Copyright © 2002, American Society for Microbiology. All Rights Reserved.
Development and Application of a New Scheme for Typing
Campylobacter jejuni
and
Campylobacter coli
by PCR-Based
Restriction Fragment Length Polymorphism Analysis
Feng Shi,
1* Yuen Yuen Chen,
1Trudy M. Wassenaar,
2Walter H. Woods,
3Peter J. Coloe,
1and
Benjamin N. Fry
1Department of Biotechnology and Environmental Biology, Royal Melbourne Institute of Technology University, Melbourne, Victoria 3083,1and School of Pharmacy and Medical Sciences, University of South Australia, Adelaide, South Australia, 5001,3Australia,
and Molecular Microbiology and Genomics Consultants, Zotzenheim, Germany2
Received 28 November 2001/Returned for modification 10 January 2002/Accepted 3 March 2002
A molecular typing approach forCampylobacter jejuniandCampylobacter coliwas developed with restriction fragment length polymorphism analysis of a 9.6-kb PCR-amplified portion of the lipopolysaccharide gene cluster. Sixty-one Penner serotype reference strains were analyzed with this new genotyping scheme, and 32 genogroups were found. Eleven additional genogroups were obtained from 87 clinicalC.jejunistrains tested. This molecular typing method shows a correlation with the Penner heat-stable serotyping method, a phenotypic typing method based on lipopolysaccharide structures that is often used as a “gold standard” for subtyping
Campylobacter spp. This strong correlation suggests that the data obtained can be directly compared with
epidemiological data collected in the past by classical serotyping ofC. jejuniandC. coli.In contrast to the high percentage of nontypeability by phenotyping, this molecular typing method results in 100% typeability and provides a superior alternative to serotyping.
Campylobacter jejuniandCampylobacter coliare recognized as two of the most common causes of food-borne bacterial gastroenteritis. Furthermore,C.jejunihas been implicated as a frequent antecedent to the development of the neurologic dis-eases Guillain-Barré syndrome (GBS) (20) and Miller Fisher syndrome (37).
Numerous subtyping methods have been developed to dif-ferentiateCampylobacterstrains for epidemiologic purposes in the past two decades. More than 30 current typing methods have been reviewed elsewhere (27, 28, 29, 38). The various typing systems can be placed in two categories: phenotypic methods, which are based on expressed features such as so-matic antigens or enzyso-matic activity, and genotypic methods, which are based on specific molecular features of chromosomal or plasmid DNA.
Two serotyping schemes have been used exclusively for phe-notypic typing in the past, the scheme developed by Penner and Hennessy, which detects heat-stable (HS) antigens (31), and the one developed by Lior et al., which detects heat-labile antigens (16). The former is the most widely accepted and well evaluated phenotypic method. The molecular basis for the HS antigenic diversity inC. jejuniandC. coliis the expression of somatic (O) lipopolysaccharide (LPS) (17, 18, 21, 22, 33, 34, 35). LPS is a major constituent of the outer membrane in gram-negative bacteria and comprises three covalently linked regions: lipid A, core oligosaccharide (inner core and outer
core), and O polysaccharide. The variability of the
Campy-lobacter LPS outer core and O polysaccharide is thought to
contribute to the antigenic basis of the Penner serotyping sys-tem. Serotyping methods like these are time consuming and technically demanding, and antisera are costly to produce, which limits the use of these typing systems to specialized diagnostic laboratories. Furthermore, phenotypes can be un-stable, resulting in nonreproducible results or nontypeable strains as well as antiserum cross-reactivity, which hampers the interpretation. Genotyping methods are independent of ex-pressed features and are therefore a better alternative for typing. Several genotyping methods have recently been devel-oped, such as pulsed-field gel electrophoresis (11, 12, 40), amplified fragment length polymorphism (5, 15), flagella gene PCR-restriction fragment length polymorphism (PCR-RFLP) (1, 4, 23, 24, 26), ribotyping (6, 25, 36), and random amplified polymorphic DNA analysis (7, 10, 30). These genotyping sys-tems are more generally available and applicable than pheno-typic methods. However, most of these techniques still have their own drawbacks, such as less discriminatory power, poor reproducibility, and complex methodology. The preferred method in terms of handling, costs, and time, is RFLP analysis of PCR products. Such a method has been described for the flagellin genes (23, 26). However, when this method was ap-plied, no correlation could be detected between flagellin ge-notypes and HS serotypes (23). This greatly reduces the appli-cation of flagellin genotyping in long-term epidemiological studies. More importantly, none of these methods correlate well with the serotyping scheme used in past decades, so his-torical epidemiological trends cannot be determined. For these reasons, genotypic subtyping methods have not been widely used in epidemiological practice and remain to be developed and improved.
The LPS biosynthesis gene cluster of C. jejuni 81116 has recently been characterized in our laboratory (9). In this study,
* Corresponding author. Mailing address: Dept. of Biotechnol. and Environ. Biol., Royal Melbourne Inst. of Technol. Univ., Bundoora W. Campus, Bldg. 223, Lvl. 1, Plenty Rd., Bundoora 3083 VIC, Mel-bourne, Australia. Phone: 61-3-992-57132. Fax: 61-3-992-57110. E-mail: [email protected].
1791
on May 15, 2020 by guest
http://jcm.asm.org/
a new genotyping scheme, the LG (LPS genes) genotyping system, which is based on PCR-RFLP of this gene cluster, has been established. The application value of this new system was also evaluated by typing the reference Penner serotype strains and a number ofC. jejuni and C. coli clinical isolates. This typing scheme is the first genotyping scheme to our knowledge that has the same molecular basis as the Penner serotyping scheme.
MATERIALS AND METHODS
Bacterial strains and growth conditions.The strains used in this study are listed in Table 1.
Cultures were stored at⫺70°C in heart infusion broth (Difco) containing 50% glycerol. Bacteria were grown on Columbia agar (Oxoid) supplemented with 5% (vol/vol) defibrinated horse blood andCampylobacterselective supplement (Skir-row) (Oxoid) for 48 h at 42°C under microaerophilic conditions (CO2, 10.5%;
O2, 5%; balance, N2).
Chromosomal DNA preparation.DNA was isolated from pure cultures by the cetyltrimethylammonium procedure (3). Briefly, one lawn plate ofCampylobacter was grown overnight, harvested in 9.4 ml of Tris-EDTA buffer and 0.1 ml of 0.5 M EDTA, and lysed with 0.5 ml of 10% (wt/vol) sodium dodecyl sulfate. Pro-teinase K was added to a final concentration of 0.1 mg/ml, and the mixture was incubated at 37°C for 2 h. Then 1.8 ml of 5 M NaCl and 1.5 ml of 10% (wt/vol) cetyltrimethylammonium in 0.7 M NaCl were added, and the mixture was incu-bated for 30 min at 65°C. After the addition of 5 ml of 24:1 chloroform-isoamyl alcohol, the mixture was centrifuged for 10 min at 4°C. The aqueous layer was removed, and 5 ml of 25:24:1 phenol-chloroform-isoamyl alcohol was added. The aqueous layer was removed after centrifugation. The DNA was precipitated with a 0.6-volume of isopropanol and dissolved in water. The concentration of the DNA was measured by spectrophotometric absorbance at 260 nm.
Gene distribution analysis.The gene distribution of thewlacluster was ana-lyzed by PCR. Primer sets used for amplification of the individual genes in the wlacluster are given in Table 2. PCR was performed in a 50-l reaction volume with a Perkin Elmer GeneAMP 2400 thermal cycler. The reaction mixtures consisted of 1⫻reaction buffer with 2.0 mM MgSO4(Promega), 2.5 U ofTaq
DNA polymerase (Promega), 100 ng of forward and reverse primer, 0.2 mM concentrations of deoxynucleoside triphosphates, and 100 ng of template DNA. The reaction included an initial denaturation of DNA at 94°C for 1 min and then 35 cycles of consecutive denaturation (30 s, 94°C), primer annealing (30 s, 60°C), and chain extension (based on a rate of 1 kb/minute, 72°C). A final elongation step was performed for 10 min at 72°C.
Penner serotyping.Penner serotypes ofCampylobacterspp. were detected by using the passive hemagglutination technique described by Penner and Hennessy (31), with a heated supernatant from the bacterial culture as the antigen. Anti-sera were prepared by injecting rabbits intravenously with saline bacterial sus-pensions ofCampylobacterserostrains. Some antisera were absorbed with
cross-reacting HS reference strains to clarify the serotyping results; in particular, antiserum prepared against HS-4 was absorbed with HS-16 bacteria to remove minor reactions with HS-13 and HS-16.
PCR-RFLP analysis.The primers galE1 and wlaH3 (Table 2) were used to amplify a 9.6-kb fragment. PCRs were performed as stated above except that a MgSO4concentration of 2.0 mM was used with 2.5 U ofPfuDNA polymerase
(Promega) instead ofTaqand a chain extension of 15 min at 72°C per cycle. After amplification, 10l of PCR product was digested with 10 U of restriction enzymes (HhaI andNlaIII [New England Biolabs] andDdeI andHindIII [Pro-mega]) in a total volume of 20l with 2g of bovine serum albumin for more than 3 h at 37°C. The digest was analyzed by electrophoresis by using a 1.5% agarose gel and stained with ethidium bromide. Lambda DNA digested withPstI was used as a reference size marker.
RESULTS
Amplification analysis of LPS gene cluster.All 12 genes in the wla cluster were tested for their distribution among se-lected reference serotypeC. jejunistrains. In order to detect duplications, gene order rearrangements, or deletions, PCR amplification was carried out for each single gene as well as across two genes that were found adjacent in thewlacluster of
[image:2.587.42.282.83.267.2]C. jejuni81116. Primers used in the distribution analysis were designed from the published sequence of the LPS gene cluster
TABLE 1. Bacterial strains used in this study
Organism and strain(s) Source(s) or reference No. ofstrains
E. coliO111 9 1
S. entericaserovar
Typhi-murium Melbourne, Australia, human 1
C. lari Melbourne, Australia, human 1 C. hyoilei Melbourne, Australia, human 1 C. hyointestinalis Melbourne, Australia, human 1 C. sputorum Melbourne, Australia, human 1 C. fetus Melbourne, Australia, human 1 C. upsaliensis Melbourne, Australia, human 1 Campylobacter concisus Melbourne, Australia, human 1 C. jejuni81116 United Kingdom, human 1 C. jejuniclinical strains Adelaide, Australia, human 67 C. jejuniclinical strains Canberra, Australia, human 6 C. jejuniclinical strains Sydney, Australia, human GBS 1 C. jejuniclinical O:19 strains China, Mexico, and Japan, human 6 C. jejuniclinical strains South Africa, human 6 C. jejunistrains South Africa, chicken and ostrich 1
Penner serotype strains 32 61
TABLE 2. Primers used for gene distribution analysis and PCR-RFLP
Target gene (amplicon size [bp]) Primer
a Positionb Sequence
galE(911) galE1 1185–1206 GCGGTGGTGCAGGTTATATAGG galE2 2077–9096 TAGGCTGCCAAGAAGTAAGA
wlaB(1,419) wlaB1 2372–2391 TTGGAGTGGGGCTTATTGTT wlaB2 3772–3791 TGCGTTATCGTAGAAAGGCG
wlaC(599) wlaC1 4167–4186 ACTCCACTCATCATTAGCGA wlaC2 4747–4766 TTCATCATCACAACCTACCA
wlaD(396) wlaD1 5146–5165 TTAGATCAAGCAAGTGGCGA wlaD2 5521–5542 CTAAAGTCACGCTATCTTGCCT
wlaE(969) wlaE1 5858–5877 TTTAGGAAGTGGTGGTGCTG wlaE2 6808–6827 ATCGTCTTCAAGCATAGTCT
wlaF(1,163) wlaF1 7176–7195 TCCGCGCTTACTTATTGGCT wlaF2 8318–8339 CACATCGCTATAATAACGCACA
wlaG(824) wlaG1 9186–9205 CCGCAAGATGAATACACGCA wlaG2 9991–10010 AGCCTCTACACAACCTTCAC
wlaH(232) wlaH1 10388–10407 TGAGCGATGAAAGAGATGAG wlaH2 10601–10620 CGCCCATCCTGTTATACCTG wlaH3 10783–10806 TCAGTTCTTGCCATTAAATTTCTC
wlaI(245) wlaI1 10820–10839 TATGGTGCTAGTGGTCATGG wlaI2 11046–11065 CACGCTTGCACTAGGACTTA
wlaK(631) wlaK1 11868–11887 CATCTTTATGGCAATGCGGC wlaK2 12478–12499 ACTTCTTGAGTGTGCATAGCTT
wlaL(569) wlaL1 13312–13331 GAAGAACTTGTAGCTTATGG wlaL2 13862–13881 CTCACAAACTCTCTTAGTGC
wlaM(738) wlaM1 14507–14526 ACTTCCTGATATTCACCTTG wlaM2 15226–15245 GCTCACTCCACCGATAAGAT
aAll the primers were designed within the genes.
bNucleotide positions are according to the National Center for Biotechnology Information published sequence number Y11648.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.587.300.542.353.702.2]fromC. jejuni81116 (9) and listed in Table 2. The individual genes were amplified by primers designed within the gene; for example, thegalEgene was analyzed by the primer pair galE1-galE2. Two adjacent genes were analyzed by a primer combi-nation of the upstream primer of the upstream gene and the downstream primer of the downstream gene; for example, the
adjacent genesgalE and wlaB were amplified by the primer
pair galE1-wlaB2. All of the 12 genes were detected in the 12
C. jejunistrains investigated (data not shown), which indicated
that the genes in the wla cluster were conserved among C.
jejuni serotype reference strains. Of the 12 strains tested, 7 strains gave a PCR fragment of 2.3 kb with primers wlaI1 and wlaK2, which was compared to the 1.7-kb fragment obtained for the other strains (data not shown). This indicated that an extra fragment, possibly carrying a gene, is present between the
wlaIand thewlaKgenes in these strains. A gene namedwlaJ
was described at this position by Wood et al. (39) in 54% ofC. jejuni isolates. The highly conserved nature of thewla gene cluster inC. jejunimakes it a suitable target for the develop-ment of a gene typing system.
Of the 12 genes in thewlacluster, thegalEandwlaHgenes are most similar to genes found in other gram-negative bacte-ria, with⬎52 and⬎53% amino acid similarity, respectively (9). Therefore, it was reasoned that these genes would be the most
conserved withinC. jejunistrains. Sequence alignment of the
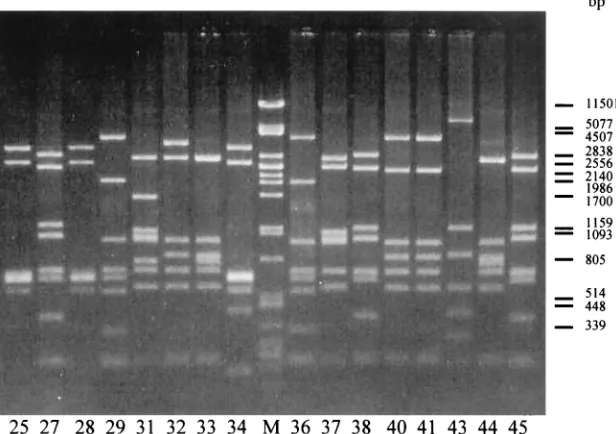
galEgene from threeC. jejunistrains showed that, indeed, the
galEgene is very conserved amongC. jejunistrains (Fig. 1a). Also, thewlaHgene was found to be conserved when aligning this gene with fiveC. jejuni strains (Fig. 1b). The most con-served regions within thegalEand thewlaHgenes were used to design the primers galE1 and wlaH3 to amplify part of the LPS gene cluster (Fig. 1). These two primers gave a 9.6-kb product when used in a PCR, which spans eight genes in thewlacluster. All 61 reference Penner serotypeCampylobacterstrains were tested and gave the expected products (data not shown).
The use of strongly conserved genes increases the risk that other species would be detected by the PCR. Therefore, we tested the cross-reactivity of the PCR primers on other bacte-ria within or outside theCampylobactergenus. No PCR prod-ucts could be obtained from the DNA ofCampylobacter lari,
Campylobacter hyoilei, Campylobacter hyointestinalis, Campy-lobacter sputorum,Campylobacter fetus, andCampylobacter up-saliensisor from the DNA of the enterobacteria Escherichia coliandSalmonella entericaserovar Typhimurium. The result remained negative when the annealing temperature was low-ered to 58°C (data not shown). This indicates that this typing scheme is specific forC. jejuniandC. coli.
FIG. 1. (a) Sequence alignment analysis of part of thegalEgene fromC. jejuni81116 (HS-6) (9),C. jejuni11168gp (HS-2) (Sanger Centre website [ftp://ftp.sanger.ac.uk/pub/pathogens/cj/]), andC. jejuniHB9313 (HS-19) (unpublished data). (b) Sequence alignment analysis of part of thewlaHgene fromC. jejuni81116 (HS-6),C. jejuni11351 (HS-23) (39),C. jejuni11168gp (HS-2),C. jejuni11168db (HS-2) (39), andC. jejuni
HB9313 (HS-19). The bold characters indicate the primer sequences. The arrows indicate the orientation of the primers.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:3.587.55.538.76.399.2]RFLP within the amplified PCR product.The presence and degree of conservation of restriction enzyme recognition sites was evaluated for individualwlagenes in a pilot study (data not shown). As a result of this inventory, the enzymes HindIII,
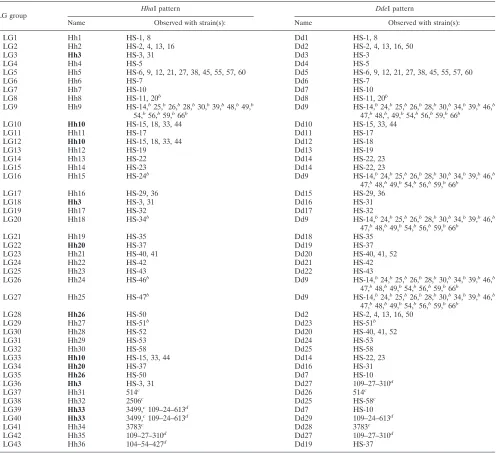
HhaI, DdeI, and NlaIII were chosen to digest the obtained 9.6-kb amplified amplicon from all Penner serotype reference strains. This resulted in RFLP profiles of which typical exam-ples are shown in Fig. 2. Eleven distinctHindIII patterns, 30
HhaI patterns, 25DdeI patterns, and 19NlaIII patterns were
detected. The combination of the HhaI and DdeI patterns
resulted in 32 separate genogroups (LG1 to LG32) for the 61 reference Penner serotype strains, and this enzyme combina-tion was the method of choice for the LG genotyping scheme (Table 3). In most cases, theHhaI patterns will decide which
LG genogroup an unknown strain belongs to.DdeI patterns
need only be tested if theHhaI pattern is not unique, such as for the patterns Hh3, 10, 20, 26, and 33. All obtained profiles are available at the website http://www.bh.rmit.edu.au/abbt /campylobacter/typing.html.
This new typing scheme was then tested on 87 clinical C. jejunistrains, among which 80 strains were HS serotyped. This resulted in an additional 11 new LG genogroups, LG33 to LG43 (Table 3).
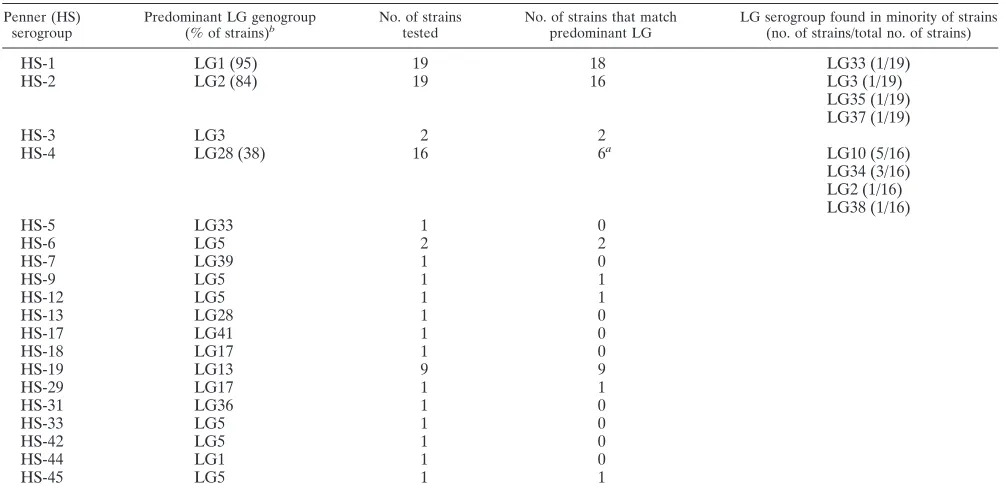
The results of the LG genotyping of the 80 HS serotyped clinicalC. jejunistrains were compared with the Penner sero-typing (Table 4). When two or more strains from the same Penner serogroup were typed, there was a correlation with the LG system, except for HS-4.
DISCUSSION
As for other infectious agents, subtyping methods are re-quired forCampylobacterspp. to recognize temporal and re-gional trends and to identify and recognize pathotypes. The most common phenotypic subtyping scheme forC. jejuniis the
Penner serotyping system. This method was recently modified by Frost et al. (8) to improve the method and to make it less technically demanding. Despite this improvement, serotyping is still time consuming and costly because of the amount and numbers of the sera needed to be produced, maintained, and quality controlled. Although a number of genotyping methods have recently been established and are regarded as more ad-vantageous than phenotypic methods due to better reliability, higher discriminatory power, better typeability, and general accessibility, serotyping methods still could not be replaced completely as they have been widely used for many years. The serological heterogeneity ofC. jejunistrains is determined by variation in LPS structures for which the most likely molecular basis is DNA polymorphism in the genes encoding the LPS biosynthetic enzymes. Therefore, a molecular typing method has been developed based on thewlalocus which combines the advantage of a simple and fast PCR-RFLP-based technique with a high typeability and a strong correlation to the HS
serotyping scheme used in the past. The wla gene cluster,
involved in LPS biosynthesis, is highly conserved in Campy-lobacterstrains, and within this cluster, thegalEand thewlaH
genes show a high degree of similarity to genes in other gram-negative bacteria (9). Therefore, these genes are likely to be highly conserved among different C. jejuni strains and were used to design primers to amplify the intragenic region.
Our results show that the method of LG genotyping is spe-cific forC. jejuniandC. coliand has a higher discriminatory power than the Penner serotyping method (Simpson’s index is 0.872 for the LG typing method and 0.842 for the Penner typing method). Sixty-one reference strains representing dif-ferent Penner serotypes ofC. jejuniandC. coliand 87 clinical
C. jejuni strains were included for LG analysis. All tested strains yielded a PCR product of consistent size (9.6 kb) that could be digested by the restriction enzymesHhaI andDdeI to achieve 100% typeability. The RFLP patterns have a
reason-FIG. 2. PCR-RFLP patterns ofHhaI from part of reference HS serotype strains. Lane numbers in the figure indicateC. jejunireference HS serotype strain numbers. Lane M,DNA digested withPstI.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.587.137.445.78.295.2]able size range which can be separated by a 1.5% midi agarose gel. The patterns are very easy to read and can be digitized for comparison between laboratories.
A total of 43 LG profile types were identified among the 61 reference HS serostrains ofC. jejuni and C. coliand the 87 clinicalC. jejunistrains. A comparison of the LG typing data of 80C. jejunistrains with the Penner serotyping data showed, as expected, a strong association between these two typing meth-ods. All strains belonging to Penner groups 3, 6, 9, 12, 19, 29, 31, and 45 shared their LG banding patterns with the respec-tive Penner reference strains (Table 4). Notably, Penner group 19, which has been recognized to have a strong association with
[image:5.587.46.542.83.536.2]GBS, is recognized as a separate LG genotype with conserved banding patterns for the 9 clinical isolates tested, originating from different sources. Strains belonging to the Penner groups 1 and 2 had identical LG patterns for 95 and 84% of the investigated strains, respectively. In contrast, the predominant LG genotype of strains belonging to the HS-4 complex sero-group was only found in 38% of the strains tested. Penner serogroups 1, 2, 3, and especially 4, which together comprise a large proportion of clinical isolates in certain parts of the world, are not clonal but represent a genetically diverse pop-ulation (2, 13, 14, 19, 27, 32). In this respect, the strong cor-relation between LG genotype and HS-1, HS-2, and HS-3
TABLE 3. LG genotyping systema
LG group HhaI pattern DdeI pattern
Name Observed with strain(s): Name Observed with strain(s):
LG1 Hh1 HS-1, 8 Dd1 HS-1, 8
LG2 Hh2 HS-2, 4, 13, 16 Dd2 HS-2, 4, 13, 16, 50
LG3 Hh3 HS-3, 31 Dd3 HS-3
LG4 Hh4 HS-5 Dd4 HS-5
LG5 Hh5 HS-6, 9, 12, 21, 27, 38, 45, 55, 57, 60 Dd5 HS-6, 9, 12, 21, 27, 38, 45, 55, 57, 60
LG6 Hh6 HS-7 Dd6 HS-7
LG7 Hh7 HS-10 Dd7 HS-10
LG8 Hh8 HS-11, 20b Dd8 HS-11, 20b LG9 Hh9 HS-14,b25,b26,b28,b30,b39,b48,b49,b
54,b56,b59,b66b Dd9 HS-14,
b24,b25,b26,b28,b30,b34,b39,b46,b 47,b48,b, 49,b54,b56,b59,b66b
LG10 Hh10 HS-15, 18, 33, 44 Dd10 HS-15, 33, 44
LG11 Hh11 HS-17 Dd11 HS-17
LG12 Hh10 HS-15, 18, 33, 44 Dd12 HS-18
LG13 Hh12 HS-19 Dd13 HS-19
LG14 Hh13 HS-22 Dd14 HS-22, 23
LG15 Hh14 HS-23 Dd14 HS-22, 23
LG16 Hh15 HS-24b Dd9 HS-14,b24,b25,b26,b28,b30,b34,b39,b46,b 47,b48,b49,b54,b56,b59,b66b
LG17 Hh16 HS-29, 36 Dd15 HS-29, 36
LG18 Hh3 HS-3, 31 Dd16 HS-31
LG19 Hh17 HS-32 Dd17 HS-32
LG20 Hh18 HS-34b Dd9 HS-14,b24,b25,b26,b28,b30,b34,b39,b46,b 47,b48,b49,b54,b56,b59,b66b
LG21 Hh19 HS-35 Dd18 HS-35
LG22 Hh20 HS-37 Dd19 HS-37
LG23 Hh21 HS-40, 41 Dd20 HS-40, 41, 52
LG24 Hh22 HS-42 Dd21 HS-42
LG25 Hh23 HS-43 Dd22 HS-43
LG26 Hh24 HS-46b Dd9 HS-14,b24,b25,b26,b28,b30,b34,b39,b46,b 47,b48,b49,b54,b56,b59,b66b
LG27 Hh25 HS-47b Dd9 HS-14,b24,b25,b26,b28,b30,b34,b39,b46,b 47,b48,b49,b54,b56,b59,b66b
LG28 Hh26 HS-50 Dd2 HS-2, 4, 13, 16, 50
LG29 Hh27 HS-51b Dd23 HS-51b
LG30 Hh28 HS-52 Dd20 HS-40, 41, 52
LG31 Hh29 HS-53 Dd24 HS-53
LG32 Hh30 HS-58 Dd25 HS-58
LG33 Hh10 HS-15, 33, 44 Dd14 HS-22, 23
LG34 Hh20 HS-37 Dd16 HS-31
LG35 Hh26 HS-50 Dd7 HS-10
LG36 Hh3 HS-3, 31 Dd27 109–27–310d
LG37 Hh31 514c Dd26 514c
LG38 Hh32 2506c Dd25 HS-58c
LG39 Hh33 3499,c109–24–613d Dd7 HS-10 LG40 Hh33 3499,c109–24–613d Dd29 109–24–613d
LG41 Hh34 3783c Dd28 3783c
LG42 Hh35 109–27–310d Dd27 109–27–310d LG43 Hh36 104–54–427d Dd19 HS-37
aThe LG groups with their denominatedHhaI andDdeI patterns. Each pattern name is followed by a list of HS strains in which the pattern is observed. Bold Hh pattern names indicate nonunique patterns for which determination of theDdeI pattern is compulsive.
bC. colistrain.
cClinical isolate from Adelaide, Australia. dClinical isolate from Canberra, Australia.
on May 15, 2020 by guest
http://jcm.asm.org/
serotype is remarkable. The strains from the HS-4 group more often belonged to genotypes LG10, 28, or 34, than to LG2, which is the LG genotype for the HS-4 reference serotype strain. This indicates that the serotyping reference strain does not have the most common DNA polymorphism for this ge-netically diverse serogroup. In conclusion, we believe that this fast and simple method is a suitable alternative to serotyping as an application for typing clinical and food isolates ofC. jejuni
and as a method for facilitating epidemiological research of
Campylobacterspp.
The added value of the proposed LG typing scheme to the genotyping schemes already existing forC. jejuniis the strong correlation to the classical HS serotyping scheme, which allows comparison with historical data. In addition, LG typing allows for the classification of strains that are nontypeable by sero-typing, since such strains are likely to be LG typeable. The observed discriminatory power of LG is slightly higher than that of serotyping, especially for serotype HS-4. In conclusion, we believe that this fast and simple method is a suitable alter-native to serotyping and a valuable addition to the genotyping methods available for typing clinical and food isolates ofC. jejuni and facilitating epidemiological research of Campy-lobacterspp.
ACKNOWLEDGMENTS
We thank Irving Nachamkin for providing some of the clinicalC. jejunistrains for this study. We also gratefully acknowledge the coop-eration of diagnostic laboratories throughout South Australia in pro-viding isolates for typing.
REFERENCES
1.Alm, R., P. Guerry, and T. J. Trust.1993. Distribution and polymorphism of the flagellin genes from isolates ofCampylobacter coliandCampylobacter jejuni. J. Bacteriol.175:3051–3057.
2.Asrat, D. A., A. Hathaway, E. S. Sjögren, E. Ekwall, and B. Kaijser.1997. The serotype distribution ofCampylobacter jejuniandCampylobacter coli isolated from patients with diarrhea and controls in Tikur Anbassa Hospital, Addis Ababa, Ethiopia. Epidemiol. Infect.118:222–226.
3.Ausubel, F. M., R. Brent, R. E. Kingston, D. M. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.).1995. Current protocols in molecular biology. John Wiley & Sons, New York, N.Y.
4.Ayling, R. D., M. J. Woodward, S. Evans, and D. G. Newell.1996. Restriction fragment length polymorphism of polymerase chain reaction products ap-plied to the differentiation of poultry campylobacters for epidemiological investigations. Res. Vet. Sci.60:168–172.
5.Duim, B., T. M. Wassenaar, A. Rigter, and J. Wagenaar.1999. High-reso-lution genotyping ofCampylobacterstrains isolated from poultry and humans with amplified fragment length polymorphism fingerprinting. Appl. Environ. Microbiol.65:2369–2375.
6.Fayos, A., R. J. Owen, M. Desai, and J. Hernandez.1992. Ribosomal RNA gene restriction fragment diversity amongst Lior biotypes and Penner sero-types ofCampylobacter jejuniandCampylobacter coli. FEMS Microbiol. Lett. 95:87–94.
7.Fayos, A., R. J. Owen, J. Hernandez, C. Jones, and A. Lastovica.1993. Molecular subtyping by genome and plasmid analysis ofCampylobacter jejuni serogroups O1 and O2 (Penner) from sporadic and outbreak cases of human diarrhoea. Epidemiol. Infect.111:415–427.
8.Frost, J. A., A. N. Oza, R. T. Thwaites, and B. Rowe.1998. Serotyping scheme forCampylobacter jejuniandCampylobacter colibased on direct agglutina-tion of heat-stable antigen. J. Clin. Microbiol.36:335–339.
9.Fry, B. N., V. Korolik, J. A. ten Brinke, M. T. T. Pennings, R. Zalm, B. J. J. Teunis, P. J. Coloe and B. A. M. van der Zeijst.1998. The lipopolysaccharide biosynthesis locus ofCampylobacter jejuni81116. Microbiol.144:2049–2061. 10.Fujimoto, S., B. M. Allos, N. Misawa, C. M. Patton, and M. J. Blaser.1997. Restriction fragment length polymorphism analysis and random amplified polymorphic DNA analysis of Campylobacter jejunistrains isolated from patients with Guillain-Barré syndrome. J. Infect. Dis.176:1105–1108. 11.Gibson, J. R., C. Fitzgerland, and J. Owen.1995. Comparison of PFGE,
ribotyping and phage-typing in the epidemiological analysis of Campy-lobacter jejuniserotype HS2 infections. Epidemiol. Infect.115:215–225. 12.Imai, Y., M. Kikuchi, M. Matsuda, M. Honda, M. Fukuyama, M. Tsukada,
and C. Kaneuchi.1994. Macro-fingerprinting analysis at the chromosomal genomic DNA level of isolates of thermophilic Campylobacter coliand Campylobacter jejuni, by pulsed-field gel electrophoresis. Cytobios78:115– 122.
[image:6.587.42.545.83.328.2]13.Jones, D. M., J. D. Abbott, M. J. Painter, and E. M. Sutcliffe.1984. A comparison of biotypes and serotypes ofCampylobactersp. isolated from patients with enteritis and from animal and environmental sources. J. Infect. 9:51–58.
TABLE 4. Comparison of LG and HS typing methods Penner (HS)
serogroup Predominant LG genogroup(% of strains)b No. of strainstested No. of strains that matchpredominant LG LG serogroup found in minority of strains(no. of strains/total no. of strains)
HS-1 LG1 (95) 19 18 LG33 (1/19)
HS-2 LG2 (84) 19 16 LG3 (1/19)
LG35 (1/19) LG37 (1/19)
HS-3 LG3 2 2
HS-4 LG28 (38) 16 6a LG10 (5/16)
LG34 (3/16) LG2 (1/16) LG38 (1/16)
HS-5 LG33 1 0
HS-6 LG5 2 2
HS-7 LG39 1 0
HS-9 LG5 1 1
HS-12 LG5 1 1
HS-13 LG28 1 0
HS-17 LG41 1 0
HS-18 LG17 1 0
HS-19 LG13 9 9
HS-29 LG17 1 1
HS-31 LG36 1 0
HS-33 LG5 1 0
HS-42 LG5 1 0
HS-44 LG1 1 0
HS-45 LG5 1 1
aOnly for HS-4 does the predominant LG genogroup differ from that found with the serotype reference strain. bPercentage given only if not 100 or 0%.
on May 15, 2020 by guest
http://jcm.asm.org/
14.Karmali, M. A., J. L. Penner, P. C. Fleming, A. Williams, and J. N. Hen-nessy.1983. The serotype and biotype distribution of clinical isolates of Campylobacter jejuniandCampylobacter coliover a three-year period. J. In-fect. Dis.147:243–246.
15.Kokotovic, B., and S. L. W. On.1999. High-resolution genomic fingerprinting of Campylobacter jejuniandCampylobacter coli by analysis of amplified fragment length polymorphisms. FEMS Microbiol. Lett.173:77–84. 16.Lior, H., D. L. Woodward, J. A. Edgar, L. J. Laroche, and P. Gill.1982.
Serotyping ofCampylobacter jejuni by slide agglutination based on heat-labile antigenic factors. J. Clin. Microbiol.15:761–768.
17.Mandatori, R., and J. L. Penner.1989. Structural and antigenic properties of Campylobacter coli. Infect. Immun.57:3506–3511.
18.Mills, S. D., W. C. Bradbury, and J. L. Penner.1985. Basis for serological heterogeneity of thermostable antigens ofCampylobacter jejuni. Infect. Im-mun.50:284–291.
19.Mills, S. D., G. O. Aspinall, A. G. McDonald, T. S. Raju, L. A. Kurjanczyk, and J. L. Penner.1992. Lipopolysaccharide antigens ofCampylobacter jejuni, p. 223–229. InI. Nachamkin, M. J. Blaser, and L. S. Tompkins. (ed.), Campylobacter jejuni: current status and future trends. American Society for Microbiology, Washington, D.C.
20.Mishu, B., and M. J. Blaser.1993. Role of infection due toCampylobacter jejuniin the initiation of Guillain-Barré syndrome. Clin. Infect. Dis.17:104– 108.
21.Moran, A. P., and T. U. Kosunen.1989. Serological analysis of the heat-stable antigens involved in serotyping Campylobacter jejuniand Campy-lobacter coli. APMIS97:253–260.
22.Moran, A. P., and J. L. Penner.1999. Serotyping ofCampylobacter jejuni based on heat-stable antigens: relevance, molecular basis and implications in pathogenesis. J. Appl. Microbiol.86:361–377.
23.Nachamkin, I., K. Bohachick, and C. M. Patton.1993. Flagellin gene typing ofCampylobacter jejuniby restriction fragment length polymorphism analy-sis. J. Clin. Microbiol.31:1531–1536.
24.Nachamkin, I., H. Ung, and C. M. Patton.1996. Analysis of HL and O serotypes ofCampylobacterstrains by the flagellin gene typing system. J. Clin. Microbiol.34:277–281.
25.Owen, R. J., M. Desai, and S. Garcia.1993. Molecular typing of thermotol-erant species ofCampylobacterwith ribosomal RNA gene patterns. Res. Microbiol.144:709–720.
26.Owen, R. J., A. Fayos, J. Hernandez, and A. Lastovica.1993. PCR-based restriction fragment length polymorphism analysis of DNA sequence diver-sity of flagellin genes ofCampylobacter jejuniand allied species. Mol. Cell. Probes7:471–480.
27.Owen, R. J., and J. R. Gibson.1995. Update on epidemiological typing of Campylobacter. PHLS Microbiol. Dig.12:2–6.
28.Patton, C. M., I. K. Wachsmuth, G. M. Evins, J. A. Kiehlbauch, B. D. Plikaytis, N. Troup, L. Tompkins, and H. Lior. 1991. Evaluation of 10 methods to distinguish epidemic-associatedCampylobacterstrains. J. Clin. Microbiol.29:680–688.
29.Patton, C. M., and I. K. Wachsmuth.1992. Typing schemes—are current methods useful?, p. 110–128. InI. Nachamkin, M. J. Blaser, and L. S. Tompkins (ed.), Campylobacter jejuni: current status and future trends. American Society for Microbiology, Washington, D.C.
30.Payne, R. E., M. D. Lee, D. W. Dreesen, and H. M. Barnhart.1999. Molec-ular epidemiology ofCampylobacter jejuniin Broiler flocks using randomly amplified polymorphic DNA-PCR and 23s rRNA-PCR and role of litter in its transmission. Appl. Environ. Microbiol.65:260–263.
31.Penner, J. L., N. Hennessy.1980. Passive hemagglutination technique for serotypingCampylobacter fetussubsp.jejunion the basis of soluble heat-stable antigens. J. Clin. Microbiol.12:732–737.
32.Penner, J. L., J. N. Hennessy, and R. V. Congi.1983. Serotyping of Campy-lobacter jejuniandCampylobacter colion basis of thermostable antigens. Eur. J. Clin. Microbiol.2:378–383.
33.Perez Perez, G. I., and M. J. Blaser.1985. Lipopolysaccharide characteristics of pathogenic campylobacters. Infect. Immun.47:353–359.
34.Perez Perez, G. I., J. A. Hopkins, and M. J. Blaser.1985. Antigenic heter-ogeneity of lipopolysaccharides from Campylobacter jejuni and Campy-lobacter fetus. Infect. Immun.48:528–533.
35.Preston, M. A., and J. L. Penner.1987. Structural and antigenic properties of lipopolysaccharides from serotype reference strains ofCampylobacter jejuni. Infect. Immun.55:1806–1812.
36.Russell, R. G., J. A. Kiehlbauch, J. I. Sarmiento, P. Panigrahi, D. C. Blake, and R. Haberbager. 1994. Ribosomal RNA patterns identify additional strains ofCampylobacter jejuniand C. coliamong isolates serotyped by heat-stable and heat-labile antigens. Lab. Anim. Sci.44:579–583. 37.Salloway, S., L. A. Mermel, M. Seamans, G. O. Aspinall, J. E. Nam Shin,
L. A. Kurjanczyk, and J. L. Penner.1996. Miller-Fisher syndrome associated withCampylobacter jejunibearing lipopolysaccharide molecules that mimic human ganglioside GD3. Infect. Immun.64:2945–2949.
38.Wassenaar, T. M., and D. G. Newell.2000. Genotyping ofCampylobacter spp. Appl. Environ. Microbiol.66:1–9.
39.Wood, A. C., N. J. Oldfield, C. A. O’Dwyer, and J. M. Ketley.1999. Cloning, mutation and distribution of a putative lipopolysaccharide biosynthesis locus inCampylobacter jejuni. Microbiology145:379–388.
40.Yan, W., N. Chang, and D. E. Taylor.1991. Pulsed-field gel electrophoresis ofCampylobacter jejuniandCampylobacter coligenomic DNA and its epi-demiologic application. J. Infect. Dis.163:1068–1072.

![FIG. 1. (a) Sequence alignment analysis of part of the galEthewebsite [ftp://ftp.sanger.ac.uk/pub/pathogens/cj/]), and gene from C](https://thumb-us.123doks.com/thumbv2/123dok_us/8240877.830555/3.587.55.538.76.399/fig-sequence-alignment-analysis-galethewebsite-sanger-pathogens-gene.webp)