles, suggesting a close evolutionary relationship between isolates obtained from four outbreaks. In addition, all isolates were clonally related to an amyloid arthropathy reference strain from The Netherlands, previously shown to be globally dispersed. Initial investigation of the isolated small-colony variant phenotype revealed no difference in whole-cell protein profiling between normal and pinpoint colonies. However, the pinpoint colony isolates appeared to be more virulent in an in vivo challenge model in chickens than their normal-sized-colony counterparts. In addition, pinpoint morphology and associated slow growth were expressed without reversion after in vitro and in vivo passage, suggesting a genuine altered phenotype, and in some instances normal colonies converted to pinpoint morphology postinfection. In conclusion, small-colony variants of
E. faecalis are described for the first time from veterinary clinical sources and in relation to amyloid
arthropathy in chickens.
Small-colony variants (SCV) are slowly growing subpopu-lations of bacteria with distinctive phenotypic and patho-genic traits. They exhibit atypical colony morphology and other unusual phenotypic characteristics (15). SCV seem to be better able to persist intracellularly and to be less sus-ceptible to antibiotics than their wild-type counterparts (28). In addition, they may cause latent and recurrent infections, and a low growth rate and other unusual phenotypic char-acteristics make them a challenge for clinical microbiolo-gists to identify. SCV have mostly been studied for
Staphy-lococcus aureusbut have also been reported to occur among
Pseudomonas aeruginosa(12),Vibrio cholerae(30),Brucella
melitensis(13), Escherichia coli(9), Salmonella enterica
se-rovar Typhimurium (32), and Neisseria gonorrhoeae (25) strains in addition to other genera and species. So far, SCV
for Enterococcus spp. have been reported only from a
hu-man clinical case of endocarditis (14).
Amyloid arthropathy was first described in brown layers (18). Since then, unilateral or bilateral polyarticular amyloid arthropathy frequently accompanied by systemic amyloidosis involving mainly the liver and spleen has been reported exten-sively in brown layers and less frequently in broiler parents (33).Enterococcus faecalishas been the most frequently dem-onstrated pathogen in cases of field outbreaks and is consid-ered to have a significant role in pathogenesis (17).
Experi-mental infection studies withE.faecalisare able to reproduce the typical lesions observed in the joints (19, 20). Infections occur only when the arthropathic and amyloidogenic strain is administered intravenously, intra-articularly, or intraperitone-ally; however, natural infection routes remain to be elucidated (19). Natural outbreaks associated withE.faecalisseem to be clonal or clonally related as demonstrated by pulsed-field gel electrophoresis (PFGE) (33). Recently, a specific clone was demonstrated by multilocus sequence typing (MLST) to be associated with amyloid arthropathy worldwide (26). Other pathogens have also been reported from naturally occurring cases (21).
In this study we report for the first time the isolation of
E. faecalis, demonstrating pinpoint colony morphology, from
Lohmann Silver chickens with amyloid arthropathy. Further characterization of the isolates including comparisons to nor-mal-colony morphology is reported, including animal passage.
MATERIALS AND METHODS
Diagnostic submissions.Lohmann Silver layers aged between 24 and 37 weeks with clinical signs of arthritis were received during a period of 6 months (January to June 2004) from four independent farms with free-range egg production. From each farm between one and four submissions were received, constituting a total of 59 chickens. All flocks affected originated from the same parent stock but were reared on three epidemiologically distinct farms. All chickens received were subjected to a full postmortem examination. Material for further investigations was obtained upon indication.
Lesions (joints and spleen) were swabbed with a sterile cotton wool swab and subsequently plated onto blood agar (blood agar base, CM55; Oxoid, Basing-stoke, Hampshire, United Kingdom) containing 5% sterile bovine blood. Stan-dard methods were used for incubation and subsequent identification of patho-gens associated with lesions observed (34). A total of 102 joints and 43 spleens were sampled and subsequently cultured bacteriologically.
* Corresponding author. Mailing address: University of Copenhagen, Faculty of Life Sciences, Department of Veterinary Pathobiology, Stig-bøjlen 4, 1870 Frederiksberg C, Denmark. Phone: 45 35333686. Fax: 45 35332757. E-mail: [email protected].
䌤Published ahead of print on 25 June 2008.
2686
on May 16, 2020 by guest
http://jcm.asm.org/
16S rRNA gene sequencing.16S rRNA gene sequencing of strains C19315/Led 5A (normal-colony morphology) and C19315/Led 5B pp (SCV) was performed as reported previously (3, 8). A BLAST search (1) was performed in GenBank (5) to document the relationship to other sequences. Pairwise comparisons for similarity were performed by the program WATER included in EMBOSS (31).
PFGE.A total of 62 strains, including three reference strains ofE.faecalis, namely, 6085.94, 1244.94, and 199-2558-9EW, previously associated with re-ported cases of chicken amyloid arthropathy (18), were included in the PFGE analysis. PFGE conditions were as described previously (7) with digestion of DNA with SmaI.
MLST.MLST was performed on 12 outbreak isolates, representing different PFGE types, and the reference strain 6085.94. Primers and PCR conditions were as described for the typing scheme published at theE.faecalisMLST home page (http://efaecalis.mlst.net/), with the exception of the primers published for the
gydgene. It was not possible for us to generate a sequence from the PCR product; therefore, a new set of primers was designed which were used for both the PCR amplification and the sequencing of the amplicon. The primers were gydFnew (5⬘-CAAACCATGAAACATTAACTGGA-3⬘) and gydRnew (5⬘-AA GTTAGCGAAGTATTCTAAAGTACGA-3⬘). These primers amplify a larger part of thegydgene, surrounding the part which is amplified by the primers in the published MLST scheme. Amplification products were purified in Microspin S-400 HR columns (GE Healthcare, Buckinghamshire, United Kingdom) and sequenced in both directions using the PCR primers. Sequencing was performed at Macrogen Inc., Seoul, Korea. The obtained sequences were compared to published alleles at theE.faecalisMLST home page (efaecalis.mlst.net), and a sequence type (ST) was assigned to each strain. Sequences and strain informa-tion were deposited at the same website.
Protein analysis. Preparation of whole-cell protein samples for protein profiling was carried out as reported previously (6). The NuPAGE Bis-Tris electrophoresis system (Invitrogen, United Kingdom) was used for separation of proteins which subsequently were stained with Simply Blue (Invitrogen, United Kingdom) following the manufacturer’s instructions. Six isolates were analyzed, including reference strain 6085.94.
Chicken MHC haplotyping.To investigate whether affected birds in a flock were genetically predisposed to disease compared with those not affected, major histocompatibility complex (MHC) haplotyping using polymorphic microsatellite marker LEI0258 was performed according to the method of Fulton et al. (11). Included in the analysis were 20 affected and 20 nonaffected birds from the same flock (farm I).
Experimental infection.Strains chosen were C19315/Led 1b pp (SCV) and C19315/Led 1a (normal-colony morphology). Both isolates were originally iso-lated from the same chicken suffering from arthritis and amyloid arthropathy and demonstrated an identical PFGE type. Cultures were taken from⫺80°C, incu-bated overnight at 37°C on blood agar base (CM0055; Oxoid) containing 5% sterile bovine blood, and subsequently cultured overnight in 10 ml brain heart infusion (Oxoid) at 37°C with shaking. Optical density was measured at 620 nm and adjusted to 0.1, to equate to a bacterial concentration of approximately 1⫻ 108CFU/ml. The bacterial concentrations were confirmed by plate counts. A total of 81 11-week-old Lohmann brown layers obtained from a commercial farm having a high level of biosecurity without a history of disease or increased mortality were randomly divided into seven groups as shown in Table 4. After 1 week of acclimatization the birds were inoculated intravenously or subcutane-ously or in the footpad with 0.5 ml buffered peptone water containingE.faecalis
C19315/Led 1b pp orE.faecalisC19315/Led 1a. The control group was inocu-lated in the same way with sterile buffered peptone water. All birds were
pro-vided water and a standard feed ad libitum. Groups were separated and kept under free-range housing conditions without outdoor access. Three weeks after infection all birds were killed and examined as explained under “Diagnostic submissions.” Experimental conditions were approved by the Danish Animal Ethics Committee.
Nucleotide sequence accession number.For reasons given below, only the sequence of strain C19315 Led 5A was deposited in GenBank, under accession number EU285587.
RESULTS
Mortality.A total of four flocks of Lohmann Silver chickens were investigated. Three of the flocks suffered from increased first-week mortality (2.5 to 3.2%) while the fourth flock dem-onstrated 1.6% first-week mortality, compared to a normal first-week mortality of less than 1% (M. Bisgaard, personal communication). A slightly increased mortality was also ob-served at the ages of 4 to 5, 5 to 6, 6 to 9, and 9 weeks, with total mortalities during rearing constituting 7.0, 3.3, 2.5, and 5%, respectively. The reasons for these mortalities were, however, not investigated.
Pathology. Although retrospective investigations showed that all four flocks demonstrated some unevenness at the end of rearing, which resulted in culling of small birds before trans-portation to egg-laying farms, none of the owners submitted birds for examinations. However, during the start of egg pro-duction clinical signs indicative of arthritis became obvious and the flocks turned more uneven. Up to 13% of affected birds were culled, with the remaining healthy birds reported to de-velop normal production with no further signs of disease.
The submitted affected birds were emaciated, demonstrating unilateral or bilateral enlarged femorotibial and/or tibiometa-tarsal joints. With the exception of a few hip joints other joints were normal. Upon postmortem examination, deposits of or-ange material typical of amyloid were demonstrated within the superficial layers of articular cartilage and periarticular capsu-lar structures. Cartilage surfaces were eroded, with erosions sometimes extending into bone tissue, resulting in osteomyeli-tis. The menisci of the stifle joints also showed orange deposits. Synovial fluid was increased, containing floccular greyish yel-low exudates. Surprisingly, tendons and tendon sheaths re-mained unaffected.
[image:2.585.46.543.82.202.2]Spleens were swollen, demonstrating proliferation of the white pulpa. In addition, hepatomegaly and renomegaly were often observed, with affected livers and kidneys showing a bronze to slightly yellowish color.
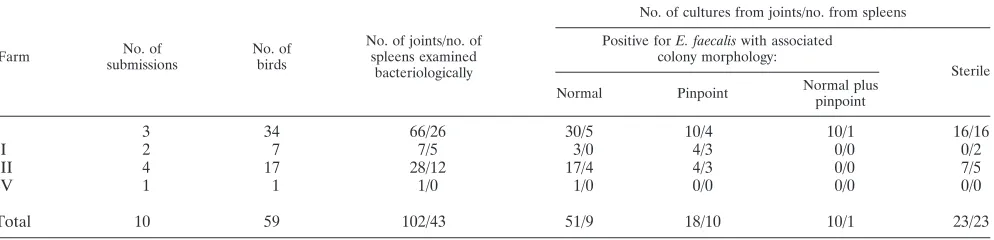
TABLE 1. Bacteriological examinations of joints and spleens of submitted animals from affected farms
Farm No. of
submissions
No. of birds
No. of joints/no. of spleens examined
bacteriologically
No. of cultures from joints/no. from spleens
Positive forE. faecaliswith associated colony morphology:
Sterile
Normal Pinpoint Normal plus pinpoint
I 3 34 66/26 30/5 10/4 10/1 16/16
II 2 7 7/5 3/0 4/3 0/0 0/2
III 4 17 28/12 17/4 4/3 0/0 7/5
IV 1 1 1/0 1/0 0/0 0/0 0/0
Total 10 59 102/43 51/9 18/10 10/1 23/23
on May 16, 2020 by guest
http://jcm.asm.org/
Bacteriology.The bacteriological examinations of joints and spleens resulted in positive cultures from 79 out of 102 joint samples examined and 20 out of 43 spleens examined (Table 1). Two types of pure cultures ofE.faecaliswere demonstrated after 24 h of incubation, normal and pinpoint sized (Fig. 1). The proportion of pinpoint-sized colonies was greatest in the spleens (approximately 50% of positive spleens). In the joints approximately 25% of the positive samples demonstrated pure cultures of pinpoint-sizedE.faecaliscolonies (Table 1).
16S rRNA sequencing. Two strains, C19315/Led 5A (nor-mal-colony morphology) and C19315/Led 5B pp (SCV), iso-lated from the same animal, were identical in their nucleotide sequence (1,455 bp compared) and showed 99% similarity with other published 16S rRNA gene sequences forE.faecalis in-cluding the type strain. For this reason only the sequence of strain C19315 Led 5A was deposited in GenBank under acces-sion number EU285587.
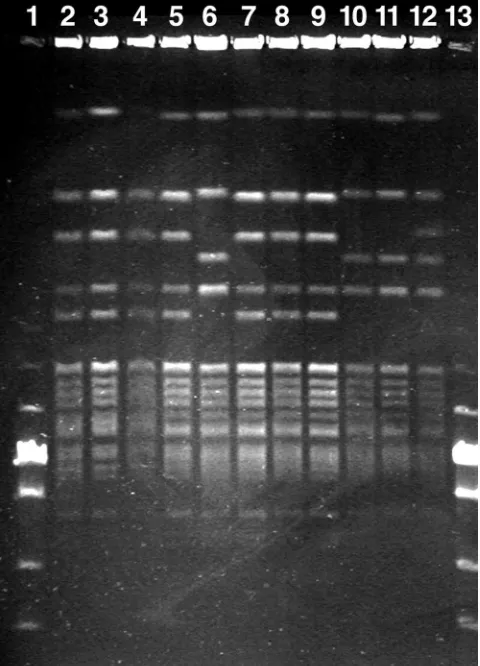
PFGE.A subset of 62 isolates was typed using PFGE. The typing demonstrated seven different band patterns (Table 2 and Fig. 2). All seven patterns were subtypes of the typical pattern demonstrated by the included reference strains (pat-tern B in this study) and differed at most with two band size variations. The most common band pattern (A) was demon-strated in 32 isolates, from all four farms.
MLST.Most strains included in the MLST were identified as ST82, including the amyloid arthropathy reference strain
6085.94 (Table 3). The only exceptions were the two strains with PFGE type D, which demonstrated a new sequence type, ST170. This new sequence type was identical in six out of seven alleles and thus was closely related to the remaining analyzed isolates.
[image:3.585.45.284.70.210.2]FIG. 1. Primary plate with mixed culture of pinpoint (predomi-nant) and normal-sized colonies ofEnterococcus faecalis.
[image:3.585.302.542.71.404.2]FIG. 2. PFGE of Danish outbreak isolates ofEnterococcus faecalis associated with amyloid arthropathy. Lanes 1 and 13, PFGE Low Range marker set (New England Biolabs); lanes 2 to 5 and 7 to 9, PFGE type A; lanes 6, 10, and 11, PFGE type B; lane 12, PFGE type C.
TABLE 2. Distribution of PFGE types ofE.faecalisfrom different farms and three amyloid arthropathy reference strains
Strain group (farm or reference)
No. of isolates
Total
PFGE typea
A B C D E F G
I 51 25 15 1 5 1 1 3
II 2 2 0 0 0 0 0 0
III 5 4 1 0 0 0 0 0
IV 1 1 0 0 0 0 0 0
Reference (strains 085.94, 1244.94, and 199-2558-9EW)
3 0 3 0 0 0 0 0
a
All PFGE types are subvariants of the reference type denoted B (33).
TABLE 3. Comparison of STs and PFGE types
Isolate ST PFGE typea
Reference 6085.94 ST82 B
C19315/Led 5A ST170b D
C19315/Led 4A ST82 A
C19445/Led1PPc ST82 NDd
OTB/Led 13 ST82 A
C19363/4 Led 2 ST170 D
SC 3 ST82 B
C19363-11/Led 1 ST82 ND
C19315/Led 1A ST82 E
C19315/Led 1a ST82 B
C19315/Led 1b ppc ST82 B
C19363-18/Led 1 ST82 G
C19315/Led 2A ST82 B
a
All PFGE types are subvariants of the type denoted B.
b
ST170 is identical to ST82 in six out of seven alleles.
c
SCV.
d
ND, not determined.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:3.585.302.541.553.693.2]Protein analysis.No band pattern differences were observed among isolates demonstrating normal and pinpoint-size colony morphology (data not shown).
MHC.Three allele sizes for marker LEI0258 were demon-strated (252 bp, 312 bp, and 364 bp). Affected chickens dem-onstrated seven alleles of 252 bp, 16 alleles of 312 bp, and six alleles of 364 bp. Corresponding numbers for nonaffected chickens were 7, 20, and 5, respectively. No statistical differ-ence was found between affected and nonaffected birds (P⫽
0.82, chi-squared test), indicating that the two groups were similar in their MHCs.
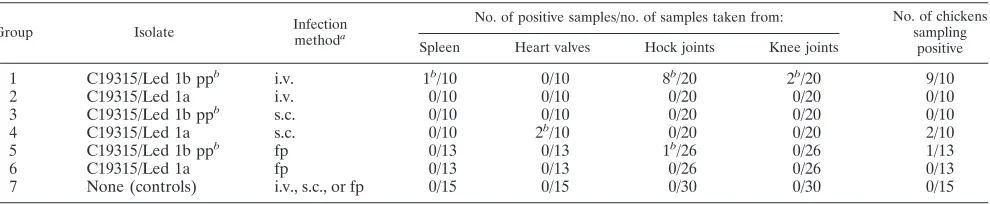
Experimental infection.Groups 1, 3, and 5 (Table 4) were inoculated intravenously or subcutaneously or in the footpad, respectively, with isolate C19315/Led 1b pp (7.8⫻108CFU),
demonstrating pinpoint colonies, while groups 2, 4, and 6 were inoculated with isolate C19315/Led 1a (2.0⫻108CFU), which
showed normal colony morphology. In group 2, one bird died 2 weeks postinfection. Lesions in the liver were observed, but all bacteriological samples from this bird were sterile. In group 1, eight birds demonstrated increased fluid in hock joints. No other pathological lesions were observed when birds were sac-rificed 3 weeks postinfection. Bacteriological examinations of the sacrificed birds revealed that for the intravenous inocula-tion, 9 out of 10 birds infected with pinpoint colonies sampled positive with recovery of the pinpoint colony morphology after 3 weeks. This was in contrast to none of the birds being fected with normal-sized colonies (Table 4). Subcutaneous in-fection resulted in positive sampling from 2 out of 10 animals inoculated with normal-size colonies. However, the reisolated strains demonstrated pinpoint-size colony morphology.E.
fae-caliswas reisolated from only one of a total of 26 birds
inoc-ulated in the footpad (Table 4). The inocinoc-ulated and reisolated strains both demonstrated pinpoint-size colony morphology. All control birds sampled negative and demonstrated no clin-ical or pathologclin-ical signs of disease.
DISCUSSION
In this study it was demonstrated that SCV ofE.faecalismay be associated with outbreaks of amyloid arthropathy in chick-ens. To our knowledge, this is the first time that SCV ofE.
faecalishave been associated with lesions in veterinary
medi-cine. Kaase et al. (14) described SCV of E. faecalisfrom a human endocarditis patient. It was reported that the pheno-type was not stably expressed, as a portion of the bacteria reverted to a normal-sized phenotype (14). SCV have
previ-ously been observed among a wide range of bacteria (see the review by Proctor et al. [29]). Our observations indicate that SCV remain stable in vitro under laboratory conditions just as pinpoint colonies inoculated in vivo were reisolated 3 weeks postinfection. However, normal colonies inoculated subcuta-neously were in two cases reisolated as pinpoint colonies 3 weeks later. It has been demonstrated in vivo that the SCV phenotype may be induced by exposure to sulfamethoxazole-trimethoprim (2), suggesting that the formation of SCV in vivo may be a response to the presence of antibiotics. However, in our study the experimental animals did not receive any antibi-otic treatment.
SCV of S. aureus appear to be less virulent than normal strains but persist as well as parent strains do in animal models and human infections. The chicken infection study carried out demonstrated a difference between the two morphology types. When chickens were inoculated intravenously, only chickens infected with the pinpoint-sizedE. faecalissampled positive after 3 weeks while all chickens inoculated with normal-sized bacteria sampled negative (Table 4). No significant difference between the two test strains was demonstrated for the other two infection routes (subcutaneously and in the footpad). Amyloid deposits were not demonstrated in the infection ex-periment. The same observation was made by Landman et al. (23), who observed chickens 10 weeks post-infection withE.
faecalis. A chronic inflammatory process will increase the
chances for amyloid arthropathy, and thus, longer observation periods may be needed (23). In contrast, histological amyloid deposits were demonstrated between synovial cells and in the superficial layer of articular surfaces 5 days post-intravenous infection by the same research group (20). In group 1 in our study, increased synovial fluid was observed in all birds sam-pling positive forE.faecalis. Splenomegaly, however, was ob-served in only one out of nine affected birds, indicating a slow progression of amyloid arthropathy.
[image:4.585.47.542.80.182.2]PFGE, MLST, and whole-cell protein analysis did not reveal any differences between normal- and pinpoint-sized colony morphology types ofE.faecalisisolated from the same animal. Also the 16S rRNA sequences were identical for a set of isolates (pinpoint and normal) from the same animal. The association of pinpoint colonies with amyloid arthropathy, in contrast to previously described strains with normal colony morphology, could not be explained by protein profiling. Since the pinpoint phenotype appeared stable, the mechanism be-hind it is difficult to explain as a result of downregulation of
TABLE 4. Study design and reisolation ofEnterococcus faecalisfrom Lohmann brown layers, 3 weeks postinfection
Group Isolate Infection
methoda
No. of positive samples/no. of samples taken from: No. of chickens sampling
positive Spleen Heart valves Hock joints Knee joints
1 C19315/Led 1b ppb i.v. 1b/10 0/10 8b/20 2b/20 9/10
2 C19315/Led 1a i.v. 0/10 0/10 0/20 0/20 0/10
3 C19315/Led 1b ppb s.c. 0/10 0/10 0/20 0/20 0/10
4 C19315/Led 1a s.c. 0/10 2b/10 0/20 0/20 2/10
5 C19315/Led 1b ppb fp 0/13 0/13 1b/26 0/26 1/13
6 C19315/Led 1a fp 0/13 0/13 0/26 0/26 0/13
7 None (controls) i.v., s.c., or fp 0/15 0/15 0/30 0/30 0/15
a
Abbreviations: i.v., intravenous; s.c., subcutaneous; fp, footpad.
b
Isolates demonstrating small-colony morphology.
on May 16, 2020 by guest
http://jcm.asm.org/
production of surface structures. Although slightly larger col-onies were observed after 48 h of incubation, it seems likely that the pinpoint colonies should be explained as a result of a decreased growth potential. Previous studies of SCV in other bacterial species have indicated defects in pathways that are involved in electron transport (4). Further investigations are required to clarify the mechanisms behind the small-colony morphology of theE.faecalisstrains isolated in this study.
Previous studies have indicated that different susceptibilities to Marek’s disease may be correlated with different haplotypes (10). Considering the low prevalence of amyloid arthropathy observed in affected flocks, it might be suggested that only certain MHC types are susceptible to amyloid arthropathy. MHC haplotyping with marker LEI0258 revealed no difference between affected and nonaffected chickens from the same flock, suggesting that genetic difference between the two groups is un-likely.
The two molecular typing methods, PFGE and MLST, dem-onstrated different discriminatory powers. The PFGE was able to specify different subgroups of isolates while most isolates included in the MLST demonstrated the same ST or, in the few exceptions, a closely related ST. By MLST, the genes encoding enzymes involved in conserved metabolic and information pro-cesses are compared at the DNA sequence level (24) and results are portable and easily comparable via Internet-based servers.E.faecalisisolates demonstrating ST82 were first de-scribed from a human clinical infection (16) and have recently been shown to be associated with amyloid arthropathy world-wide (26).
In this study amyloid arthropathy is reported from four flocks of Lohmann Silver layer birds. Field cases of amyloid arthropathy have predominately been demonstrated in brown layers and only occasionally in broiler parents (35). The four outbreaks described in this study indicate that the disease may be more common among layer lines other than brown layers than previously assumed. However, all affected birds in this study could be traced back to the same hatchery and parent flock.
Although amyloid arthropathy has been reproduced exper-imentally, the pathogenesis remains speculative (19, 20, 22, 23). Previous indications and the present experimental infec-tions clearly indicate a slowly developing disease. The possi-bility of vertical transmission has been suggested by Petersen et al. (26) to explain the existence of a global clone. The increased first-week mortality observed in three of the four affected flocks seems to support this hypothesis. However, increased mortality was also observed subsequently for all four flocks affected.
Several putative virulence genes have been identified for isolates ofE.faecalisinfecting humans (27). Similar investiga-tions have not yet been performed with avian isolates. Further studies are required to clarify the virulence properties of the isolates, including both the normal-sized-colony isolates and the SCV.
In conclusion, SCV ofEnterococcus faecaliswere obtained from lesions in chickens associated with amyloid arthropathy. To our knowledge this is the first veterinary report and the first characterization of this phenotype amongE. faecalisstrains. Preliminary molecular characterizations could not detect any differences from normal-sized counterparts. However,
infec-tion experiments suggested a higher infecinfec-tion potency for the SCV than for ordinaryE.faecalis.
ACKNOWLEDGMENTS
This research was supported by grants from The Danish Poultry Council (project 23) and The Danish Research Council for Technology and Production Sciences (274-05-0358).
W. J. M. Landman is thanked for donation of reference strains. Tony Bønnelycke, Gitte Petersen, and Katrine Madsen are thanked for skillful technical assistance.
REFERENCES
1.Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller, and D. J. Lipman.1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res.25:3389–3402. 2.Anderson, S. W., J. R. Stapp, J. L. Burns, and X. Qin.2007. Characterization
of small-colony-variantStenotrophomonas maltophiliaisolated from the spu-tum specimens of five patients with cystic fibrosis. J. Clin. Microbiol.45:529– 535.
3.Angen, O., P. Ahrens, P. Kuhnert, H. Christensen, and R. Mutters.2003. Proposal ofHistophilus somnigen. nov., sp. nov. for the three species incer-tae sedis ‘Haemophilus somnus,’ ‘Haemophilus agni’ and ‘Histophilus ovis’. Int. J. Syst. Evol. Microbiol.53:1449–1456.
4.Balwit, J. M., P. Vanlangevelde, J. M. Vann, and R. A. Proctor.1994. Gentamicin-resistant menadione and hemin auxotrophicStaphylococcus au-reuspersist within cultured endothelial cells. J. Infect. Dis.170:1033–1037. 5.Benson, D. A., I. Karsch-Mizrachi, D. J. Lipman, J. Ostell, and D. L.
Wheeler.2007. GenBank. Nucleic Acids Res.35:D21–D25.
6.Bisgaard, M., D. J. Brown, M. Costas, and M. Ganner.1993. Whole cell protein profiling ofActinobacillus-like strains classified as taxon 2 and taxon 3 according to Bisgaard. Zentralbl. Bakteriol.279:92–103.
7.Chadfield, M. S., J. P. Christensen, J. Juhl-Hansen, H. Christensen, and M. Bisgaard.2005. Characterization ofEnterococcus hiraeoutbreaks in broiler flocks demonstrating increased mortality because of septicemia and endo-carditis and/or altered production parameters. Avian Dis.49:16–23. 8.Christensen, H., M. Bisgaard, O. Angen, and J. E. Olsen.2002. Final
classification of Bisgaard taxon 9 asActinobacillus arthritidissp. nov. and recognition of a novel genomospecies for equine strains ofActinobacillus lignieresii. Int. J. Syst. Evol. Microbiol.52:1239–1246.
9.Colwell, C. A.1946. Small colony variants ofEscherichia coli. J. Bacteriol.
52:417–422.
10.Dalgaard, T. S., L. Vitved, K. Skjodt, B. Thomsen, R. Labouriau, K. H. Jensen, and H. R. Juul-Madsen.2005. Molecular characterization of major histocompatibility complex class I (B-F) mRNA variants from chickens dif-fering in resistance to Marek’s disease. Scand. J. Immunol.62:259–270. 11.Fulton, J. E., H. R. Juul-Madsen, C. M. Ashwell, A. M. McCarron, J. A.
Arthur, N. P. O’Sullivan, and R. L. Taylor, Jr.2006. Molecular genotype identification of theGallus gallusmajor histocompatibility complex. Immu-nogenetics58:407–421.
12.Gerber, A. U., and W. A. Craig.1980. Small colony variants ofPseudomonas aeruginosa—characterization and virulence in normal and leukopenic mice. Clin. Res.28:A735.
13.Jacob, J., G. M. Hort, P. Overhoff, and M. E. A. Mielke.2006.In vitroandin vivocharacterization of smooth small colony variants ofBrucella abortusS19. Microbes Infect.8:363–371.
14.Kaase, M., A. Anders, and S. G. Gatermann. 2004. First description of small-colony variants ofEnterococcus faecalisisolated from an endocarditis patient. Int. J. Med. Microbiol.294:146.
15.Kahl, B. C., G. Belling, R. Reichelt, M. Herrmann, R. A. Proctor, and G. Peters.2003. Thymidine-dependent small-colony variants ofStaphylococcus aureusexhibit gross morphological and ultrastructural changes consistent with impaired cell separation. J. Clin. Microbiol.41:410–413.
16.Kawalec, M., Z. Pietras, E. Danilowicz, A. Jakubczak, M. Gniadkowski, W. Hryniewicz, and R. J. L. Willems.2007. Clonal structure ofEnterococcus faecalisisolated from Polish hospitals: characterization of epidemic clones. J. Clin. Microbiol.45:147–153.
17.Landman, W. J., A. E. vd Bogaard, P. Doornenbal, P. C. Tooten, A. R. Elbers, and E. Gruys.1998. The role of various agents in chicken amyloid arthrop-athy. Amyloid5:266–278.
18.Landman, W. J. M., E. Gruys, and R. M. Dwars.1994. A syndrome associ-ated with growth depression and amyloid arthropathy in layers: a preliminary report. Avian Pathol.23:461–470.
19.Landman, W. J. M., D. R. Mekkes, R. Chamanza, P. Doornenbal, and E. Gruys.1999. Arthropathic and amyloidogenicEnterococcus faecalis infec-tions in brown layers: a study on infection routes. Avian Pathol.28:545–557. 20.Landman, W. J. M., N. H. M. T. Peperkamp, C. A. M. Koch, P. C. J. Tooten, P. A. P. Crauwels, and E. Gruys.1997. Induction of amyloid arthropathy in chickens. Amyloid4:87–97.
21.Landman, W. J. M., and A. Feberwee.2001. Field studies of the association
on May 16, 2020 by guest
http://jcm.asm.org/
between amyloid arthropathy andMycoplasma synoviaeinfection, and ex-perimental reproduction of the condition in brown layers. Avian Pathol.
30:629–639.
22.Landman, W. J. M., K. T. Veldman, D. J. Mevius, and J. H. H. van Eck.2001. Aerosol transmission of arthropathic and amyloidogenicEnterococcus fae-calis. Avian Dis.45:1014–1023.
23.Landman, W. J. M., K. T. Veldman, D. J. Mevius, and J. H. H. van Eck.2003. Investigations ofEnterococcus faecalis-induced bacteraemia in brown layer pullets through different inoculation routes in relation to the production of arthritis. Avian Pathol.32:463–471.
24.Maiden, M. C., J. A. Bygraves, E. Feil, G. Morelli, J. E. Russell, R. Urwin, Q. Zhang, J. Zhou, K. Zurth, D. A. Caugant, I. M. Feavers, M. Achtman, and B. G. Spratt.1998. Multilocus sequence typing: a portable approach to the identification of clones within populations of pathogenic microorganisms. Proc. Natl. Acad. Sci. USA95:3140–3145.
25.Morton, H. E., and J. Shoemaker.1945. The identification of Neisseria gonorrhoeaeby means of bacterial variation and the detection of small colony forms in clinical material. J. Bacteriol.50:585–587.
26.Petersen, A., H. Christensen, and M. Bisgaard.2007. Investigations on the existence of a global clone of Enterococcus faecalis associated with amyloid arthropathy in chickens, abstr. 09-022, p.254. Abstr. 15th World Vet. Poult. Congr. World Veterinary Poultry Association, Beijing, China.
27.Pillar, C. M., and M. S. Gilmore.2004. Enterococcal virulence—pathoge-nicity island ofE. faecalis. Front. Biosci.9:2335–2346.
28.Proctor, R. A., P. van Langevelde, M. Kristjansson, J. N. Maslow, and R. D.
Arbeit.1995. Persistent and relapsing infections associated with small-colony variants ofStaphylococcus aureus. Clin. Infect. Dis.20:95–102.
29.Proctor, R. A., C. von Eiff, B. C. Kahl, K. Becker, P. McNamara, M. Herr-mann, and G. Peters.2006. Small colony variants: a pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat. Rev. Micro-biol.4:295–305.
30.Rashid, M. H., C. Rajanna, D. L. Zhang, V. Pasquale, L. S. Magder, A. Ali, S. Dumontet, and D. K. R. Karaolis.2004. Role of exopolysaccharide, the rugose phenotype and VpsR in the pathogenesis of epidemicVibrio cholerae. FEMS Microbiol. Lett.230:105–113.
31.Rice, P., I. Longden, and A. Bleasby.2000. EMBOSS: the European Molec-ular Biology Open Software Suite. Trends Genet.16:276–277.
32.Schiemann, D. A.1995. An unstable small-colony variant of a noninvasive mutant of SalmonellaTyphimurium is highly invasive for MDCK cells. FEMS Microbiol. Lett.130:45–49.
33.Steentjes, A., K. T. Veldman, D. J. Mevius, and W. J. Landman.2002. Molecular epidemiology of unilateral amyloid arthropathy in broiler breeders associated withEnterococcus faecalis. Avian Pathol.31:31–39.
34.Wages, D. P.1998. Streptococcosis, p. 58–60.InD. E. Swayne, J. R. Glisson, M. W. Jackwood, J. E. Pearson, and W. M. Reed (ed.), A laboratory manual for the isolation and identification of avian pathogens. The American Asso-ciation of Avian Pathologists, Kennett Square, PA.
35.Zekarias, B., W. J. Landman, P. C. Tooten, and E. Gruys.2000. Leukocyte responses in two breeds of layer chicken that differ in susceptibility to induced amyloid arthropathy. Vet. Immunol. Immunopathol.77:55–69.