THE EFFECT OF A TEMPERATURE GRADIENT ON
THE EARLY DEVELOPMENT OF THE FROG
BY I. L. DEAN, B.Sc, M. E. SHAW, M.Sc. AND
M. A. TAZELAAR, B.Sc.
From the Zoological Laboratory, King's College, University of London. (With Two Plates and Fourteen Text-figures.)
{Received -znd February 1928.)
1. INTRODUCTION.
THE present paper is an account of experiments carried out under the direction of Professor Julian S. Huxley, in April and May, 1927, in an attempt to amplify and extend similar experiments done by the latter in the spring of 1926 (Huxley, 1926). The object of these experiments was to ascertain what effects a steep apico-basal temperature gradient has on the developing frog's egg. The present experiments were carried out on a larger scale, their range being widened to include the effects of both apico-basal and lateral temperature gradients on later embryonic stages. The apparatus was improved, making it possible to use a steeper temperature gradient and so obtain more striking results. Some of the results have been embodied in a general paper (Huxley, 1927).
The only other work of a nature at all similar was that of Vogt (1927) and early work on the chick by Dareste (1877), and Warynski and Fol (1884). The experi-ments of the early workers were carried out with the object of ascertaining the causes underlying the production of teratological embryos, the ones which are of interest in this connection being those dealing with the cause of heterotaxia in the chick. Their results showed that it was possible to obtain considerable differential growth by heating a part of the embryo and perhaps even to alter the relative proportions of the parts (Dareste, 1877).
Vogt's paper only came to our notice after our experiments were finished and the manuscript of this paper written. However, his method was somewhat different, involving the cooling of one complete half of the embryo and the heating of the other complete half, and, moreover, his experiments were carried out with the object of finding whether the "organiser" would form a complete embryo in the heated half of the original embryo. His results were similar to our results with lateral temperature gradients, but more marked as the eggs were exposed to the gradient for a much longer time (up to five days).
310 I. L. DEAN, M. E. SHAW and M. A. TAZELAAR
the effects of temperature gradients "adjuvant" (animal pole heated, vegetative cooled), "antagonistic" (animal pole cooled, vegetative heated), and lateral (one side of egg heated, the other cooled) to the egg's own apico-basal gradient were investigated. These experiments were developed with the idea that a graded stimulus applied to the developing egg should be of considerable value in testing Child's work on axial gradients and some of the conclusions he drew from it.
It is unnecessary to recapitulate in detail Child's well-known theory of Axial Gradients. A strong mass of evidence supports the view that there exists a gradient in rate of protoplasmic activity passing down the main axis of the egg from the animal to the vegetative pole. Bellamy (1919) and Bellamy and Child (1924), by the method of susceptibility to various chemicals, showed that the animal pole and dorsal lip of the blastopore in the frog's egg were regions of greatest physiological activity: while Geinitz' (19256), Spemann and Mangold's (1924) and Bautzmann's (1926) organiser grafts, Ekman's, Stohr's and Copenhaver's work on rudiments of limb buds, gills and heart, all bear evidence that there exists an antero-posterior gradient in the vertebrate. It would seem that along this axis the rudiments of the different organs and parts of the body are determined at various levels. Huxley (1924) expressed the view that during gastrulation, in addition to the gradient system a chemo-differentiation which is irreversible sets in (see also Goldschmidt, 1927), so that if either the original gradient or the process of chemo-differentiation taking place with relation to it, could be influenced by a temperature gradient (or any other means), it is reasonable to expect that the modification would be morphologically expressed in the embryo.
From the experiments of Bellamy (1921, 1922, 1924) and of Cotronei (1921, 1922) on frog's eggs, and of Stockard (1909) on fish embryos, it was supposed that both adjuvant and antagonistic gradients would have a marked effect on the relative proportions of the tadpoles developing from eggs to which these gradients had been applied. According to the axial gradient theory, the head region should be especially sensitive and one would expect stimulation of the head with adjuvant, and inhibition with antagonistic gradient, to result in micro- and mega-cephalous tadpoles.
Huxley demonstrated by his 1926 experiments that temperature gradients influenced the segmentation of the egg so that the normal relative sizes of animal and vegetative cells were markedly altered. In extreme cases where antagonistic gradient was used the animal cells (in mid-segmentation) were as large as the yolk cells. Moreover tadpoles which developed from eggs treated with tem-perature gradients were found to show an appreciable alteration in relative head size, adjuvant gradients producing slightly megacephalous and antagonistic gradients slightly microcephalous tadpoles (see Huxley, 1927).
bud and later stages, with the object of ascertaining whether the application of temperature gradients during these later phases of development, when the processes of chemo-differentiation and growth are actively taking place, has an effect on the proportions of the tadpole.
2. APPARATUS, MATERIAL, METHOD.
The apparatus (figured by Huxley 1927, p. 508) consisted of silver discs fused on to shallow copper plates supplied with inlet and outlet tubes through which, a flow of water at the required temperature could pass. As the experimental material was placed between pairs of these vessels, silver, which is a good conductor of heat and not injurious to living specimens, was the metal chosen for the inner surface (against which the material would rest). Through one plate of a pair cold water was passed. In the first few experiments the temperature of tap water was lowered by circulating it through a copper coil placed in an insulated ice tank, and this cooled water passed through the apparatus. By this method the lowest temperature that could be obtained was 0-9° C , but the lowest average that could be maintained for long periods was 50 C. Later a lower temperature still was attained by abandoning the copper coil method and substituting one in which brine produced from a mixture of ice and salt was siphoned through the series of plates. This gave temperatures as low as — 8° C , but the average used was about — 30 C. as in the course of experiments it was often found fatal to develop-ment to go below — 50 C. Through the other plate was passed hot water, the temperature of which was kept constant by circulating it through a copper coil placed in a constant temperature bath electrically heated and regulated. The rate of flow was controlled by an adjustor fixed to the main tap. The maximum tem-perature used was 400 C , but it was found that any temperature from 28°-4O° C. might be more or less successful according to the susceptibility of the eggs, the degree of compression, etc. Latterly when the material was less fresh we found that it was not safe to use temperatures above 300 C. The actual temperature difference between the two poles of the egg has not yet been measured (a thermo-couple applied first to one pole and then to the other would probably prove to be the most satisfactory method), but allowing for the fall of temperature through the jelly the temperature difference must be greater than 50 C.
312 I. L. DEAN, M. E. SHAW and M. A. TAZELAAR
successive pairs were made as short as possible to reduce to a minimum the slight diminution in the temperature gradient due to radiation. To ensure a moist atmo-sphere round the material, water was gently pipetted between the edges of the plates at intervals. In the majority of the earlier experiments the plates were kept in the horizontal position. For side-to-side heating up to the neural plate stage the plates were kept vertical between wooden clamps on a stand, and in these experiments water had to be pipetted between the plates at much more frequent intervals to keep the material in a sufficiently moist condition.
Owing to the constant attention needed, experimental conditions were never continued for more than twelve hours together, but in some cases after a day's treatment the material was kept at a low temperature in a moist chamber to retard further development, and was then treated again the next day. Notes on the effects of the treatment were made from a binocular examination of the living specimens at the close of each experiment and specimens from each pair of chambers were then fixed, while the remainder were put in tap water in glass jars to go on developing. They were observed at intervals, one or two fixed at various stages of development and any necessary drawings, photographs and measurements made. Young tadpoles were fed on Spirogyra, older ones on small pieces of frog flesh.
The difficulty of sectioning frog's eggs is well known but we found successful the following methods of fixation.
A. Fixative for Frog's Eggs.
i gram ... ... ... Bichromate of potash 2-5 c.c Glacial acetic acid 5 „ Formalin
92 „ ... ... ... Water
Fix for 48 hours, changing the solution each day, at a low temperature. Rinse in water. Keep in 5 per cent, formalin in dark for at least two weeks, changing the formalin as often as it becomes discoloured, then keep in 70 per cent, alcohol for as long as desired.
B. Fixation Method for Frog's Eggs (Blastula and Gastrula stages).
Place eggs in 4 per cent, formalin heated to 75-850 C. and allow to remain in this solution for three to five minutes. The jelly is then easily removed with the aid of two needles. Keep indefinitely in 4 per cent, fomol. By this method the softness of the yolk is unchanged. Eggs fixed in this way should be embedded as follows: 70 per cent, alcohol, 30 minutes; 90 per cent, alcohol, 30 minutes; absolute alcohol, 20 minutes; toluol, 5 minutes; toluol-paraffin, 20-25 minutes; paraffin, 15 minutes.
Staining. Sections of eggs were stained in Erlich's haematoxylin, and sections
3. EXPERIMENTS AND RESULTS.
A. T H E EFFECT ON SEGMENTATION.
I. Apico-basal temperature gradient.
(a) Applied from fertilisation. Eggs placed about 1 cm. apart on the plates
and fertilised two hours before treatment. Duration of the experiment 7J hours; maximum and minimum temperatures used, 270 C. and — z° C. Two sets of plates used both lying horizontally, i.e. the eggs were in the normal position. Gradient antagonistic for the first pair of plates and adjuvant for the second pair. For convenience in description we shall term the first pair of plates c/h and the second pair h/c.
At the close of the experiment it was found that the segmentation, though still in a very early stage, had been markedly affected. In the h/c eggs, which were as a whole at a slightly later stage of development than the c/h, the animal hemisphere had divided into a considerable number of cells, and the vegetative hemisphere into very few cells (see PI. V, fig. A). No blastocoel had developed, a common feature after application of a temperature gradient, which we shall discuss later. In the c/h the animal cells were very much larger than in the h/c, and segmentation of the yolk cells had gone so far that they were about the same size as the animal cells and in one or two cases slightly smaller.
(b) Applied from 2-32 cell stage. Eggs fertilised the night before, kept in tap
water on ice over night, then placed on the plates at the 2-4 cell stage. Two pairs of horizontal plates used, one h/c, the other c/h, these acting as controls to each other. As in all other experiments, normal controls from the same fertilisation kept in water at a temperature intermediate between the highest and lowest temperature of the experimental gradient. Length of treatment in some cases as much as 12 consecutive hours, and in several experiments continued on the following day.
Consistent results were obtained throughout. Good contrasts in the effects of adjuvant and antagonistic gradients can be seen in PI. V, figs. B and C, showing eggs at about the same stage of development. The h/c (adjuvant gradient, Exp. C 3) is an egg from a one-day experiment which lasted 11J hours with maximum and minimum temperatures of 350 C. and 50 C. The c/h (Exp. Nz) was kept under experimental conditions for two days, the treatment lasting for 8 hours on the first day and i\ hours on the second day at maximum and minimum tempera-tures of 280 C. and — 6° C. In both experiments, C 3 and Nz, the c/h chambers
(N 2 A, C 3 A) were first and the h/c (N 2 B, C 3 B) second in the series. The animal
cells of C 3 B are markedly smaller than those of AT 2 A and of its control C 3 A. On the other hand, the yolk cells of N 2 A are very much smaller than those of
C 3 B, these being enormous with an average size five and a half times that of the
and also the diminution in size of the animal cells from the animal pole to the equator.
Eggs from these different experiments, N 2 and C 3, have been selected for description, as they show the maximum effects that were obtained during the 1927 experiments from the application of adjuvant and antagonistic gradients.
In the c/h (C 3 A) controlling the C 3 B egg already described, the animal cells are large but not larger at the pole than at the equator or larger than the vegetative cells. This is probably due to the fact that the egg is still at an early stage of segmentation, the maximum effects of both types of gradient being seen in middle segmentation. The blastocoel is smaller than in C 3 B h/c.
In the h/c (N 2 B) controlling N 2 A the animal pole has segmented into four layers of extremely small cells. The blastocoel is distinctly larger and the whole
egg has reached such a late stage of development that the yolk cells are slightly
smaller than those of the c/h (N 2 A).
Table I.
Measurements of cell size in eggs from experiments N 2 and C 3. {Eyepiece micrometer units.)
Experiment Animal pole Vegetative pole Ratio av. vegetative cell sizeav. animal cell size C3B ( h / c )
C3A ( c / h )
iV2B(h/c) iV2A(c/h)
av. 5 (max. 8) 9 (min. 5) 5
40 (max. 49)
av. 70
9 (min. 7) 28 (max. 35)
18 (min. 10)
71/1 1/1 1-8/10 22-2/10
Normal ratio av. vegetative cell size 10 10av. animal cell size _ 3^5 4
In the course of the experiments it was found that the highest temperatures used were unsuitable as the development of a number of eggs was inhibited, and the mortality among these eggs very high (this was presumably due to decreased resistance of the eggs, the frogs having then been kept on ice for several weeks). Experiments were then carried out to ascertain the maximum temperature which could be applied to the egg with safety, and it was found that for these less resistant eggs about 280 C. was the optimum temperature in the warm chamber. The egg shown in PI. V, fig. D, illustrates clearly the effects of inhibition at the animal pole, very large animal cells being scattered among the normal small animal cells. This egg and others in which there was inhibition at the animal pole showed, when sectioned, an entire absence of blastocoel. It is interesting to note that this absence of blastocoel was not confined to inhibited eggs, or, in fact, to eggs treated in any special way, small abnormal blastocoels or entire absence of blastocoel occurring occasionally with all types of gradient. This question is discussed more fully later. All these "inhibited" eggs died during gastrulation.
316 I. L. DEAN, M. E. SHAW, and M. A. TAZELAAR
II. Lateral temperature gradient,
{a) Applied from fertilisation. In this experiment the eggs were arranged on
the plates in such a position that they lay on one side with animal poles all orientated in one direction. They were then fertilised (using a very fine pipette) on that part of the animal hemisphere which ensured that the grey crescent would be formed
between the two plates when these were clamped together in the vertical position,
and the plane of symmetry of the eggs parallel to the plane of the plates. In this way the path of the temperature gradient through the egg was arranged to be transverse to its median axis, i.e. the future right and left sides of the embryo were subjected to heating and cooling. The duration of this experiment was six hours, and the maximum and minimum temperatures 330 C. and 2-8° C.
Some of the fertilisations were successful, about half of the eggs having segmented, and these mainly on one side (the heated side). In a photograph of a section of one of these eggs (PI. V, fig. E) complete cells are seen on the heated side, while cleavage furrows are only just forming on the opposite side. As is to be expected at such an early stage, the divisions are mostly at the animal pole, the yolk remaining practically undivided.
Text-fig. 1 shows a drawing of an egg from an experiment where the eggs were treated in exactly the same way as in the experiment described above, but segmentation was further advanced at the close of the experiment.
(b) Applied from the 2-4 cell stage. In this experiment the eggs were placed
gastrulation, so that the effect of the temperature gradient on later stages of development could not be determined.
Dr Carter, of Glasgow University, has suggested in conversation that, when a lateral gradient was applied in the 2-cell stage, there would in most specimens be a period during which the whole of one half of the egg would be heated, the whole of the rest cooled, since there would be no cell-walls to impede circulation of the contents: and a very similar result would arise after the 2nd or 3rd cleavage. This would result in the whole of one half of the egg getting a start over the other, and would at least contribute to the sharp line of demarcation sometimes observed between large and small blastomeres in later cleavage stages. We believe that this suggestion will help to explain the point, though possibly other factors are also at work. It will not, however, explain the sharp demarcation in the case of adjuvantly-treated eggs whose animal pole was inhibited by excessive heat.
B. EFFECT ON GASTRULATION AND EARLY EMBRYOS.
I. Apico-basal temperature gradient.
(a) Eggs treated in early gastrula stage (before formation of dorsal lip of
blastopore). Exps. M1 and M 2. The plates were set up in the usual way, the eggs being placed in the natural position, and two sets of plates being used, one
(M 1 c) h/c and the other (M 1 D) c/h. Experimental conditions were continued
for six hours and the maximum and minimum temperatures were 280 C. and — 5-2° C. After treatment the eggs were examined under binoculars and it was noticed that while in the h/c eggs the blastopore rim was never more than semi-circular, and in some cases not nearly so advanced, in the c/h eggs it was horse-shoe shaped to circular although still quite large. In a number of these c/h eggs the presence of abnormally pale areas was noted, as if the animal cells at the blastopore lip were drawn out and thin owing to active intucking, and without being reinforced by growth from the animal hemisphere. Sections showed what appeared in some ways to be a contradictory result. In the M 1 c h/c egg (Text-fig. 2), sectioned, the dorsal lip of the blastopore had grown down almost to the centre of the vegetative pole, whereas in the M 1 D c/h egg (Text-fig. 3) it had only reached a point just over half way between the equator and the vegetative pole, although the egg was definitely further advanced, the ventral lip of the blastopore being visible.
These results agree with the 1926 results of Huxley, who found that with antagonistic gradient the blastopore Up was formed farther away from the vegetative pole. He explained this as due to the greater heating of the lip region as against the animal pole and therefore the absence of the normal downpushing of the cells of the animal pole which carries the blastopore lip towards the vegetative pole. Similar results were obtained by Bellamy (1919).
This experiment was continued the next day for nine hours (Exp. M 2) and the results were similar to those obtained on the previous day, the effects of adjuvant and antagonistic gradients being intensified. In h/c eggs (see Text-fig. 4)
318 I. L. D E A N , M. E. SHAW and M. A. TAZELAAR
the blastophore was closed but the yolk plug fairly large, and only in one specimen was there any indication of neural plate or neural folds; on the other hand, in the c/h eggs (see Text-fig. 5) the yolk plug had already been almost completely with-drawn, the blastopore was small and slit like and the neural plate and even beginnings of neural folds were visible in most specimens.
Unfortunately all the h/c eggs were in a bad state of preservation and it was not possible to ascertain the degree of development and the germinal layers and size of archenteron, etc. A sagittal section (Text-fig. 5) of a c/h egg shows it to be definitely at a later stage of development than the M z c (h/c) egg (Text-fig. 4). The blastopore is nearly closed and the yolk plug small, the thickened ectoderm of the neural plate visible and also the beginning of the transverse neural fold at the anterior end. On comparison with a normal gastrula of the same stage of development it was clear that in the M 2 D gastrula the notochordal tissue was not so clearly marked off from the endoderm, while at the same time the ventral ectoderm was slightly more definite and clearly defined than in the normal specimen and the archenteron layer due to greater reduction in the amount of yolk. This result is easily understood when it is remembered that although at the commence-ment of the expericommence-ment the region where the formation of the blastopore takes place is heated and therefore gastrulation further advanced in the c/h eggs, later rotation takes place and during this process the anterior and dorsal region is cooled and the ventral region heated. Thus the egg to which an antagonistic gradient had been applied had reached a later stage of development than the one which received the opposite treatment, showing neural plate and notochordal tissue in spite of the low temperature applied dorsally. This was no doubt due to the fact that in the earlier stage of treatment the blastopore lips which form below the equator were stimulated to active growth by the high temperature applied ventrally, causing gastrulation and neural plate formation to take place earlier in spite of the fact that the dorsal lip was formed farther towards the animal pole than usual and the neural plate region cooled. The result of the dorsal cooling was only apparent on comparing the h/c egg with a normal control at the same stage of development.
(b) Eggs treated in yolk plug stage. In this case the eggs were placed under
experimental conditions when gastrulation was nearly completed. The treatment was continued for 8 hours and the results were similar to those obtained in the preceding experiment, the c/h eggs appearing to be, from an external examination, at a more advanced stage of development than the h/c.
the neural plate is markedly thickened and neural folds are present, but the differentiation of the nervous system proper to this stage (i.e. formation of neural crest) does not seem to have taken place. The notochord is not very clearly defined, nor is the mesoderm clearly marked off from the roof of the archenteron or the lateral parts o£ the neural plate. The mesoderm is well differentiated ventrally and there is a normal large archenteron.
The anterior ends of both these embryos were examined in transverse section, and it was found that in the h/c there are large neural folds anteriorly and an abnormally deep neural groove as though there had been a premature attempt at investigation to form the brain. In the c/h, nothing of a similar nature is seen, and in fact the neural groove is deeper and the neural folds much better defined in the trunk region. In the h/c the notochord and mesoderm are very indistinct posteriorly while in the c/h these structures become more distinct posteriorly, both this and the appearance of the anterior ends of the two embryos showing the effect of the rotation of the egg during the experiment.
Thus it is not simply that the c/h embryo has reached a later stage of develop-ment, although this appears to be the case from an external inspection because of the presence of larger neural folds and the embryo's greater size. Differentiation, at any rate as far as the notochord and mesoderm are concerned, has proceeded further dorsally in the h/c and further ventrally in the c/h. This is probably the case with the nervous system also, since there has been an attempt to form the brain in the anterior region of the h/c, but the effect is obscured in the trunk region by the greater amount of nervous tissue present, deeper neural groove and larger neural folds in the c/h specimen. This apparent contradiction in the degree of differentiation of the nervous system is probably to be ascribed partially to rotation of the egg during the experiment, since the notochord and mesoderm show the same effect to a lesser degree, but apart from this there has certainly been a great proliferation of nervous tissue in the c/h specimen and why low temperature applied dorsally should have this effect is not at all clear. The same problem cropped up again and again in the course of our experiments, quantitative effects of treatment (due to proliferation of tissue) and also qualitative effects (due to differentiation and growth) being both found, often in the same specimen. This is more fully discussed later, but Huxley (1927) suggests that the problem is a complex one and involves a knowledge of the stage of determination and differentiation of each organ and part of the body (in this particular case germinal layer) at the time of treatment.
320 I. L. DEAN, M. E. SHAW and M. A. TAZELAAR
slightly further advanced (as shown chiefly by the degree of development of the sense organs) even when compared with c/h embryos with complete closure of neural folds. Not only this, but it seems that the brain is actually larger in proportion to the head in the h/c tadpoles (see Text-figs. 8 and 9).
Another experiment was carried out in which the eggs were .first treated in the yolk plug stage and then exposed to experimental conditions again the next
10 A. 10 B.
A. 11B.
Text-figs. 10, 11. Camera lucida drawings of tadpoles at the end of the second day of treatment with adjuvant (10) and antagonistic (11) temperature gradients first applied in the late yolk plug stage / c J • .u\ \ T\ -i • j. h. ant. and dors. , ,
(rotation of egg during experiment). 10 A. Dorsal view of tadpole showing c. post, and vent.
neural folds closed anteriorly but slightly open posteriorly. 10 B. Side view same tadpole showing „ ., , , ~. , . h. ant. and dors. . , ,
small tail bud. Dorsal view -z tadpole showing neural folds closed posteriorly but c. post, and vent. r J not yet closed anteriorly. 11B. Side view same tadpole showing well-developed tail bud. a, sense plate;
b, gill plate; c, branchial arch; d, gill slit; e, brain; / , pronephros; g, pronephric duct; h, open
medullary fold; k, tail bud; I, sucker; s, somite.
dorsally were further advanced than those heated posteriorly and ventrally. Specific results were obtained, some of the c/h embryos showing failure to close the neural folds anteriorly, but complete closure posteriorly, while some of the h/c showed the reverse effect although to a less extent (see Text-figs. 10 A, 11 A). Another interesting result is seen in the relative stages of development (i.e. sizes) of head and tail bud (see Text-figs. 10 B, 11 B). In the tadpole cooled anteriorly and dorsally the tail bud is very well developed even in small specimens, while in those heated anteriorly and dorsally the tail bud is poorly developed even in larger embryos. The opposite result is also seen, i.e. where the tail bud is poorly developed (h/c) the head is well marked off from the trunk and the branchial arches and suckers at a more advanced stage of development than in the c/h.
II. Lateral temperature gradient.
(a) Eggs treated at early dorsal lip stage. The eggs were treated for six hours,
left to develop under normal conditions until neural folds appeared, and then preserved. Owing to the difficulty of manipulation it was impossible in this experiment to say which side of the embryo had been heated, but PI. VI, fig. G shows the striking asymmetry of neural folds developed in one of the embryos.
Serial sections of this embryo show that the neural fold on the right side commences more anteriorly than that on the left and is consistently larger down the whole length of the embryo, although disappearing in the posterior sections before the left fold. In the trunk region the mesoderm on the left side is more clearly defined than the mesoderm on the right, but in the anterior region the converse effect is seen, and the mesoderm on the right forms a thicker band throughout the whole length of the embryo. These facts lead us to suppose that heat was applied to the right side of the embryo and caused greater growth of tissue on that side.
(b) Eggs treated at dorsal lip stage. Eggs were chosen which showed a
well-marked dorsal lip and placed in such a position that the blastopore would be situated between and not touching the plates when they were clamped together in the vertical position, and the high and low temperatures applied to the future right and left sides of the embryo. No asymmetry was apparent at the close of the experiment and the embryos were left to develop under normal conditions. Five days later all those heated left side had died, but those heated right side showed striking asymmetry, which is discussed later under Regulation.
(c) Eggs treated in early neural plate stages. Eggs at this stage were easily
orientated so that the future right and left sides were in contact with the plates (placed in the vertical position) and the gradient passes through the embryo from side to side. The eggs were treated for eight hours, and then left to develop under normal conditions until the next day, when it was found that both the heated right and heated left specimens showed marked asymmetry.
322 I. L. DEAN, M. E. SHAW and M. A. TAZELAAR
folds, bent round to the unheated side. This specimen was observed and drawn at intervals (see notes under Regulation).
(d) Eggs treated at neural fold stage. At this stage the right and left sides are
easily determined, and it is a fairly easy task to orientate the embryos so that a known side is heated. These specimens were treated for nine hours, at the end of which time they had reached the early tail bud stage. Under the binocular it was seen that the branchial arches, the pronephric eminence and pronephric duct were all appreciably larger on the heated side. Serial sections showed that a striking asymmetry had been produced especially in the eye region. On the heated side the early optic vesicle and the optic invagination were at a later stage of development than those on the unheated side, and the tubule formation of the pronephros was more clearly defined. The remainder were left to continue development under normal conditions and will be discussed under Regulation.
C. EFFECT ON OLDER EMBRYOS.
A number of experiments were carried out on tadpoles which had reached the tail bud stage. In these cases the manipulation was easier and considerably more satisfactory as the embryos had elongated sufficiently to lie on one side between horizontal plates. In the great majority of cases the jelly was not removed, but in two later experiments an attempt was made to steepen the gradient and obtain more marked effects by removing the embryos from the jelly. This, however, was not very successful as the embryos were apt to be injured by direct contact with the hot plate, and there was great difficulty in preventing them from drying up. In one case only was the experiment continued for a second day, as the embryos had by then generally reached too late a stage of development for further treatment.
The results were on the whole consistent, similar effects showing in a varying degree in different specimens and experiments. Two constant results which were generally obvious at the close of the treatment were a slight enlargement of the branchial region and of the pronephros and its duct on the heated side. Great asymmetry was seen in the yolk region of the trunk but was so inconsistent, showing enlargement either on the heated or cooled side, that it was found impossible to draw any conclusions. These notes were made from living specimens and it was observed that very slight flexure of the trunk often caused relatively enormous projection of the yolk region on the flexed side, so that it is probable that these differences in the amount of yolk on heated and unheated sides were due to this and had no other significance.
mass of nephrogenous tissue which in one region shows the beginning of a central cavity very far from distinct; on the heated side tubule formation is well advanced. This specimen is interesting in that it shows great multiplication of undifferentiated tissue on the cooled side, in the pronephric region, but greater differentiation on the heated side. This has previously been noted in the case of neurulae, and in the later stages was found to be a fairly common effect. In the yolk the cells are much smaller and more numerous on the heated side, indicating more active cell-division on that side.
Though the ear and olfactory pit gave no perceptible results at this stage, the eye showed throughout the most constant asymmetries that were obtained in any organ: the eye on the heated side was invariably at a later stage of development. The primary optic vesicle was narrower and more compressed owing to the greater invagination and development of the retinal layer. In one specimen (see PI. VI, fig. H) the retinal layer, markedly thicker and more invaginated on the heated side, is actually folded on itself concertinar-wise, an apparently mechanical effect due to excessive growth and an exaggeration of the very slight creasing which is occasionally seen. Murray and Huxley (1925) in the grafts on the chick obtained several cases of this mechanical effect of overgrowth in the eye. No difference was noted in the development of the lens of the two sides. Measurements made of the maximum diameter of the optic vesicle in this stage of the early optic cup showed that the actual size of the optic vesicle was greater on the heated side (see Table II).
Table II.
Asymmetry of the eye of tadpoles subjected to lateral temperature gradient. (Micrometer eyepiece units)
Maximum length of optic vesicle Heated side
13-0 15-5 22-5 iS-5 i8-5 I3-5
Cooled side 12-0 14-0 20-5 14-4 17-8 13-0
% difference
10
2 \ heated 2 days
6-3J
4 ° | heated 1 day
In two experiments treatment was continued over a second day, as usual the material having been kept on ice over night. A slight accentuation of the asymmetry of the pronephros was noted, but a slight diminution of that of the branchial arches. Eye measurements showed a distinct increase in the effect (see Table II).
D. REGULATION.
324 I. L- D E A N , M. E. SHAW and M. A. TAZELAAR
far as possible the material from each experiment was followed up through later stages, and certain results, which will be put forward, seem on the whole to be fairly conclusive.
Apico-basal temperature gradients.
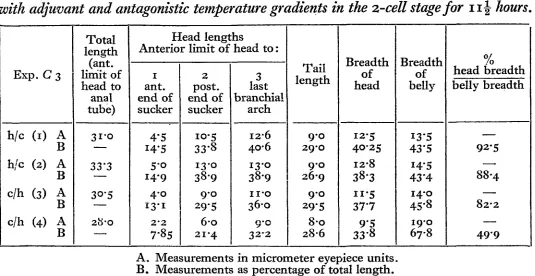
The effects of an apico-basal temperature gradient on the segmentation of the egg have been described and shown to be very striking, the time relations of cell division being much modified. Later stages were examined and measurements made to ascertain whether any change of proportions had been effected. Accurate measurements with an ocular micrometer could be made with comparative ease on the early tail bud stage, but the twisting of the tail and the difficulty of finding an accurate head measurement in the later stages made the results obtained on older tadpoles unreliable. Measurements of embryos at the early tail bud stage, subjected to an adjuvant gradient in the egg stage (Exp. C3), showed that the relative sizes of the head and tail had definitely been influenced (see Table III).
Table I I I .
Showing relative head size in tadpoles {early tail bud stage) obtained from eggs treated with adjuvant and antagonistic temperature gradients in the z-cell stage for 11J hours.
Exp. C 3
h/c (1) A B h/c (2) A B c/h (3) A B c/h (4) A
B Total length (ant. limit of head to anal tube) 31-0 — 33'3 3O-5 — 28-0 — Head lengths Anterior limit of
1 ant. end of sucker 4-5 14-5 14-9
4 - 0 13-1
2 - 2
7-85 2 post. end of sucker 10-5 33-8 13-0 38-9
9 - 0
29*5 6-o 21-4 head to: 3 last branchial arch
1 2 6
40-6
13-0
38-9 u - o
36-0
9 - 0 32-2
Tail
length
9 0 29-0
9 - 0
26-9
9 - 0
29-5 8-o 28-6 Breadth of head 12-5 40-25 I2'8 38-3
" • 5
37-7 9'5 33-8 Breadth of belly 13-5 43*5 14-5 43'4 14-0 45-8 19-0 67-8 rt / 0/ /o head breadth belly breadth — 92'5 — 88-4 — 82-2 —
4 9 9 A. Measurements in micrometer eyepiece units.
B. Measurements as percentage of total length.
[image:16.648.59.594.430.706.2]Sections made of (1) and (3), Table III, showed that although at apparently similar stages of development the rudiments of the sense organs showed a relatively greater size in the h/c. The following table gives measurements of the size of the eye relative to the brain in these two specimens.
Table IV.
Exp. C 3
h/c c/h Width of brain 36-0 3O-5 Depth of brain 43-o 35'5 Max. eye depth I9'S 13-6 0/ /o d e
The measurements set forth in Table IV were made on preserved material. Owing to the necessity for economy of material, living tadpoles were anaesthetised in chloratone (1 in 1000 in tap water) and drawings, which were analysed for body measurements later, made under the camera lucida. In these tadpoles twisting of the body made it difficult to find the head and body lengths accurately from drawings. The results obtained from measuring head and body breadths, however, are very striking and again show that the head is relatively larger in those in which the animal pole of the egg was heated (see Table V).
[image:17.648.90.545.361.566.2]Table V.
Showing the relative head size in tadpoles obtained from eggs treated with adjuvant and antagonistic gradients in the z-cell stage. Measurements made on camera lucida drawings of tadpoles a few days after hatching.
Exp.
C3( h / c )
C 3 (c/h)
Breadth of head mm. 2*2O i"75 1-70 1-72 1-82 1-85 i-8o 1*55 1-50 1-52 i-55 Breadth of body mm. 1-94 i'73 i-59 i-6o 1-67 **53 1-74 1-65 1-70 1-62 1-89 0/ /o head breadth body breadth 113-2 IOI-I 106-9 107-4 109-0 120-9 IO3-5 94-0 88-2 93-9 82-0
These measurements were the only ones of this series which were consistent throughout. On the other hand, even these results may be unreliable for as a general rule the h/c tadpoles are a little further advanced than the c/h, and at this period of development the body breadth decreases with growth owing to the reduction of yolk. This would tend to increase the ratio . , ,—-5-r in the caseJ
body breadth of the h/c tadpoles.
Further measurements were made at the external gill stage where again the difficulty of finding the head length with sufficient accuracy made quantitative results impossible. Even if, however, it had been possible to make accurate measurements it was apparent that any differences that might exist would have been small. In another series of tadpoles treated with adjuvant gradient in the egg and where segmentation had been most markedly affected, measurements on anaesthetised tadpoles in four later stages of development were made, random samples being taken from the experimental material (all h/c) and controls.
When the external gills were practically covered over by the operculum (stage 1), , . « , head and trunk analysis of measurements showed that the average percentage t o t aHength
326 I. L. D E A N , M. E. SHAW and M. A. TAZELAAR
weeks later when the internal gills had then been formed (the rate of growth throughout was slow and the limb buds had not yet appeared) was measured by means of a stage micrometer. The resulting percentages were similar to those for the preceding stage—experimental 43-6, controls 39-85 (9 and 10 specimens respectively). At the next stage measured (3) 12 days later, the remaining living specimens from the previous stage showed the following percentages: experimental 40*8, controls 37*85 (6 specimens of each). The head and trunk length in stage (1) was taken to the junction of trunk and tail, as seen in dorsal view, stages (2) and (3) to the anal tube, so that the figures of (1) are not comparable with those of (2) and (3). One hesitates to draw too firm conclusions from these figures, but even
Text-fig. 12. Photographs of a markedly asymmetrical tadpole (heated right side) in the early tail bud stage taken at two different stages of its development: (a) early stage—dorsal view, (b) later stage— dorsal view. Both (a) and (6) show bending of the axis towards the cooled side, apparent anterior shifting of the right eye, greater growth on the right side.
Text-fig. 14. Photograph of a markedly asymmetrical tadpole (heated left side in the early tail bud stage for 9 hours), ventral view, showing bending of the axis towards the cooled side and asymmetrical position of the mouth.
Lateral temperature gradients.
Owing to the difficulty of ascertaining the right and left sides of material treated during segmentation with a side to side temperature gradient, and the high mortality following these experiments, we have been unable to follow the further development of their later stages. Of those treated with a side to side gradient in the dorsal lip stage nothing was noted until the external gills were being resorbed, when two tadpoles were observed to show definite asymmetry. The heated side of the head was very noticeably larger than the unheated side and the axis of the body in the head region twisted towards the unheated side. Also the mouth was lying well over to the unheated side and the eye and sucker of the heated side farther forward. This asymmetry continued in one specimen until it died (it unfortunately disintegrated before fixation), in the other specimen it became rather less apparent before it also died in a generally abnormal and oedematous condition (see Text-fig. 12 (a) and (&)).
The embryo shown in Text-fig. 13, heated on the right in the neural plate stage, which, as previously described, showed a
definite asymmetry, was kept under observation. This specimen and one other, which died at an early stage, were the only two of this experiment which showed any external asymmetry—the others were kept for some weeks but appeared to be quite symmetrical at all stages. Ten days after treatment it was apparent that the right side of the head and trunk was larger than the left, and though the main axis of the body did not yet curve visibly to the left, the right eye was a little
more anterior than the left. Very slight curvature of Text-fig. 13. Camera lucida
trie m a i n s»vt<a fiersime j m n a r e n t five r1av<? later F m i r d r a w i nS o f a n embryo exposed
me mam axis became apparent nve aays later, r o u r t o k t e r a l t e m p e r a t u r e gradient
days later still the tadpole died and unfortunately was in the yolk plug and early unfit for sectioning neural plate stage. Embryo
&
heated right, showing larger
T h e effect of a lateral temperature gradient on the right side and bending of the neural fold stage has already been described (p. 321). ™ towards the left, a, sense
o r 1 P i J 1 ^ 1£ 1 PI a t e; b* g*11 Pl a t e; c> o u t e r
Sections or tadpoles preserved 24 hours alter the close meduilary plate; d, inner
me-of treatment showed them to be still asymmetrical, the dullary plate.
328 I. L. D E A N , M. E. SHAW and M. A. TAZELAAR
really marked difference in size, that on the heated side being notably larger. In this case it would appear that regulation towards the normal had taken place to a certain extent since the pronephros only still showed asymmetry.
The fate of the marked effects already noted in those tadpoles heated at the tail bud stage was followed through later stages. The external gills often showed irregularities, very seldom being equal and occasionally being developed on one side only, which might be either the heated side or the cooled side. Controls showing this too, these asymmetries were disregarded and in most cases complete symmetry was gained after their resorption. Text-fig. 14 shows an extremely asymmetrical tadpole which was treated in the tail bud stage and retained its asymmetry till it died.
Visibly asymmetrical specimens were sectioned about three weeks after treat-ment (in the tail bud stage) and all showed similar divergencies from the normal. In every case the greatest and most easily detected differences between the two sides of the embryo appeared in the optic cups, the retinal layer being thicker and further invaginated and the maximum depth of the vesicle greater on the heated side.
Table VI.
Asymmetry of the eye in tadpoles treated with lateral temperature gradient in the tail hid stage. {Measurements in micrometer eyepiece units.)
Specimen
r
2
3
Maximum length optic vesicle
Heated side
22-5 iS-5 25-5
Cooled side
20-5 14-0 2I'O
% difference in size between heated and cooled
sides
12-2
107 16-6
Table VII.
Micrometer eyepiece units.
Heated side 15-5 14*5 12-5 7-0
A
I*'
21 7'2——
Cooled side
i3'7
?3-° i i - 5
6 7 6-3 7-2 — 0/ /o difference
1 3 1
" ' 5 8-7 4-8
- i - 6
o-o — Heated side 6 0 5-o 4-8 4*3
B I5"2
(5'i
4*5
Cooled side
5'7
4 - 0 4 - 0
3 9 4 - 2 4 - 2 4-0 /o difference 5'25 25-0 20-0 10-3 23-8 21-45 12-5
A and B presumably represent the branchial and sciatic enlargements.
The gut is much enlarged and lies well over towards the cooled side. The limb bud on the cooled side was a shade larger than that on the heated side. In no case could we observe any parallel asymmetry in the limb buds—if any existed, either way, it was very slight and probably within the limits of normal variation.
It should be noted that one asymmetrical specimen developed where the cooled side showed the greater size and development. Whether this was an inhibiting effect of excessive heat on a more sensitive embryo, or whether some other factor was involved, we cannot say. Controls might show slight asymmetries but not nearly so frequently or so markedly as did the experimental tadpoles. The question of these asymmetries need a further and more statistical treatment than has been possible in this investigation.
STATEMENT OF GENERAL RESULTS AND DISCUSSION.
Segmentation.
330 I. L. D E A N , M. E. SHAW and M. A. TAZELAAR
the smallest cells being at the equator and the largest at the two poles, and to reduce or reverse the normal disparity in size between the extreme animal and vegetative cells.
It was found difficult to contrast the effects of adjuvant and antagonistic gradients on cell size, owing to the fact that the h/c had almost invariably reached a further stage of development than the c/h from the same experiment. Extremely interesting is such a case as that of Exp. N 2 (previously referred to), where in the h/c eggj owing to its more advanced stage of development, the yolk cells which were cooled are actually smaller than the yolk cells of the c/h which were heated. The effects of adjuvant and antagonistic gradients on cell size can still be traced in the early gastrula, the animal cells of the h/c being appreciably smaller than those of the c/h (no difference in size of vegetative pole cells could be detected). At later stages of development there is no difference as regards size between heated and cooled animal cells, indicating that after the early gastrula stage a limit of cell size is reached.
We have mentioned that absence or reduction in size of the blastocoel was frequently induced by all types of treatment. Bellamy (1919) induced absence or great reduction of the blastocoel by differential inhibition in toxic solutions—a case somewhat similar to ours. There seems to be a tendency for the blastocoel to be less well developed in the c/h eggs, and this may well be due to inhibition of the sensitive animal cells and consequently no downward movement from the animal pole (cf. Bellamy, 1919). A similar explanation would account for absence of blastocoel in "inhibited" eggs. As a similar effect was produced in a certain (fairly small) percentage pf cases by all methods of treatment, it may be due to some experimental factor such as too great pressure or absence of sufficient moisture, but it seems more probable that in general the cause is to be found in lack of synchrony in rate of segmentation in the two hemispheres and consequent abnormal cell sizes and delayed (or accelerated) cell movements.
Our measurements on tadpoles which had developed from eggs treated with antagonistic and adjuvant gradients showed that there was a slight but appreciable alteration of proportions, the c/h eggs developing into microcephalous tadpoles and vice versa. It was unfortunate that we were unable to collect later data from those eggs which were treated with lateral temperature gradient in the segmentation stage.
of determination of the different organs and parts along the axis, the head extending farther back when this region is heated and vice versa. This would be in accord with the general postulates of Child's Theory. See Child (1915, 1924).
Gastrulation.
The results obtained by the application of apico-basal temperature gradients to the egg during gastrulation agree with Huxley's 1926 results. As has been already described, with antagonistic gradient the dorsal lip of the blastopore is formed farther away from the vegetative pole than normally. Similar results were obtained by Bellamy (1919). The formation of the dorsal lip of the blastopore nearer the animal pole than usual in c/h eggs may be explained as due to the absence of the downpushing and very active cell division which normally goes on in the animal hemisphere especially in the region of the dorsal lip: the earlier gastrulation of these eggs may be explained as a simple stimulation of the region below the equator from which the blastopore lips form, and a continuence of this stimulation while the blastopore is closing. Although the result of applying a temperature gradient during gastrulation appears to be simply an acceleration of development in the c/h eggs, later the differential effect of the gradient becomes evident, the germinal layers being poorly defined in the dorsal region, but well developed ventrally in the c/h eggs and vice versa in the h/c eggs. Later still this effect becomes even more evident, the head and dorsal region being definitely inhibited in the c/h and the posterior region inhibited in the h/c. Whether this differential effect of the temperature gradient causes any permanent changes in the proportions of the tadpole has not yet been worked out, but that the relative head sizes are affected in the tail bud stage is evident (see Text-figs. 10 and 11). This is a problem for investigation next spring.
As regards side-to-side treatment during gastrula and neurula stages, the results were usually of a similar nature, namely, a quicker rate of development and larger amount of tissue on the heated side, giving rise to asymmetry most apparent externally in the bending of the axis towards the heated side, thus causing apparent shifting forward of the eye and the sucker on the heated side.
A point of interest that arises here is that asymmetrical development of the eyes can be brought about by the application of a temperature gradient in a very early stage of gastrulation, i.e. the dorsal lip stage. Huxley (1924) suggests that the brain is already chemo-differentiated in this stage, and the work of Durken (1926), who transplanted pieces of the frog's blastula and obtained parts of brain and ear vesicle from them, seems to bear this out (also see Vogt, 1927). This seems to suggest that the eye rudiment as well as that of the brain is determined at a very early stage. Spemann's work (1925) shows that the eye rudiment is definitely determined in the neurula stage.
332 I. L. D E A N , M. E. SHAW and M. A. TAZELAAR
Later Embryos.
Considerable differential growth resulting in asymmetry was produced by the application of temperature gradients to tadpoles in which the main processes of chemo-differentiation had been completed and visible differentiation and growth had begun. These tadpoles were all treated in the tail bud stage and the results are of a similar nature to those obtained by side-to-side heating during gastrulation, except that structures that are in an active state of growth differentiation at this stage, e.g. pronephros and its duct, branchial region, ear, etc., are affected as well as the eye region and general greater growth on the heated side.
The brain and sense organs have by the early experiments of Bellamy (1919) been shown to be the parts most inhibited by toxic solutions and therefore the most susceptible parts of this organism. Hyman (1927) working on chick embryos has shown the eye region to be especially susceptible. Our results on these latter embryos seem to confirm these results, for the eye was consistently larger on the heated side and was usually at a further stage of development. In only one case was asymmetry of the central nervous system itself obtained (PI. VI, fig. K)— a rather puzzling result.
Regulation.
The problem of regulation is a complex one and it has been impossible to work it out by any means fully this year. Measurements made on macrocephalous and control tadpoles at various stages of development seemed to indicate that a certain amount of regulation took place, although owing to the difficulty of getting an accurate measurement of the head this result is by no means conclusive. There are two possibilities—the actual level of determination of the head may have been altered, or the alteration in relative head size may be an expression of the different time relations of development, the heated head, for example, being at a later stage of development and therefore larger than the cooled trunk and tail regions. If .his is the true explanation, complete regulation might be expected to take place on attainment of normal growth rates. Only further experiments with (if possible) more intense stimulation and certainly longer treatment to induce more marked effects can settle this point. Very marked microcephalous forms have been produced in the frog (Bellamy, 1919; Cotronei, 1921,1922 a and b) and in fish (Stockard, 1909) by severe inhibition (no regulation), so that it is possible that both the time relations of development and the axial gradient relations of the parts have been affected in our case, but that the former outweigh the latter due to relatively weak stimulation and inhibition, leading in course of development to almost complete regulation.
If the amount of asymmetry produced is below this point, the embryo can regulate back to normal; if above, the asymmetry continues and may possibly become exaggerated. Further evidence, however, of a more statistical nature is needed before this very tentative view can be considered seriously.
Considerable light has been thrown on this problem by Huxley (1927):
We thus have to deal it appears with two problems. There is (1) a question of time relations, and (2) one of differential growth. If the organs of one side merely get a start in development over those of the other without also attaining a size difference, the asym-metry, we may suppose, is a temporary one only. If, on the other hand, the amount of material which is destined to be affected by the differentiating process is differentially multiplied on the two sides, it would appear that the difference is not afterwards obliterated (or possibly small differences are regulated, large ones not). Questions of the time of treatment in relation to the development of each organ would thus be expected to play a part, and the problem is seen to be a highly complex one of which these experiments have only touched the fringe.
This satisfactorily explains our contradictory results on the regulation of asymmetries. Here there is no question of alteration of axial gradient relations, and in fact it may be supposed that by this time, chemo-differentiation and deter-mination of organs having taken place with relation to the original axial gradient, this is too stable to be influenced by such relatively slight stimuli as the temperature gradients applied in the present series of experiments.
Conclusion.
In conclusion it may be said that no such marked alteration of proportions of the parts of the tadpole as was expected from the work of Bellamy (1919), Stockard (1909), Cotronei (1921,1922a and b) was obtained, but this is in all probability due to the comparative mildness of the stimulating and inhibiting agents used. We were working with an agent in no way abnormal or injurious to the organism. Normal development was possible at either the maximum or minimum temperature used (except where inhibiting temperatures were used unintentionally) and abnormal results were obtained by applying differential treatment to the various regions of the egg or embryo and causing abnormal rates of development. Results from later stages indicate that the exact effect produced depends on the organs differen-tiating at the time of treatment, showing that this method of tackling the problem of developmental mechanics should prove very useful, especially as the temperatures used are in no way abnormal to the embryo, and the method is therefore free from most of the objections to the use of other external agencies causing teratological development.
SUMMARY.
1. Temperature gradients were passed through the developing frog's egg and embryos. These gradients were applied either (a) apico-basally, when they were either (i) adjuvant, or (ii) antagonistic to the egg's own main gradient; or (b) trans-versely to the egg's main axis—lateral gradients.
334 I- L- D E A N , M. E. SHAW and M. A. TAZELAAR
2. (a) By this means considerable modification of segmentation and of cell size was induced, and was especially marked in the mid-blastula. Adjuvant gradients accentuated the normal differences in cell size between the animal and vegetative poles. Antagonistic gradients produced a double gradient in cell size, the smallest cells being in the region of the equator, and animal cells, in extreme cases, larger than yolk cells.
(b) Several cases of the non-formation or obliteration of the blastocoel were
obtained by all methods of treatment.
(c) Too high temperature with adjuvant gradient produced inhibition at
the animal pole, the large retarded cells being very sharply marked off from the surrounding small cells.
(d) Lateral gradients produced a great difference in cell size on the two sides
of the eggy and, as in the cases of "inhibition," a sharp line of demarcation may
appear between the large cells of the cooled side and the small cells of the heated side.
(e) When two sets of exactly similar eggs were treated simultaneously in
opposite ways, then those subjected to the adjuvant gradient were always, at the close of the experiment, at a more advanced stage of development than those subjected to an antagonistic gradient. Because of this the yolk cells of the " adjuvant" eggs were smaller than those of the "antagonistic" eggs, although the former were cooled and the latter heated.
(/) There seems to be a slight permanent effect of the gradient applied during segmentation. Eggs treated with antagonistic gradient tend to develop into microcephalous tadpoles and vice versa.
3. (a) Antagonistic gradients during gastrulation cause a reduction of the gastrular angle. (For definition see Bellamy (1919).)
(b) Antagonistic gradient causes the eggs to gastrulate sooner than adjuvant
eggs under exactly similar experimental conditions.
(c) In the neurula stage the differential effect of the gradient is seen in the
inhibition of the head and dorsal region in those subjected to antagonistic gradient, and inhibition of tail and ventral region in those subjected to adjuvant gradient.
(d) Whether this alteration of relative sizes of head and tail regions is
maintained in later development has not yet been ascertained.
(e) Eggs exposed to lateral gradients in all stages of gastrulation showed
marked asymmetries, some of which were apparently regulated later, while others persisted till the death of the tadpole.
REFERENCES.
BAUTZMANN, H. (1926). " Experimentelle Untersuchungen zur Abrengung des
Organisations-zentrums bei Triton taeniatiis. Mit einem Anhang: Ober Induktion durch Blastulamaterial."
Arch. Ent. Meek. 108, 283.
BELLAMY, A. W. (1919). "Differential susceptibility as a basis for modification and control of
early development in the frog. I." Biol. Bull. 37, 312.
(1921). "Note concerning the polarity of the frog's egg. A correction." Biol. Bull. 4 1 , 351. • (1922). " Differential susceptibility, etc. I I . " Amer. Journ. Anat. 30.
BELLAMY, A. W. and CHILD, C. M. (1924). "Susceptibility in Amphibian development." Proc.
Roy. Soc. Lond. B, 94, 132-45.
CHILD, C. M. (1915). Individuality in Organisms. Chicago.
(1924). The Physiological Foundations of Behaviour. New York: Henry Holt.
COPENHAVER, W. M. (1926). "Experiments on the development of the heart of
Amblystomapuncta-tum." Journ. Exp. Zool. 43, 321.
COTRONEI, G. (1921). " I processi di inhibizione differenziale nel vestibolo boccale degli Anfibi
Anuri." Riv. di biol. 3, 28.
(1922a)." Susceptibilita differenziale, gradiente assiale e rapporti tra correlazione e differenzia-zione." Rendiconti d. R. Accad. Nat. dei IAncei (Sci. fisiche mat. e naturali), 31, ser. 5, 473—6.
(1922 b). " I processi di inhibizione differenziale nella regione olfactoria degli Anfibi Anuri."
Rendiconti d. R. Accad. Nat. dei Lincei (Sci. fisiche mat. e naturali), 3 1 , ser. 5, 433-6.
DARESTE, C. (1877). Recherches sur la Production Artifidelle des Monstruosites. Paris, 1877.
DURKEN, B. (1926). "Das Verhalten embryonaler Zellen im Interplantat." Arch.Ent. Mech. 107,
727.
EKMAN, G. (1925). "Experimentelle Beitrage zur Herzentwicklung der Amphibien." Arch. Ent.
Mech. 106, 320.
GEENITZ, B. (19256). "Zur weiteren Analyse des Organisation-zentrums." Zeit. Ind. Abst. und
Vererb. 37, 117.
GOLDSCHMIDT, R. (1927). Physiologische Theorie der Vererbung. Berlin: Julius Springer, 1927. HUXLEY, J. S. (1924). "Early embryonic differentiation." Nature, 23, Feb. 1924.
(1926). " Modification of development by means of temperature gradients." Anat. Rec. 34. (1927). "Modification of development by means of temperature gradients." Arch. Ent.
Mech. 112, 480.
HYMAN, L. H. (1927). " T h e metabolic gradients of Vertebrate embryos. III. The chick." Biol.
Bull. 52, 1.
MURRAY, P. D. F . and HUXLEY, J. S. (1925). " T h e development of grafted embryonic fragments
of the chick." Brit. Journ. Exp. Biol. 3, 9.
SPEMANN, H. (1925). "Some Factors of Animal Development." Brit. Journ. of Exp. Biol. 2, 493.
SPEMANN, H. and MANGOLD, H . (1924). "Ober Induktion von Embryonalanlagen durch Implanta-tion artfremder Organisatoren." Arch. Mikr. Anat. und Ent. Mech. 100, 597.
STOCKARD, C. R. (1909). " The development of artificially produced cyclopean fish. The magnesium
embryo." Journ. Exp. Zool. 6, 2, 285.
STOHR, P. Jun. (1925). " Experimentelle Studien an embryonalen Ampbibienherzen." Arch. Ent.
Mech. 106, 409.
VOGT, W. (1927). " Uber Hemmung der Formbildung an einer Halfte des Keimes." Anat. Anzeiger,
63.
WARYNSKI, ^ . and FOL, H. (1884). "Recherches experimentales sur la cause de quelques mon-struosites simples, et de divers processus embryogeniques." Recueil Zoologique Suisse, 1,
336 I. L. DEAN, M. E. SHAW and M. A. TAZELAAR
DESCRIPTION OF TEXT-FIGS. 1-9.
Text-fig. 1. Camera lucida drawing of the dorsal pole of an egg treated with lateral temperature gradient from fertilisation for n hours.
Text-figs. 2, 3. Effect of adjuvant (2) and antagonistic (3) gradients on gastrulation. 2. Sagittal section of an h/c egg. Note extreme ventral position of the dorsal lip of the blastopore and absence of the ventral lip. 3. Sagittal section of c/h egg. Note dorsal lip not yet grown down to the centre of the vegetative pole but ventral lip of the blastopore and yolk plug clearly defined.
Text-figs. 4, 5. Effect of adjuvant (4) and antagonistic (5) gradients on late gastrulation. 4. Sagittal section of an h/c egg. Note large yolk plug. (Due to disintegration of the egg it is not possible to represent the germinal layers.) 5. Sagittal section of c/h egg. Note egg at a later stage of develop-ment than (4), small slit-like blastopore and yolk plug almost withdrawn.
Text-figs. 6, 7. The effects of antagonistic (6) and adjuvant (7) temperature gradients on late gastrulation and the formation of the neurula. 6. Transverse section of a c/h egg, trunk region. Note great multiplication of nervous tissue and formation of neural folds; notochord and mesoderm poorly developed dorsally but mesoderm well differentiated ventrally. 7. Transverse section of h/c egg trunk region. Note absence of thickened neural plate and small neural folds; clear delimita-tion of the notochord and mesoderm dorsally but absence of ventral mesoderm and extremely small abnormal archenteron.
Text-figs. 8,9. Transverse sections of embryos exposed to temperature gradients in the yolk plug stage (before rotation of the egg). 8. Antagonistic gradient. Note slight degree of development of the optic vesicles and small relative size of the grain. 9. Adjuvant gradient. Note more advanced stage of development of the optic vesicles.
DESCRIPTION OF PLATES.
PLATE V.
FIG. A. Effect of adjuvant gradient on early segmentation. Note large vegetative cells—absence of segmentation cavity. Treated from fertilisation for 7J hours. Section through centre of egg. FIG. B. Extreme effect of adjuvant gradient on later segmentation. Note very small animal pole cells and large yolk cells and irregular blastocoel. Treated from 2-cell stage for n £ hours. Section through centre of egg.
FIG. C. Extreme effect of antagonistic gradient on later segmentation. Note animal cells larger at the pole than at the equator. Small vegetative pole cells, irregular blastocoel. Treated from 2-cell stage for 8 hours first day and 7^ hours second day.
FIG. D. Inhibition at the animal pole with too great heat in adjuvant gradient. Note sharp line of demarcation between large and small animal pole cells and entire absence of blastocoel. Treated from 2-cell stage for 7 J hours, temperature of hot inflow 300 C. Section through centre of the egg. FIG. E. Effect of lateral gradient on early segmentation. Note complete cells on one side only (heated side). Treated from fertilisation for 6 hours.
FIG. F. Effect of lateral gradient on later segmentation. Note large animal cells on cooled side, small animal cells on heated side; also sharp line of demarcation between large and small animal cells and almost entire absence of blastocoel. Section approximately through centre of egg. Treated
from 2-cell stage for 5^ hours.
PLATE VI.
FIG. G. Effect of lateral gradient applied at dorsal lip stage and continued for 6 hours. Asymmetrical neurula showing greater growth on heated side. Transverse section through anterior region—neural fold on right side only.
FIG. H. Effect of lateral temperature gradient on later embryos. Section through tadpole treated for 9 hours in the tail bud stage (heated right side) and preserved and sectioned at the close of treatment. Note more advanced stage of development of optic vesicle on the right and folding of the retina. FIG. J. Showing asymmetrical eyes of a tadpole subjected to lateral temperature gradient for 9 hours in the tail bud stage and sectioned 19 days after treatment. Eye on the heated side (1) con-siderably larger than that on the cooled side (2).