Animal breeding is based on population genetics as most of the economically important traits have polygenetic nature. Molecular genetics provides valuable information which could contribute to the knowledge of genes underlying quantitative pro-duction traits (Czarnik et al., 2007; Matějíček et al., 2007). However, the effect of many polymorphous variants of candidate genes or potential genetic markers is not properly known yet.
Growth hormone (GH) directly or indirectly af-fects numerous aspects of animal lactation, growth and reproduction. It plays an important role dur-ing nutrient partitiondur-ing. A significant role of GH during lactation is demonstrated by the 10–15% increase in milk yield which occurs in dairy cows treated with recombinant bovine GH (Bauman, 1999). The best-known polymorphism of GH1 is
the leucine to valine substitution at position 127 in exon 5. Lucy et al. (1993) reported the differ-ent frequencies of GH1L and GH1V alleles in major dairy breeds, and the correlation of gene variants with estimates of milk production in cows.
Growth hormone exerts its influence on growth and metabolism by interaction with growth hor-mone receptor on the surface of the target cells. The changes in the functional regions of GHR can affect its binding capacity and signalling pathway, and therefore alter the activity of GH in the target tissues (Argetsinger and Carter-Su, 1996). Several polymor-phisms of bovine GHR were described by Aggrey et al. (1999) or Ge et al. (2000) but their association with meat production traits was insignificant.
The locus of acylCoA-diacylglycerol acyltrans-ferase1 (DGAT1) was identified as that underlying
Supported by the Ministry of Education, Youth and Sports of the Czech Republic (Project No. MSM 6007665806).
The relation of
GH1, GHR
and
DGAT1
polymorphisms
with estimated breeding values for milk production
traits of German Holstein sires
E. Hradecká
1, J. Čítek
1, L. Panicke
2, V. Řehout
1, L. Hanusová
11Agricultural Faculty, South Bohemian University in České Budějovice, Czech Republic
2FBN Dummerstorf, Germany
AbstrAct: We analysed the relations of estimated breeding values (EBV) of 315 German Holstein sires to their genotypes in growth hormone gene (GH1), growth hormone receptor gene (GHR) and acylCoA-diacylglycerol acyltransferase 1 (DGAT1). The strong relation of DGAT1 K232A to the estimated breeding values for milk production traits has been confirmed, when allele DGAT1K was connected with higher milk fat yield, milk fat and milk protein content, while allele DGAT1A increased milk yield and milk protein yield. The effect of DGAT1 genotype explained from 4.70% of variability of EBVs for fat yield to 31.90% of vari-ability of EBVs for fat content. The evaluation of GH1 127 Leu/Val and GHR 257 SNP polymorphisms did not reveal an association of their polymorphism with EBVs for milk production traits, except the EBVs of GHRG/GHRG homozygotes for fat yield, which were significantly lower. The effect of GH1 or GHR genotype explained only a negligible portion of variability of EBVs (R2 < 1.00% in most cases).
the quantitative trait locus for milk production pa-rameters in the centromeric region of bovine chro-mosome 14 (Coppieters et al., 1998). Grisart et al. (2002) described four polymorphisms, among them an ApA (Adenine p Adenine) to GpC (Guanine p Cytosine) dinucleotide substitution in exon 8, causing a lysine to alanine amino acid substitution (K232A). They found an extremely significant ef-fect of this mutation on daughter yield deviations (DYDs) for milk yield, protein yield, fat yield, pro-tein and fat content, which was lately confirmed by Spelman et al. (2002) or Čítek et al. (2007).
The aim of our paper was to analyse the relation of growth hormone gene (GH1), growth hormone receptor gene (GHR) and acylCoA-diacylglycerol acyltransferase1 (DGAT1) gene polymorphisms with breeding values of German Holstein sires for milk production traits.
MAteriAl And MetHods
A total of 315 German Holstein sires born in 1998 to 2001 were genotyped for 127 Leu/Val polymor-phism in GH1, 257 SNP (exon 10) in GHR loci, and K232A polymorphism in DGAT1. DNA was isolated from whole blood or from frozen sperm. Analyses were performed by PCR/RFLP techniques.
For GH1, the primer sequences were as in Mitra et al. (1995). An amplified fragment of DNA (223 bp) was digested by AluI overnight and digested frag-ments were visualised on 3.5% agarose gel stained with ethidium bromide.
For GHR, the primer sequences were as in Di
Stasio et al. (2005). An amplified fragment of DNA (342 bp) was digested by AluI.
For DGAT1, the primer sequences were as in
Winter et al. (2002). The K232A polymorphism was distinguished by a restriction fragment length poly-morphism using the restriction endonuclease CfrI.
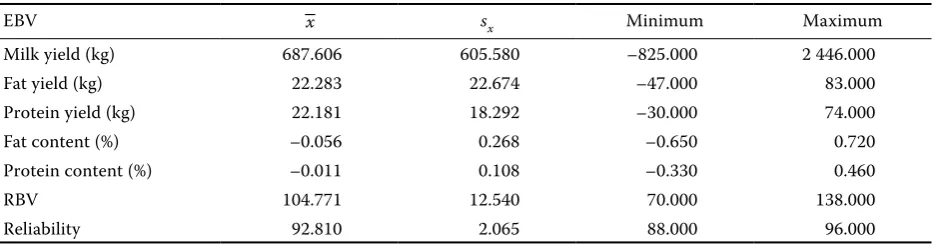
We analysed the relation between the detected genotypes and estimated breeding values (EBV) for milk production traits, i.e. 305 days milk yield (kg), fat content (%), fat yield (kg), protein content (%) and protein yield (kg) and the relative breed-ing value for milk production (RBVM). Estimated breeding values were provided by Vereinigte Informationssysteme Tierhaltung, Verden, Ger-many (EBVs of 2007/05). Generally, the use of daughter yield deviations (DYD) is preferred to EBVs, because EBVs contain information from other relatives than the bull’s daughters. However, the large number of daughters makes the difference between the DYD and EBV negligible (Viitala et al., 2003). As shown in Table 1, the reliabilities of EBVs were equable (on average 146 daughters per sire), which enables their direct comparison.
The analysis was based on this mixed model equation:
EBV = μ + Gi + eijk
where:
EBV = the estimated breeding value for partial milk produc-tion parameter
μ = the overall mean
Gi = the fixed effect of DGAT1, GH1 or GHR alleles and their combinations
eijk = the residual effect
We used the software STATISTICA (StatSoft, Inc., Tulsa, Oklahoma, USA) for evaluation, namely the ANOVA/MANOVA procedure with post-hoc com-parison of means (LSD test; P value). The Hardy-Weinberg equilibrium was tested by the χ2 test.
results
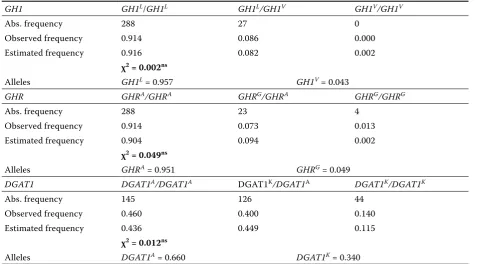
[image:2.595.65.539.636.758.2]The genotype frequencies at all loci are shown in Table 2. Predicted genotypic frequencies were
Table 1. Basic statistics of estimated breeding values of analysed sires
EBV –x sx Minimum Maximum
Milk yield (kg) 687.606 605.580 –825.000 2 446.000 Fat yield (kg) 22.283 22.674 –47.000 83.000 Protein yield (kg) 22.181 18.292 –30.000 74.000 Fat content (%) –0.056 0.268 –0.650 0.720 Protein content (%) –0.011 0.108 –0.330 0.460
RBV 104.771 12.540 70.000 138.000
similar to the observed ones (χ2 = 0.002 – 0.049) suggesting that genotype distributions were in the Hardy-Weinberg equilibrium.
GH1
The genetic analysis showed a very low occur-rence of allele V in German Holstein sires. The statistical analysis did not reveal a significant rela-tionship between EBVs and GH1 genotype, though the values tended to be higher in GH1L/GH1V het-erozygotes (Table 3). The effect of genotype/allele described only insignificant portions of variability, when the value of R2 did not exceed 1%. In cases
of EBVs for fat and protein content the variability within heterozygotes even exceeded total variability of the evaluated set. The frequency of heterozygous sires was too small and their variability too large to allow to draw a decisive conclusion.
GHR
A low frequency of GHRG allele and only 4 GHRG per GHRG homozygous sires (Table 2) were detect-ed. As shown in Table 4, the effect of GHR genotype on milk production EBVs was mostly insignificant, which is partly due to the extreme allelic frequen-cies and high variance within low-numbered
class-Table 2. Genotype and allelic frequencies of DGAT1, GH1, and GHR genes
GH1 GH1L/GH1L GH1L/GH1V GH1V/GH1V
Abs. frequency 288 27 0
Observed frequency 0.914 0.086 0.000 Estimated frequency 0.916 0.082 0.002
χ2 = 0.002ns
Alleles GH1L = 0.957 GH1V = 0.043
GHR GHRA/GHRA GHRG/GHRA GHRG/GHRG
Abs. frequency 288 23 4
Observed frequency 0.914 0.073 0.013 Estimated frequency 0.904 0.094 0.002
χ2 = 0.049ns
Alleles GHRA = 0.951 GHRG = 0.049
DGAT1 DGAT1A/DGAT1A DGAT1K/DGAT1A DGAT1K/DGAT1K
Abs. frequency 145 126 44
Observed frequency 0.460 0.400 0.140 Estimated frequency 0.436 0.449 0.115
χ2 = 0.012ns
Alleles DGAT1A = 0.660 DGAT1K = 0.340
[image:3.595.62.540.101.365.2]NS = differences are not significant
Table 3. Mean breeding values ±sx for milk production traits according to GH1 genotype
Genotype Milk (kg) Fat (kg) Protein (kg) Fat (%) Protein (%) RBVM
GH1L/GH1L 676.003 ± 593.173 21.771 ± 22.256 21.840 ± 17.859 –0.0564 ± 0.266 –0.011 ± 0.105 104.510 ± 12.211
GH1L/GH1V 811.370 ± 726.365 27.741 ± 26.600 25.815 ± 22.481 –0.0510 ± 0.297 –0.017 ± 0.132 107.556 ± 15.636
P < 0.267 0.191 0.281 0.923 0.763 0.228
R2 0.001 0.002 0.001 –0.003 –0.003 0.001
GH1L 682.065 ± 599.217 22.038 ± 22.458 22.018 ± 18.072 –0.056 ± 0.267 –0.011 ± 0.107 104.647 ± 12.375
GH1V 811.370 ± 726.365 27.741 ± 26.600 25.815 ± 22.481 –0.051 ± 0.297 –0.017 ± 0.132 107.557 ± 15.636
P < 0.277 0.201 0.291 0.924 0.768 0.238
[image:3.595.69.533.623.756.2]es. A post-hoc comparison showed significantly lower EBVs of GHR G/GHRG homozygotes for fat yield (P < 0.028) also with a tendency to lower EBVs for fat content (P < 0.085). This relation was not confirmed when the effect of alleles was tested. The portion of EBV variability explained by the effect of genotype/allele did not exceed 1.00%.
DGAT1
The frequency of DGAT1K allele in the analysed data set reached 0.340. Table 5 summarizes the evaluation of polymorphism effect on milk pro-duction traits. The effect of K232A substitution on almost all analysed traits was significant (P < 0.001) with the exception of relative breeding value (RBV) for milk. Breeding values of DGAT1A/DGATA ho-mozygotes compared to those of DGAT1K/DGATK were on average higher by 548 kg for milk yield and by 12.6 kg for protein yield, while they were lower by 15.4 kg for fat yield, by 0.45% for fat content and by 0.13% for protein content. The highest portion of variability was explained in EBVs for fat content (31.90% by genotype; 17.80% by allele).
Joint effect of DGAT1-GH1
Table 6 shows the results of the analysis of joint effect of DGAT1-GH1 genotypes. Though its effect on EBVs was always evaluated as statistically highly significant, this was mainly due to DGAT1 poly-morphism. GH1 caused no significant differences within the classes according to DGAT1, with the exception of EBV for protein content. The EBV of protein content of DGAT1K/DGAT1A-GH1L/GH1V heterozygotes was higher by 0.066% than the EBV of DGAT1K/DGAT1A-GH1L/GH1L sires (P < 0.042) and exceeded also the breeding values of DGAT1K per DGAT1K-GH1L/GH1L and DGAT1K/DGAT1K -GH1L/GH1V genotypes (insignificantly).
discussion
GH1
[image:4.595.65.534.101.248.2]The frequency of GH1V allele was low, which is generally reported in Holstein cattle (Lucy et al., 1993; Sorensen et al., 2002). As reported by Lucy Table 4. Mean breeding values ±sx for milk production traits according to GHR genotype
Genotype Milk (kg) Fat (kg) Protein (kg) Fat (%) Protein (%) RBVM
GHRA/GHRA 701.615 ± 599.452 22.403 ± 23.002 22.458 ± 18.064 –0.062 ± 0.262 –0.014 ± 0.104 104.927 ± 12.476
GHRG/GHRA 577.609 ± 657.801 25.826 ± 14.025 20.826 ± 20.198 0.046 ± 0.330 0.017 ± 0.149 104.696 ± 12.481
GHRG/GHRG 311.500 ± 730.528 –6.750 ± 22.984 10.000 ± 24.372 –0.220 ± 0.241 –0.008 ± 0.073 94.000 ± 16.432
P < 0.294 0.028 0.375 0.085 0.410 0.224
R2 0.003 0.016 0.000 0.009 0.000 0.003
GHRA 696.853 ± 601.172 22.534 ± 22.706 22.396 ± 18.119 –0.058 ± 0.265 –0.013 ± 0.106 104.918 ± 12.455
GHRG 509.935 ± 661.861 17.419 ± 21.444 18.032 ± 21.004 –0.023 ± 0.325 0.011 ± 0.133 101.936 ± 13.815
P < 0.092 0.221 0.195 0.482 0.235 0.197
R2 0.003 0.001 0.001 –0.001 0.001 0.001
Table 5. Mean breeding values ±sx for milk production traits according to DGAT1 genotype
Genotype Milk (kg) Fat (kg) Protein (kg) Fat (%) Protein (%) RBVM
DGAT1A/DGAT1 896.579 ± 549.536 17.566 ± 20.997 26.641 ± 17.465 –0.204 ± 0.182 –0.041 ± 0.115 106.483 ± 12.216
DGAT1K/DGAT1 565.405 ± 590.275 23.992 ± 22.144 19.889 ± 18.019 0.017 ± 0.235 0.009 ± 0.115 103.818 ± 12.592
DGAT1K/DGAT1K 348.886 ± 587.681 32.932 ± 25.550 14.046 ± 18.106 0.225 ± 0.288 0.027 ± 0.090 101.864 ± 12.901
P < 0.000 0.000 0.000 0.000 0.000 0.055
R2 0.110 0.047 0.055 0.319 0.063 0.012
DGAT1A 796.262 ± 581.149 19.512 ± 22.656 24.596 ± 17.864 –0.137 ± 0.224 –0.026 ± 0.106 105.676 ± 12.363
DGAT1K 479.369 ± 596.090 27.668 ± 23.891 17.486 ± 18.199 0.102 ± 0.275 0.017 ± 0.105 103.014 ± 12.696
P < 0.000 0.000 0.002 0.000 0.000 0.011
[image:4.595.68.531.612.759.2]Ta bl e 6. M ea n br ee di ng v al ue s ± sx fo r m ilk p ro du ct io n tra its a cc or di ng to jo in t g en ot yp es o f D G AT 1 a nd G H 1 G en ot yp e M ilk (k g) Fa t ( kg ) Pr ot ei n (k g) Fa t ( % ) Pr ot ei n (% ) RB V M D G AT 1 A/D G AT 1 A G H 1 L /G H 1 L n = 1 30 8 73 .1 31 A
bC ±
5 17 .5 17 17 .0 62 aB
c ±
1 9. 72 9 26 .3 69 A
B ±
1 6. 36 2 –0 .1 99 A BC
D ±
0 .1 82 –0 .0 36 A
BC ±
0 .0 97 1 06 .2 69 ± 1 1. 39 3 D G AT 1 A/D G AT 1 A G H 1 L/G H 1 V n = 1 5 1 09 9. 80 0 D
EF ±
7 67 .4 76 21 .9 33
d ±
3 0. 41 5 2 9. 00 0
C ±
2 5. 76 8 –0 .2 45 EF G
H ±
0 .1 83 –0 .0 89 D
EF ±
0 .1 03 1 08 .3 3 ± 18 .3 02 D G AT 1 K/D G AT 1 A G H 1 L/G H 1 L n = 1 16 5 75 .3 88 A D
g ±
5 96 .6 82 2 3. 39 7 ae
f ±
2 2. 40 7 1 9. 67 2
A ±
1 7. 95 0 0 .0 06 A EI
J ±
0 .2 34 0 .0 04 A D
g ±
0 .1 13 1 03 .6 03 ± 1 2. 55 6 D G AT 1 K/D G AT 1 A G H 1 L/G H 1 V n = 1 1 4 99 .1 82
bE ±
5 22 .9 12 2 8. 54 6 ± 12 .0 91 23 .3 64 ± 1 8. 88 5 0 .0 98
BF ±
0 .2 60 0. 07 0 BE
g ±
0 .1 11 1 06 .5 45 ± 1 2. 77 0 D G AT 1 K/D G AT 1 K G H 1 L/G H 1 L n = 4 1 32 7. 85 4 C
Fg ±
5 95 .6 55 3 2. 51 2
Be ±
2 5. 49 5 13 .3 42
BC ±
1 8. 47 2 0 .2 30 C G
I ±
0 .2 80 0 .0 27 C
F ±
0 .0 92 1 01 .3 90 ± 1 3. 32 3 D G AT 1 K/D G AT 1 K G H 1 L/G H 1 V n = 2 4 57 .0 00 ± 4 46 .8 91 5 5. 50 0 cd
f ±
16 .2 63 1 9. 00 0 ± 4. 24 3 0 .4 20 D H
J ± 0
et al. (1993), dairy breeds with larger mature size (Holstein) are supposed to have the highest fre-quency of GH1L, which also correlates with higher milk production. As shown by Čítek et al. (2005), the spread of both alleles in the German Holstein population was affected indirectly by selection for milk production in the past, resulting in a higher occurrence of allele GH1L.
Though the relation between the allele GH1L and the level of milk production was documented (Bauman, 1999; Panicke et al., 2001), it is not always expressed when analysing the association between the genotypes and estimated breeding values. Lucy et al. (1993) found similar estimated transmitting abilities (ETA) in Holstein bulls of different GH1 genotypes, while the milk production was significantly higher in GH1L/GH1L compared to GH1L/GH1V Holstein cows. Schlee et al. (1994) found no significant dif-ference between milk breeding values in Simmental bulls, but the effects on milk fat and protein con-tent approached significance with a higher percent-age in GH1L/GH1V heterozygotes. In Polish Friesian bulls and heifers Grochowska et al. (2001) reported significant differences between genotypes, when GH1L/GH1V heterozygotes revealed the highest val-ues for milk and protein yields, while GH1L/GH1L homozygotes were superior in fat yield.
Generally, the influence of GH1 polymorphism on milk production traits is not conclusive and is rather breed-specific (Lucy et al., 1993; Sorensen et al., 2002). The association analysis is limited by the lack of homozygous GH1V/GH1V animals, which disables a direct comparison of both homozygous variants. Similar breeding values of GH1L/GH1L and GH1L/GH1V animals may also be explained as the consequence of GH1L dominance over GH1V variant (Grochowska et al., 2001; Sorensen et al., 2002). Another reason could be the genetic background of the population where the bulls are tested/mated. The indirect recombination effect should also be taken into account. The differences between breeds suggest that there may exist further linked polymorphisms in regions that are involved in the gene regulation or other mutations in regions within or close to the gene that affect the expres-sion of the gene by changing the binding affinity of transcription factors (Lucy et al., 1993; Schlee et al., 1994; Sorensen et al., 2002).
GHR
Similar allelic frequencies were described in Finnish Ayrshire, where Viitala et al. (2006) found
0.870 GHRA and 0.130 GHRG. Ge et al. (2000) de-tected a somewhat higher frequency of GHRG al-lele (0.220) in Angus cattle, while Di Stasio et al. (2005) found almost equal allelic distribution in Piemontese (GHRA = 0.490; GHRG = 0.510).
The effect of 257 SNP was tested mostly in relation to meat production traits (Ge et al., 2000; Di Stasio et al., 2005). Viitala et al. (2006) tested the effects of four GHR exonic SNPs on milk production traits, but found no effect of 257 SNP on QTL variance. We found significantly lower EBVs for fat produc-tion in GHRG/GHRG homozygotes. Though, three of the four GHRG/GHRG homozygous bulls were si-multaneously DGATA/DGATA homozygotes, which probably affected the result of the analysis. The low frequency and high variability of GHRG/GHRG ho-mozygotes disable to find any significant trend.
DGAT1
As reported by Grisart et al. (2002), following the evolutionary conservation of lysine residue,
DGAT1K is more than likely the ancestral state while
DGAT1A corresponds to a more recently evolved
genotype was found to be significantly better in the main selection criterion. The reason may be seen in the moderate selection for fat yield coupled with selection for protein in the breeding programme, which could keep the frequencies stable; and also in the fact that the DGAT1 itself does not directly influence the protein synthesis. However, it will be interesting to monitor the frequencies as a certain tendency in the allele DGAT1A in the sires of dif-ferent age has been observed.
Our results correspond well with many other au-thors (Grisart et al., 2002; Weller et al., 2003; Kühn et al., 2004) who confirmed that DGAT1K allele was associated with decreased milk and protein yield while it increased fat yield, fat and protein content. As reported by Grisart et al. (2002), a simultaneous decrease in milk and protein yield and increase in fat yield are an interesting feature of DGAT1A to
DGAT1K substitution as there exists a positive
ge-netic correlation between these three yield traits. The response of DGAT1K/DGAT1A heterozygotes could be characterized as intermediate, which con-firms our previous findings (Čítek et al., 2007) and corresponds with the hypothesis of the effect of the locus being approximately codominant with intermediate heredity (Grisart et al., 2002).
conclusions
The analyses confirmed DGAT1 as a strong can-didate for application in marker-assisted selection, though additional mutation could attribute to its effect. We did not prove a significant effect of GH1 or GHR polymorphisms on EBVs for milk produc-tion traits with the excepproduc-tion of EBV for fat yield, which was lower in GHRG/GHRG homozygotes. We can expect an additional effect primarily in joint ap-plication with other markers. The effect would be not high as the extreme allelic frequencies were already achieved by past selection. However, the important role of growth hormone and growth hormone recep-tor in the lactation process is well known, thus their polymorphisms andinteraction with other traits should be the subject of further research.
references
Aggrey S.E., Yao J., Sabour M.P., Lin C.Y., Zadworny D., Hayes J.F., Kuhnlein U. (1999): Markers within the regulatory region of the growth hormone receptor gene
and their association with milk-related traits in Hol-steins. J. Hered., 90, 148–151.
Argetsinger L.S., Carter-Su C. (1996): Mechanism of sig-nalling by growth hormone receptor. Physiol. Rev., 76, 1089–1107.
Bauman D.E. (1999): Bovine somatotropin and lactation: from basic science to commercial application. Domest. Anim. Endocrinol., 17, 101–116.
Coppieters W., Riquet J., Arranz J. J., Berzi P., Cambisano N., Grisart B., Karim L., Marcq F., Moreau L., Nezer C., Simon P., Vanmanshoven P., Wagenaar D., Georges M. (1998): A QTL with major effect on milk yield and composition maps to bovine chromosome 14. Mamm. Genome, 9, 540–544.
Czarnik U., Galiński M., Pareek Ch.S., Zabolewicz T., Wielgosy-Groth Z. (2007): Study of an association be-tween SNP 775C>T within the bovine ITBG2 gene and milk performance traits in Black and White cows. Czech J. Anim. Sci., 52, 1–6.
Čítek J., Řehout V., Bláhová B., Pávková J., Panicke L. (2005): The genetic divergence among cattle breeds on candidate loci. FBN Dummerstorf, Schriftenreihe, 15, 43–48.
Čítek J., Řehout V., Hradecká E., Večerek L., Panicke L. (2007): The breeding values of German Holstein sires and the DGAT1 polymorphism. Arch. Tierz., 50, 136–146.
Di Stasio L., Destefanis G., Brugiapaglia A., Albera A., Rolando A. (2005): Polymorphism of the GHR gene in cattle and relationships with meat production and qual-ity. Anim. Genet., 36, 138–140.
Gautier M., Capitan A., Fritz S., Eggen A., Boichard D., Druet T. (2007): Characterization of the DGAT1 K232
and a variable number of tandem repeat polymorphisms in French dairy cattle. J. Dairy Sci., 90, 2980–2988. Ge W., Davis M.E., Hines H.C., Irvin K.M. (2000): Rapid
communication: Single nucleotide polymorphism de-tected in exon 10 of the bovine growth hormone recep-tor gene. J. Anim. Sci., 78, 2229–2230.
Grisart B., Coppieters W, Farnir F., Karim K., Ford C., Cambisano N., Mni M., Reid S., Spelman R., Georges M., Snell R. (2002): Positional candidate cloning of a
QTL in dairy cattle: identification of a missense mutation in the bovine DGAT1 gene with major effect on milk yield and composition. Genome Res., 12, 222–231. Grochowska R., Sorensen P., Zwierzchowski L.,
Sno-chowski M., Lovendahl P. (2001): Genetic variation in stimulated GH release and in IGF-I of young dairy cat-tle and their associations with the leucine/valine poly-morphism in the GH gene. J. Anim. Sci., 79, 450–476. Kühn C., Thaller G., Winter A., Bininda-Emonds O.R.P.,
R. (2004): Evidence for multiple alleles at the DGAT1
locus better explains a quantitative trait locus with major effect on milk fat content in cattle. Genetics, 167, 1873–1881.
Lucy M.C., Hauser S.D., Eppard P.J., Krivi G.G., Clark J.H., Bauman D.E., Collier R.J. (1993): Variants of so-matotropin in cattle: gene frequencies in major dairy breeds and associated milk production. Domest. Anim. Endocrinol., 10, 325–333.
Matějíček A., Matějíčková J., Němcová E., Jandurová O.M., Štípková M., Bouška J., Frelich J. (2007): Joint effects of CSN3 and LGB genotypes and their relation to breeding values of milk production parameters in Czech Fleckvieh. Czech J. Anim. Sci., 52, 83–87. Mitra A., Schlee P., Balakrishman C.R., Pirchner F. (1995):
Polymorphisms at growth-hormone and prolactin loci in Indian cattle and buffalo. J. Anim. Breed. Genet., 112, 71–74.
Panicke L., Čítek J., Schmidt M., Řehout V., Erhardt G., Staufenbiel R. (2001): Enzyme activities and milk per-formance by clusters of growth hormone gene (GH) and β-casein (β-cN) in German Holsteins. Czech J. Anim. Sci., 46, 202–208.
Schlee P., Graml R., Rottsmann O., Pirchner F. (1994): Influence of growth-hormone genotypes on breeding values of Simmental bulls. J. Anim. Breed. Genet., 111, 253–256.
Sorensen P., Grochowska R., Holm L., Henryoon M., Lovendahl P. (2002): Polymorphism in the bovine
growth hormone gene affects endocrine release in dairy calves. J. Dairy Sci., 85, 1887–1893.
Spelman R.J., Ford A.C., McElhinney P., Gregory G.C., Snell R.G. (2002): Characterization of the DGAT1 gene in the New Zealand dairy population. J. Dairy Sci., 85, 3514–3517.
Viitala S., Schulman N., de Koning D., Elo K., Kinos R., Virta A., Mäki-Tanila A., Vilkki J. (2003): Quantitative trait loci affecting milk production traits in Finnish Ayrshire dairy cattle. J. Dairy Sci., 86, 1828–1836. Viitala S., Szyda J., Blott S., Schulman N., Lidauer M.,
Mäki-Tanila A., Georges M., Vilkki J. (2006): The role of the bovine growth hormone receptor and prolactin receptor genes in milk, fat and protein production in Finnish Ayrshire dairy cattle. Genetics, 173, 2151–2164. Weller J.I., Golik M., Seroussi E., Ezra E., Ron M. (2003):
Population-wide analysis of a QTL affecting milk-fat production in the Israeli Holstein population. J. Dairy Sci., 86, 2219–2227.
Winter A., Krämer W., Werner F.A.O., Kollers S., Kata S., Durstewitz G., Buitkamp J., Womack J.E., Thaller G., Fries R. (2002): Association of lysine232 alanine polymorphism in bovine gene encoding acylCoA: dia-cylglycerol acyltransferase (DGAT1) with variation at a quantitative trait locus for milk fat content. PNAS, 99, 9300–9305.
Received: 2007–09–13 Accepted after corrections: 2008–03–11
Corresponding Author
Ing. Eva Hradecká, Ph.D., Agricultural Faculty, University of South Bohemia in České Budějovice, Studentská 13, 370 05 České Budějovice, Czech Republic