Emily Williams
A thesis submitted to the faculty of the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Master of Arts in the Department of
Psychology and Neuroscience (Behavioral and Integrative Neuroscience)
Chapel Hill 2019
Approved by:
Kathryn J. Reissner
Regina M. Carelli
Todd E. Thiele
ABSTRACT
Emily Williams: The Effects of Nicotinamide on Reinstatement to Cocaine Seeking (Under the direction of Kathryn Reissner)
ACKNOWLEDGEMENTS
TABLE OF CONTENTS
LIST OF FIGURES...vi
LIST OF ABREVIATIONS...vii
Chapter I INTRODUCTION... 1
Oxidative Stress... 1
NAD+ signaling and mitochondrial functioning... 3
II METHODS... 5
Subjects and Reagents... 5
Surgical Procedures... 5
Behavioral Training... 6
Experiment 1 ... 7
Experiment 2 ... 8
Experiment 3 ... 8
Experiment 4 ... 8
Experiment 5 ... 9
III RESULTS... 11
Experiment 1 ... 11
Experiment 2 ... 12
Experiment 3 ... 13
Experiment 4 ... 13
Experiment 5 ... 14
LIST OF FIGURES
Figure
1 Chronic NAM administration reduces cue- but not cocaine-primed reinstatement in males, but not females... 23
2 The effects of chronic NAM administration are specific to cue-primed reinstatement 25
3 Acute NAM administration did not affect cue- or cocaine-primed reinstatement ... 26
8-OHG 8-hydroxyguanine ATP adenosine triphosphate BLA basolateral amygdala
CeA central nucleus of the amygdala CPP conditioned place preference
DA dopamine
Dnmt1 DNA methyltransferase FR fixed ratio
IP intraperitoneal injection IV intravenous infusion NAc nucleus accumbens
NAD+ nicotinamide adenine dinucleotide NAM nicotinamide
NADP+ nicotinamide adenine dinucleotide phosphate NAMPT nicotinamide phosphoribosyltransferase NMN nicotinamide mononucleotide
OS oxidative stress PAR poly(ADP-ribose)
PARP-1 poly(ADP-ribose) polymerase ROS reactive oxygen species SAL saline
CHAPTER 1: INTRODUCTION
Nicotinamide (NAM), a form of vitamin B3, component of crucial coenzymes NAD+ and NADP+, and free radical scavenger, is widely used clinically to treat pellagra (niacin deficiency), bullous pemphigoid, acne and rosacea, atopic dermatitis, and skin photoageing (Mutasim, 2003; Surjana & Damian, 2011). It also shows promise in preserving and enhancing neurocognitive function in a variety disease models such as Alzheimer’s, traumatic brain injury (ischemia and contusion injury), and Parkinson’s disease (Rennie, Chen, Dhillon, Vardy, & Damian, 2015). In addition, NAM has been proposed as an effective treatment for substance abuse (Namazi, 2004; Otte, Borelli, & Korting, 2005); however, empirical scientific evidence for this is lacking. Nevertheless, several cellular functions of NAM indicate that it may be beneficial in opposing cellular adaptations that mediate cocaine seeking in a preclinical rodent model of addiction, including consequences of oxidative stress and compromised cellular metabolic functions.
Oxidative Stress
Importantly, levels of ROS have been linked to cocaine seeking, as administration of ROS scavengers leads to a reduction in cocaine self-administration as well as attenuation of cocaine-induced enhancement of dopamine (DA) release (Jang et al., 2015). In addition, in comparison to animals who were assigned to a cocaine yoke procedure, animals who voluntarily self-administer cocaine show significantly altered expression of oxidative stress biomarkers in the hippocampus, frontal cortex, and dorsal striatum (Pomierny-Chamiolo et al., 2013).
NAD+ signaling and mitochondrial functioning
In addition, chronic cocaine also impairs both NAD+ signaling and mitochondrial function (de Oliveira & Jardim, 2016; Logan et al., 2018). NAD+ is important for energy homeostasis and calcium transport in the brain (Liang et al., 2017; Tamsett, Picchione, & Bhattacharjee, 2009; Ying, 2007) and is depleted following DA stimulation (Requardt et al., 2012; Requardt, Wilhelm, Rillich, Winkler, & Hirrlinger, 2010). Within neuronal mitochondria isolated from 9-12 week old rats, cocaine impairs mitochondrial respiration (Cunha-Oliveira et al., 2013), and disrupts the mitochondrial membrane potential of mitochondria in cultured neurons (Cunha-Oliveira et al., 2006; Yao, Allen, Zhu, Callen, & Buch, 2009). Cocaine exposure also results in in disrupted mitochondrial membrane potential within the mitochondria of cultured astrocytes (Yang, Yao, Lu, Wang, & Buch, 2010). As mentioned previously, NAM serves as a means to increase the bioavailability of NAD+ and other coenzymes crucial to mitochondrial functioning (Fricker, Green, Jenkins, & Griffin, 2018). Given that the many mitochondrial processes depend both on NAD+ and its phosphorylated counterpart NADP, the availability and redox state of NAD+ has important influence on all major mitochondrial processes (Dolle, Rack, & Ziegler, 2013). As a precursor to NAD+, NAM administration may therefore combat cocaine dependent disruptions in NAD+ signaling and mitochondrial functioning.
CHAPTER 2: METHODS AND MATERIALS Subjects and reagents
Male (225-250g upon arrival, Envigo) and female (175-200g upon arrival, Envigo) Sprague-Dawley rats, were housed individually in a temperature-controlled environment (20-21° C) on a 12-h reverse light cycle (7 am off, 7 pm on). Prior to the start of experimental procedures, rats were maintained on ad libitum food and were given unlimited water access throughout the experiment. Treatment and housing of animals was approved by the University of North Carolina Institutional Animal Care and Use Committee and followed the “Guide for the Care and Use of Laboratory Rats” (Institute of Laboratory Animal Resources on Life Sciences, National Research Council, 1996). Cocaine was generously provided by the NIH Drug Supply Program. Nicotinamide was purchased from Sigma-Aldrich (catalog #72340) and prepared in saline.
Surgical Procedures
(100U/mL) were administered i.v. To check for catheter patency, propofol (1 mg/0.1 mL, i.v.) was administered prior to the onset of self-administration and subsequently as needed.
Behavioral Training
Before surgery, rats were trained to lever press for food pellets (45 mg pellets; Bio Serv, Flemington, NJ) under an FR1 reinforcement schedule. Food training and self-administration sessions were performed in sound attenuated, standard operant conditioning chambers (Med Associates, St. Albans, VT). Each chamber was equipped with two retractable levers divided by a food pellet dispenser. Starting 24 hours prior to food training session, rats received rations of approximately 20g of chow daily. Lever presses on the active lever during training sessions resulting in the delivery of one food pellet (Bio Serv, Flemington, NJ). Presses on the inactive lever had no consequences for the entirety of the training session. Training session continued until animals reached a criterion of 100 active lever presses or 8 hours had elapsed.
Experiment 1: Effects of Chronic NAM administration on cue- and cocaine-primed reinstatement in males and females
A timeline for the experiment can be seen in Figure 1A. Given the relatively short half-life of NAM (100 mg/kg: 3.4 hours), rats were given daily i.p. injections of NAM (10 mg/kg, 30 mg/kg, 60 mg/kg, or 120 mg/kg; 1 mL/kg) or saline 30 minutes prior to every extinction session and subsequent reinstatement tests (Stratford & Dennis, 1994). Twenty-four hours following the last extinction session, rats were tested on a 2h cue-primed reinstatement session. This session began with a five second presentation of the light and tone cues which had been previously presented during self-administration. All subsequent active lever presses resulted in identical light and tone cue presentations without delivery of cocaine. Inactive lever presses were recorded, but maintained a lack of programmed consequences. Cue-primed reinstatement was followed by a return to extinction sessions for three days to return to baseline extinction pressing. Following the third extinction session, animals were tested on a cocaine-primed reinstatement test. For this reinstatement session, animals received NAM or saline i.p. 30 minutes prior to the session, just as in extinction and cue-primed reinstatement testing. However, immediately before entering the chambers, animals were given a 10 mg/kg i.p. injection of cocaine. Active and inactive lever presses during cocaine-primed reinstatement did not result in the delivery of light or tone cues.
type identification was made microscopically consulting methods by Cora, Kooistra, and Travlos (2015); Zenclussen, Casalis, Jensen, Woidacki, and Zenclussen (2014)
Experiment 2: Effects of Chronic NAM administration on locomotion
To examine if the effects of effects on responding were due to locomotor sedative effects, open field locomotor activity was assessed in the same cohort of rats used for reinstatement testing. Prior to locomotor testing, animals remained in their home cages 1-5 days following the final reinstatement procedure; injections of NAM or saline were given daily. On the day of locomotor testing, rats were given NAM or saline injections 30 minutes before being placed into standard locomotor chambers (Med Associates, St. Albans, VT). Locomotor behavior was recorded for a total of 2h. At the one-hour time point, all animals were given a 10 mg/kg i.p. injection of cocaine and returned to the chamber to continue locomotor recording for the second hour.
Experiment 3: Effect of chronic NAM administration on cocaine-primed reinstatement alone
In order to control for any possible ordering effects of the reinstatement test sessions, a separate cohort of rats was given only a cocaine-primed reinstatement session following extinction. All other procedures were identical to experiment 1. A timeline for the experiment can be seen in Figure 2A
Experiment 4: Effects of Acute NAM administration on cue- and cocaine-primed reinstatement
Experiment 5: Effects of Chronic NAM administration on Food Reinstatement
In an effort to determine whether or not the effects of NAM administration on reinstatement are specific to drug reinforcers, a cohort of animals completed a food self-administration, extinction, and reinstatement paradigm as previously described by Cosme, Gutman, and LaLumiere (2015) and McFarland and Kalivas (2001). Briefly described, animals initially pressed on the active lever for an FR1 schedule for delivery of a 45 mg food pellet, followed progressively by sessions on FR3 and FR5 reinforcement schedules. Each rat was required to press for at least 100 food pellets per day for at least three days in order to progress to the next schedule of reinforcement. Completion of each reinforcement schedule also resulted in the presentation of both the light and tone cue for 5 seconds. After these criteria were reached on the FR5 schedule, extinction procedures began. As for cocaine studies, NAM (120 mg/kg) or saline injections began on day 1 of extinction and continued for the remaining duration of the experimental sessions. For food-primed reinstatement, two food pellets were placed in the food hoppers before the session began. During the first 30 minutes of the session, a single food pellet was delivered non-contingently every two minutes; the remaining time proceeded as a typical extinction session. Presses on either lever had no programmed consequences for the entire duration of the session. An experimental timeline can be seen in Figure 4A.
Data Analysis
compare active lever presses on the last day of extinction and active lever pressing on the tests of reinstatement of NAM and SAL animals. Sidak's multiple comparisons test was used to examine whether each group or dose displayed an increase in active lever pressing in comparison to the final day of extinction. For locomotor tests, two-way repeated measures ANOVA was performed with time (5 minute bins) and group (NAM: 10 mg/kg, 30 mg/kg, 60 mg/kg, 120 mg/kg or SAL) as factors. Tukey’s multiple comparisons test was used to examine whether locomotor behavior increased in all groups due to administration of cocaine at the 1 hour time-point. One-way ANOVAs were performed to examine differences between groups in the total distance traveled (cm) for the first hour and second hour. Data shown are SEM and significance was considered p
CHAPTER 3: RESULTS
Experiment 1: Chronic NAM administration reduces cue- but not cocaine-primed reinstatement in males but not females
group also pressed significantly less than the 10 mg/kg group (p < 0.05). Following cue-primed reinstatement, rats underwent three additional extinction sessions, followed by a cocaine-primed reinstatement test. No effect of NAM treatment was observed on subsequent cocaine-primed reinstatement (Fig. 1D; F(4, 49) = 0.331, p = 0.856).
Experiment 2: Chronic NAM administration has no effects on open-field locomotor or cocaine-primed locomotor
The same cohort of male animals from experiment 1 were tested on open field and cocaine-induced locomotor 30 minutes following a NAM injection; results are shown in Figure 1H-J. There were no differences between groups at any time point during the entire 120 minutes (Fig. 1H, F (4,49) = 1.045, p = 0.394). There was no interaction (F(92, 1127) = 1.07, p = 0.312) and no main effect of group (F(4, 49) = 1.045, p = 0.394) but there was a significant main effect of time (F(23, 1127) = 116.4, p <0.0001). Tukey’s multiple comparison tests revealed that for all groups distance traveled (cm) increased following an injection of cocaine (p < 0.0001). There were no differences between groups in total distance traveled during the first 60 minutes of open field testing (Fig. 1I, F(4,49) = 1.886, p = 0.128) or during the final 60 minutes following experimenter administration of cocaine (Fig. 1J, F(4, 49) = 0.981, p = 0.427).
Experiment 3: Chronic NAM administration had no effect on cocaine-primed reinstatement.
SAL p = 0.05, NAM p < 0.05); however, treatment groups did not differ on active lever pressing during cocaine-primed reinstatement (Fig. 2C, F(1,11) = 0.134, p = 0.721).
Experiment 4: Acute NAM administration does not affect cue- or cocaine-primed reinstatement
Self-administration and extinction data for experiment 4 are shown in Figure 3B. Prior to receiving any NAM or saline, neither lever pressing behavior nor reinforcers earned differed across future groups (active lever pressing: (F(1, 144) = 0.543; p = 0.462); reinforcers earned: (F(1,144) = 0.695; p = 0.406). In contrast to chronic administration of NAM across extinction, there was no effect of acute 120 mg/kg NAM on active lever responding during cue-primed reinstatement (Fig. 3C, effect of reinstatement: F(1,12) = 39.93, p < 0.0001 SAL p < 0.005, NAM p < 0.005; F(1,12) = 0.092, p = 0.766). Likewise, there was no effect of acute 120 mg/kg NAM on cocaine-primed reinstatement (Fig. 3D, effect of reinstatement: F(1,12) = 26.87, p < 0.001, SAL p < 0.01, NAM p < 0.01; F(1,12) = 0.1213, p = 0.7337).
Experiment 5: Chronic NAM administration did not reduce cue or food-primed reinstatement
CHAPTER 4: DISCUSSION Discussion
Results of the current study indicate that chronic, but not acute, systemic administration of NAM specifically reduces responding in cue-primed, but not cocaine-primed reinstatement in male rats but not female rats. This effect on cue-primed reinstatement is likely not due to a generalized sedative effect, motor effect, or generalized effect on motivation, as both locomotor activity, cocaine-primed reinstatement, and food reinstatement were unaffected.
Therapeutic Uses of Nicotinamide
although NAM has been suggested by some to be a potential “anti-addiction weapon,”(Namazi, 2004), to our knowledge, there are no existing empirical scientific studies that examine NAM treatment in the context of drug self-administration, extinction, and reinstatement.
Cocaine Cue-primed Reinstatement Specificity
Our results suggest that the effects of NAM are specific to reducing cue-primed reinstatement. Examination of neural circuitry involved in different modalities of reinstatement unsurprisingly reflects that different stimuli engage overlapping yet distinct circuits. Of particular interest in regard to cue-primed reinstatement are the basolateral amygdala (BLA) and the central amygdala (CeA), as inactivation of these areas reduces cue-primed reinstatement (Feltenstein & See, 2008; See, 2005) but not cocaine-primed reinstatement (Grimm & See, 2000; McFarland & Kalivas, 2001). Though none have examined the effects of NAM, PARP-1 inhibition, or NAD+ supplementation in a reinstatement model, PARP-1 inhibition in the CeA but not the BLA reduces cocaine conditioned place preference (CPP) (Lax et al., 2017). Additionally, Lax et al. (2017) found that while PARP-1 was significantly activated in the CeA, it was not significantly increased in the BLA further implicating the potential specificity of PARP-1 effects in the CeA. The differential circuitry of cue-primed and cocaine-primed reinstatement suggests that the effects of NAM may be primarily influential within the amygdala. The importance and mechanistic effects of NAM within the CeA is yet to be determined and requires further study within a reinstatement model.
lack of effect of NAM any locomotor activity, this further confirms the lack of any generalized motor or motivational effect of chronic NAM.
Nicotinamide and Sex differences
While NAM dose-dependently reduced cue-primed reinstatement in male rats, it was without effect in females. In order to investigate potential avenues of explanation for the resulting sex differences in NAM treatment, consulting studies relating to ischemia treatment are informative.
Sex differences have been reported in the effects of both NAM administration and PARP-1 inhibition as treatments for ischemia as well as oxidative stress. For example, Siegel and McCullough (2013) found that NAM treatment was effective in reducing oxidative stress induced damage in male mice, but showed no effects in females. Further, when PARP-1 knockout animals were tested, they found that the oxidative stress induced damage was significantly greater in PARP-1 knockout females while it was decreased in knockout males. However, in both PARP-1 knockout males and knockout females, treatment with NAM reduced the oxidative stress induced damage. Similar exacerbations of ischemic damage occur in female neonatal, adult, and aging female rats when PARP-1 is disrupted or inhibited (Hagberg et al., 2004; McCullough, Zeng, Blizzard, Debchoudhury, & Hurn, 2005) These results are similar to ours in such that our NAM treatment reduced cue-reinstatement in males, but had no effect in females indicating that the mechanism by which NAM reduces cue-primed reinstatement is sex dependent.
ischemic injury (Siegel & McCullough, 2013). This could be due to the fact that females have higher NADH-linked respiration rates in brain mitochondria as NAD+ is reduced to NADH during this respiration (Gaignard et al., 2015). These differences suggest that mediation of PARP-1 or NAD+ may not be a viable treatment option in females; however, others did report that female rats’ infraction volume following ischemic injury was decreased similarly to male rats at the same dose and application method of NAM (Sakakibara, Mitha, Ogilvy, & Maynard, 2000). More research is needed regarding NAM as a viable application in females to reduce reinstatement responding. For example, future studies should investigate a variety of NAM doses in females to determine if the lack of effects on reinstatement could be related to a difference in effective dose.
Mechanism of Action
Our results are limited to the behavioral effects of NAM. Based upon the broader literature, several functions of NAM could be at play in exerting the effect on cue-primed reinstatement, including: PARP-1 inhibition, NAD+ supplementation, and oxidative stress reduction. There is, however, some disagreement in the literature surrounding NAM’s function as a PARP inhibitor. Specifically, it has been argued that NAM’s ability to inhibit PARP-1 may only exist in in vitro testing and not in vivo (Kirkland, 2009). On the other hand, it has been suggested that NAM’s function may be dependent upon dosing such that only high doses of NAM lead to PARP-1 inhibition (5mM or more) while lower doses result in PARP-1 facilitation through provision of NAD+ (Riklis et al., 1990). Facilitation through NAD+ is thought to be beneficial due to the fact that PARP-1 activation is followed by NAD+ depletion and a resulting ATP depletion due to inhibition of glycolysis (Andrabi et al., 2014; Ying, Garnier, & Swanson, 2003) in cells including astrocytes (Suh et al., 2007; Tang et al., 2010) and overall brain tissue (Cosi & Marien, 1999; Zeng et al., 2007). PARP-1 activation can also lead to mitochondria dysfunction, cell death (David, Andrabi, Dawson, & Dawson, 2009; Fatokun, Dawson, & Dawson, 2014), and inhibition of astrocytic glutamate uptake capacity (Tang et al., 2010) all of which can be prevented by supplicating with NAD+ (Alano, Ying, & Swanson, 2004; Liu, Pitta, & Mattson, 2008; Tang et al., 2010).
nicotinamide mononucleotide (NMN) which can then be converted to NAD+ (Dahl, Holm, Aukrust, & Halvorsen, 2012), it seems less likely that an increase of NAD+ would decrease cocaine seeking. Although an increase in NAMPT expression reduces ischemic injury in neurons (P. Wang et al., 2011), overexpression of NAMPT in the ventral tegmental area (VTA) of mice prior to cocaine CPP results in facilitation of cocaine-induced CPP (Kong et al., 2018). On the other hand, inhibition of VTA NAMPT prior to each CPP training session decreases cocaine-induced CPP and furthermore, the relative VTA content of NAD following CPP correlates with observed CPP scores (Kong et al., 2018). Interestingly, since NAMPT functions to convert NAM to NMN, inhibition of NAMPT would hypothetically result in an increase of NAM which could be working by unknown mechanisms to reduce cocaine CPP in this situation. The effect of NAMPT manipulation on cocaine CPP suggests that a proposed mechanism focusing on NAM’s role as an NAD+ precursor would not reduce reinstatement, but would rather facilitate reinstatement. Conversely, both deletion of PARP-1 as well as direct inhibition of PARP-1 is linked to increases the cellular content of NAD+ (Bai et al., 2011; Mukhopadhyay et al., 2017), which suggests that NAM’s actions as a PARP-1 inhibitor may be inseparable from its ability to increase NAD+ availability. To our knowledge, however, this has not been directly tested; therefore, future studies may investigate whether direct PARP-1 inhibition or supplementation with NAD+ is sufficient for attenuating reinstatement as well as whether or not the effects of chronic NAM administration are dependent upon either of these mechanisms.
PARP-1 and gene regulation
Knackstedt, Melendez, & Kalivas, 2010). The reduction in mRNA and protein expression of GLT-1 is associated with increased methylation of the GLT-1 gene, Slcla2 (Kim et al., 2018). PARP-1 has many varied roles in gene regulation including DNA methylation (Kraus & Hottiger, 2013). More specifically, PARP-1 regulates the expression and activity of the DNA methyltransferase Dnmt1 (Caiafa, Guastafierro, & Zampieri, 2009). While the activity of PARP-1 has not been directly linked to Slcla2 expression, PARP-PARP-1 activation is known to impair glutamate uptake activity in cultured astrocytes (Tang et al., 2010)Likewise, interaction of PAR with Dnmt1 following cocaine self-administration has not been examined. Therefore, future studies can examine how PARP-1 activity may change across time and between different self-administration models. In addition, future studies may examine the regulatory nature of PARP-1 on Dmnt1 across time and between different self-administration models.
B
0 5 10
0 50 100 150 Lever Presses 60 mg/kg 120 mg/kg Saline 30 mg/kg Active Inactive Self-Administration Extinction
5 10 15
10 mg/kg Active Lever Presses
*
A
Self-administration ExtinctionCue Coc Loco
12d 14-17d 3d 1-5d
NAM or saline
H
0 10 20 30 40 50 60 70 80 90 100 110 120 0 500 1000 1500 2000 2500 Time (min)
Ambulatory Distance (cm)
10 mg/kg 30 mg/kg 60 mg/kg 120 mg/kg Saline Cocaine
SAL 10 30 60 120
0 1000 2000 3000 4000 5000 Dose (mg/kg) To ta l D is ta n ce ( cm ) Open Field
(t = 0 - 60)
11 10 11 11 12
SAL 10 30 60 120 0 5000 10000 15000 20000 Dose (mg/kg) To ta l D is ta n ce ( cm ) Cocaine-Induced
(t = 60 - 120)
11 10 11 11 12
SAL NAM
0 50 100
Active Lever Presses
Cue-primed 13 12 SAL NAM 0 50 100
Active Lever Presses
Cocaine-primed
13 12
SAL 10 30 60 120 0
20 40 60 80
Active Lever Presses
Cocaine-primed
Dose (mg/kg) 11 10 11 11 12
I
J
G
0 5 10
0 50 100 150
Lever Presses
5 10 15
Self-Administration Extinction NAM Saline Active Inactive *
Active Lever Presses
E
SAL 10 30 60 120
0 20 40 60 80 100
Active Lever Presses
Cue-primed
Dose (mg/kg) 11 10 11 11 12
* *
D
C
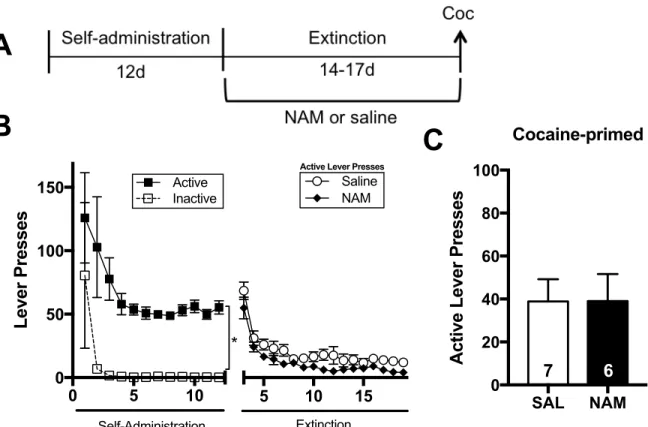
Figure 1: Chronic NAM administration reduces cue- but not cocaine-primed reinstatement in males, but not females. (A) Experimental Timeline. (B) Active and inactive lever presses across days during self-administration in male rats (left). Active lever presses between treatment groups are shown, right. No differences in active lever presses among treatment groups were seen across extinction. (C) Males who received daily i.p. 120 mg/kg and 60 mg/kg NAM injections throughout extinction pressed the active lever significantly less during a test of cue-primed reinstatement than those who received daily saline injections throughout extinction (p <0.05). Additionally, those in the 120 mg/kg group also pressed significantly less than those in the 10 mg/kg group (p < 0.05). (D) Males treated with 0-120 mg/kg NAM during extinction did not differ significantly when tested with a cocaine-primed reinstatement session. (E) Active and inactive lever presses across days during self-administration in female rats. A main effect of lever was observed between active and inactive lever presses (* p < 0.05). No differences in active lever presses among treatment groups were seen across extinction. (F) Females that received 120 mg/kg NAM did not differ significantly during a tests of cue-primed or (G) cocaine-primed reinstatement. (H) Ambulatory distance (cm) was measured beginning 30 minutes after a saline or NAM i.p. injection and continued for two hours. Arrow indicates 10 mg/kg i.p. cocaine injection given at the 60 minute mark. (I) No significant differences were seen between groups during the open field behavior of the first hour (t = 0-60). (J) No significant differences were seen between groups following the cocaine injection during the second hour (t = 60-120). Data shown in H-J are from the same male rats as in B-D. * indicates significant difference between active and inactive lever pressing (B) and between treatment groups by post-hoc analysis (C).
Figure 2: The effects of chronic NAM administration are specific to cue-primed reinstatement. (A) Experimental timeline. (B) Lever pressing across self-administration and extinction. A main effect of lever was observed between active and inactive lever presses (* p < 0.05). (C) No significant differences between groups were seen during the cocaine-primed reinstatement.
Figure 2:
A
SAL NAM 0 20 40 60 80 100Active Lever Presses
Cocaine-primed
6
7
C
B
0 5 10
0 50 100 150 Lever Presses Inactive Active NAM Saline
5 10 15
Self-Administration Extinction
Active Lever Presses
Figure 3: Acute NAM administration did not affect cue- or cocaine-primed reinstatement.
(A) Experimental timeline. (B) Active and inactive lever presses across self-administration and extinction. A main effect of lever was observed between active and inactive lever presses (* p < 0.05). (C) No significant differences between groups were seen during a cue-primed reinstatement test after receiving 120 mg/kg injections of NAM thirty minutes prior to the test session. (D) No significant differences between groups were seen during the cocaine-primed reinstatement after receiving 120 mg/kg injections of NAM 30 minutes prior to the test session.
A
Figure: 3
SAL NAM 0 50 100Active Lever Presses
Cue-primed 7 7 SAL NAM 0 50 100
Active Lever Presses
Cocaine-primed 7 7
D
5 10 0 50 100 150 Lever Presses Inactive Active5 10 15
Self-Administration Extinction *
Figure: 4
A
B
C
D
SAL NAM 0 20 40 60 80
Active Lever Presses
Cue-primed 8 8 SAL NAM 0 50 100 150 200
Active Lever Presses
Food-primed
8 8
Figure 4: Chronic NAM administration did not affect cue- or food-primed reinstatement. (A) Experimental timeline. (B) Animals reliably self-administered food and extinguished responding reliably during extinction. A main effect of lever was observed between active and inactive lever presses (* p < 0.05). There was a significant interaction between group and session during extinction. Multiple comparisons reveled that the NAM group had significantly fewer active lever presses on day 1 of extinction than the saline group (* p <0.0001). Active lever pressing the remaining days was not significantly different between groups (p > 0.9999). (C) No significant differences between groups were seen during a cue-primed reinstatement test after receiving 120 mg/kg injections of NAM 30 minutes prior to the test session. (D) No significant differences between groups were seen during the food-primed reinstatement after receiving 120 mg/kg injections of NAM 30 minutes prior to the test session.
1 3 5 7 9 11
0 500 1000 1500 Lever Presses Self-Administration Extinction NAM Saline Inactive
Active Active Lever Presses
1 3 5 7 9 11 13 15 *
REFERENCES
Alano, C. C., Ying, W., & Swanson, R. A. (2004). Poly(ADP-ribose) polymerase-1-mediated cell death in astrocytes requires NAD+ depletion and mitochondrial permeability transition. J Biol Chem, 279(18), 18895-18902. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/14960594. doi:10.1074/jbc.M313329200
Alvaro-Bartolome, M., La Harpe, R., Callado, L. F., Meana, J. J., & Garcia-Sevilla, J. A. (2011). Molecular adaptations of apoptotic pathways and signaling partners in the cerebral cortex of human cocaine addicts and cocaine-treated rats. Neuroscience, 196, 1-15. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/21925237.
doi:10.1016/j.neuroscience.2011.08.074
Andrabi, S. A., Umanah, G. K., Chang, C., Stevens, D. A., Karuppagounder, S. S., Gagne, J. P., . . . Dawson, T. M. (2014). Poly(ADP-ribose) polymerase-dependent energy depletion occurs through inhibition of glycolysis. Proc Natl Acad Sci U S A, 111(28), 10209-10214. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/24987120. doi:10.1073/pnas.1405158111
Bai, P., Canto, C., Oudart, H., Brunyanszki, A., Cen, Y., Thomas, C., . . . Auwerx, J. (2011). PARP-1 inhibition increases mitochondrial metabolism through SIRT1 activation. Cell Metab, 13(4), 461-468. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/21459330. doi:10.1016/j.cmet.2011.03.004
Bayrakdar, E. T., Armagan, G., Uyanikgil, Y., Kanit, L., Koylu, E., & Yalcin, A. (2014). Ex vivo protective effects of nicotinamide and 3-aminobenzamide on rat synaptosomes treated with Abeta(1-42). Cell Biochem Funct, 32(7), 557-564. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/25111857. doi:10.1002/cbf.3049
Caiafa, P., Guastafierro, T., & Zampieri, M. (2009). Epigenetics: poly(ADP-ribosyl)ation of PARP-1 regulates genomic methylation patterns. FASEB J, 23(3), 672-678. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/19001527. doi:10.1096/fj.08-123265
Chouinard, G., Young, S. N., Annable, L., & Sourkes, T. L. (1977). Tryptophan-nicotinamide combination in depression. Lancet, 1(8005), 249. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/64768.
Chouinard, G., Young, S. N., Annable, L., Sourkes, T. L., & Kiriakos, R. Z. (1978). Tryptophan-nicotinamide combination in the treatment of newly admitted depressed patients.
Cora, M. C., Kooistra, L., & Travlos, G. (2015). Vaginal Cytology of the Laboratory Rat and Mouse: Review and Criteria for the Staging of the Estrous Cycle Using Stained Vaginal Smears. Toxicol Pathol, 43(6), 776-793. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/25739587. doi:10.1177/0192623315570339
Cosi, C., & Marien, M. (1999). Implication of poly (ADP-ribose) polymerase (PARP) in neurodegeneration and brain energy metabolism. Decreases in mouse brain NAD+ and ATP caused by MPTP are prevented by the PARP inhibitor benzamide. Ann N Y Acad Sci, 890, 227-239. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/10668429.
Cosme, C. V., Gutman, A. L., & LaLumiere, R. T. (2015). The Dorsal Agranular Insular Cortex Regulates the Cued Reinstatement of Cocaine-Seeking, but not Food-Seeking, Behavior in Rats. Neuropsychopharmacology, 40(10), 2425-2433. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/25837282. doi:10.1038/npp.2015.92
Cunha-Oliveira, T., Rego, A. C., Cardoso, S. M., Borges, F., Swerdlow, R. H., Macedo, T., & de Oliveira, C. R. (2006). Mitochondrial dysfunction and caspase activation in rat cortical neurons treated with cocaine or amphetamine. Brain Res, 1089(1), 44-54. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/16638611.
doi:10.1016/j.brainres.2006.03.061
Cunha-Oliveira, T., Silva, L., Silva, A. M., Moreno, A. J., Oliveira, C. R., & Santos, M. S. (2013). Mitochondrial complex I dysfunction induced by cocaine and cocaine plus morphine in brain and liver mitochondria. Toxicol Lett, 219(3), 298-306. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/23542814. doi:10.1016/j.toxlet.2013.03.025
Dahl, T. B., Holm, S., Aukrust, P., & Halvorsen, B. (2012). Visfatin/NAMPT: a multifaceted molecule with diverse roles in physiology and pathophysiology. Annu Rev Nutr, 32, 229-243. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/22462624.
doi:10.1146/annurev-nutr-071811-150746
David, K. K., Andrabi, S. A., Dawson, T. M., & Dawson, V. L. (2009). Parthanatos, a messenger of death. Front Biosci (Landmark Ed), 14, 1116-1128. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/19273119.
de Oliveira, M. R., & Jardim, F. R. (2016). Cocaine and mitochondria-related signaling in the brain: A mechanistic view and future directions. Neurochem Int, 92, 58-66. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/26707813.
de Souza, M. F., Goncales, T. A., Steinmetz, A., Moura, D. J., Saffi, J., Gomez, R., & Barros, H. M. (2014). Cocaine induces DNA damage in distinct brain areas of female rats under different hormonal conditions. Clin Exp Pharmacol Physiol, 41(4), 265-269. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/24552452.
doi:10.1111/1440-1681.12218
Dolle, C., Rack, J. G., & Ziegler, M. (2013). NAD and ADP-ribose metabolism in mitochondria. FEBS J, 280(15), 3530-3541. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/23617329. doi:10.1111/febs.12304
Fatokun, A. A., Dawson, V. L., & Dawson, T. M. (2014). Parthanatos: mitochondrial-linked mechanisms and therapeutic opportunities. Br J Pharmacol, 171(8), 2000-2016. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/24684389.
doi:10.1111/bph.12416
Feltenstein, M. W., & See, R. E. (2008). The neurocircuitry of addiction: an overview. Br J Pharmacol, 154(2), 261-274. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/18311189. doi:10.1038/bjp.2008.51
Fischer-Smith, K. D., Houston, A. C., & Rebec, G. V. (2012). Differential effects of cocaine access and withdrawal on glutamate type 1 transporter expression in rat nucleus accumbens core and shell. Neuroscience, 210, 333-339. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/22433294. doi:10.1016/j.neuroscience.2012.02.049
Fricker, R. A., Green, E. L., Jenkins, S. I., & Griffin, S. M. (2018). The Influence of
Nicotinamide on Health and Disease in the Central Nervous System. Int J Tryptophan Res, 11, 1178646918776658. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/29844677. doi:10.1177/1178646918776658
Fuchs, R. A., Eaddy, J. L., Su, Z. I., & Bell, G. H. (2007). Interactions of the basolateral amygdala with the dorsal hippocampus and dorsomedial prefrontal cortex regulate drug context-induced reinstatement of cocaine-seeking in rats. Eur J Neurosci, 26(2), 487-498. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/17650119.
doi:10.1111/j.1460-9568.2007.05674.x
Gaignard, P., Savouroux, S., Liere, P., Pianos, A., Therond, P., Schumacher, M., . . . Guennoun, R. (2015). Effect of Sex Differences on Brain Mitochondrial Function and Its
Green, K. N., Steffan, J. S., Martinez-Coria, H., Sun, X., Schreiber, S. S., Thompson, L. M., & LaFerla, F. M. (2008). Nicotinamide restores cognition in Alzheimer's disease transgenic mice via a mechanism involving sirtuin inhibition and selective reduction of Thr231-phosphotau. J Neurosci, 28(45), 11500-11510. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/18987186. doi:10.1523/JNEUROSCI.3203-08.2008
Grimm, J. W., & See, R. E. (2000). Dissociation of primary and secondary reward-relevant limbic nuclei in an animal model of relapse. Neuropsychopharmacology, 22(5), 473-479. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/10731622.
doi:10.1016/S0893-133X(99)00157-8
Gupte, R., Liu, Z., & Kraus, W. L. (2017). PARPs and ADP-ribosylation: recent advances linking molecular functions to biological outcomes. Genes Dev, 31(2), 101-126. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/28202539.
doi:10.1101/gad.291518.116
Guzyk, M. M., Tykhomyrov, A. A., Nedzvetsky, V. S., Prischepa, I. V., Grinenko, T. V., Yanitska, L. V., & Kuchmerovska, T. M. (2016). Poly(ADP-Ribose) Polymerase-1 (PARP-1) Inhibitors Reduce Reactive Gliosis and Improve Angiostatin Levels in Retina of Diabetic Rats. Neurochem Res, 41(10), 2526-2537. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/27255598. doi:10.1007/s11064-016-1964-3
Hagberg, H., Wilson, M. A., Matsushita, H., Zhu, C., Lange, M., Gustavsson, M., . . . Johnston, M. V. (2004). PARP-1 gene disruption in mice preferentially protects males from perinatal brain injury. J Neurochem, 90(5), 1068-1075. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/15312162. doi:10.1111/j.1471-4159.2004.02547.x
Hollis, F., van der Kooij, M. A., Zanoletti, O., Lozano, L., Canto, C., & Sandi, C. (2015). Mitochondrial function in the brain links anxiety with social subordination. Proc Natl Acad Sci U S A, 112(50), 15486-15491. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/26621716. doi:10.1073/pnas.1512653112
Ide, T., Tsutsui, H., Ohashi, N., Hayashidani, S., Suematsu, N., Tsuchihashi, M., . . . Takeshita, A. (2002). Greater oxidative stress in healthy young men compared with premenopausal women. Arterioscler Thromb Vasc Biol, 22(3), 438-442. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/11884287.
20(4), 663-675. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/24975938. doi:10.1111/adb.12159
Javle, M., & Curtin, N. J. (2011). The role of PARP in DNA repair and its therapeutic exploitation. Br J Cancer, 105(8), 1114-1122. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/21989215. doi:10.1038/bjc.2011.382
Jiang, H. Y., Yang, Y., Zhang, Y. Y., Xie, Z., Zhao, X. Y., Sun, Y., & Kong, W. J. (2018). The dual role of poly(ADP-ribose) polymerase-1 in modulating parthanatos and autophagy under oxidative stress in rat cochlear marginal cells of the stria vascularis. Redox Biol, 14, 361-370. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/29049980. doi:10.1016/j.redox.2017.10.002
Kander, M. C., Cui, Y., & Liu, Z. (2017). Gender difference in oxidative stress: a new look at the mechanisms for cardiovascular diseases. J Cell Mol Med, 21(5), 1024-1032. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/27957792. doi:10.1111/jcmm.13038
Kim, R., Sepulveda-Orengo, M. T., Healey, K. L., Williams, E. A., & Reissner, K. J. (2018). Regulation of glutamate transporter 1 (GLT-1) gene expression by cocaine self-administration and withdrawal. Neuropharmacology, 128, 1-10. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/28919080.
doi:10.1016/j.neuropharm.2017.09.019
Kirkland, J. B. (2009). Niacin status, NAD distribution and ADP-ribose metabolism. Curr Pharm Des, 15(1), 3-11. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/19149597.
Knackstedt, L. A., Melendez, R. I., & Kalivas, P. W. (2010). Ceftriaxone restores glutamate homeostasis and prevents relapse to cocaine seeking. Biol Psychiatry, 67(1), 81-84. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/19717140.
doi:10.1016/j.biopsych.2009.07.018
Kong, J., Du, C., Jiang, L., Jiang, W., Deng, P., Shao, X., . . . Cen, X. (2018). Nicotinamide phosphoribosyltransferase regulates cocaine reward through Sirtuin 1. Exp Neurol, 307, 52-61. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/29753648.
doi:10.1016/j.expneurol.2018.05.010
Lax, E., Friedman, A., Massart, R., Barnea, R., Abraham, L., Cheishvili, D., . . . Yadid, G. (2017). PARP-1 is required for retrieval of cocaine-associated memory by binding to the promoter of a novel gene encoding a putative transposase inhibitor. Mol Psychiatry, 22(4), 570-579. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/27595592. doi:10.1038/mp.2016.119
Liang, M., Yin, X. L., Wang, L. Y., Yin, W. H., Song, N. Y., Shi, H. B., . . . Yin, S. K. (2017). NAD+ Attenuates Bilirubin-Induced Hyperexcitation in the Ventral Cochlear Nucleus by Inhibiting Excitatory Neurotransmission and Neuronal Excitability. Front Cell Neurosci, 11, 21. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/28217084.
doi:10.3389/fncel.2017.00021
Liu, D., Pitta, M., & Mattson, M. P. (2008). Preventing NAD(+) depletion protects neurons against excitotoxicity: bioenergetic effects of mild mitochondrial uncoupling and caloric restriction. Ann N Y Acad Sci, 1147, 275-282. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/19076449. doi:10.1196/annals.1427.028
Logan, R. W., Parekh, P. K., Kaplan, G. N., Becker-Krail, D. D., Williams, W. P., 3rd,
Yamaguchi, S., . . . McClung, C. A. (2018). NAD+ cellular redox and SIRT1 regulate the diurnal rhythms of tyrosine hydroxylase and conditioned cocaine reward. Mol Psychiatry. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/29728703.
doi:10.1038/s41380-018-0061-1
Lonskaya, I., Potaman, V. N., Shlyakhtenko, L. S., Oussatcheva, E. A., Lyubchenko, Y. L., & Soldatenkov, V. A. (2005). Regulation of poly(ADP-ribose) polymerase-1 by DNA structure-specific binding. J Biol Chem, 280(17), 17076-17083. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/15737996. doi:10.1074/jbc.M413483200
McCullough, L. D., Zeng, Z., Blizzard, K. K., Debchoudhury, I., & Hurn, P. D. (2005). Ischemic nitric oxide and poly (ADP-ribose) polymerase-1 in cerebral ischemia: male toxicity, female protection. J Cereb Blood Flow Metab, 25(4), 502-512. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/15689952. doi:10.1038/sj.jcbfm.9600059
McFarland, K., & Kalivas, P. W. (2001). The circuitry mediating cocaine-induced reinstatement of drug-seeking behavior. J Neurosci, 21(21), 8655-8663. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/11606653.
Mukhopadhyay, P., Horvath, B., Rajesh, M., Varga, Z. V., Gariani, K., Ryu, D., . . . Pacher, P. (2017). PARP inhibition protects against alcoholic and non-alcoholic steatohepatitis. J Hepatol, 66(3), 589-600. Retrieved from
Mutasim, D. F. (2003). Autoimmune bullous dermatoses in the elderly: diagnosis and management. Drugs Aging, 20(9), 663-681. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/12831291.
Namazi, M. R. (2004). Nicotinamide: a potential anti-addiction weapon. Medical Hypotheses, 62(5), 844-845. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/15082118. doi:10.1016/j.mehy.2004.01.020
Negi, G., Kumar, A., Kaundal, R. K., Gulati, A., & Sharma, S. S. (2010). Functional and biochemical evidence indicating beneficial effect of Melatonin and Nicotinamide alone and in combination in experimental diabetic neuropathy. Neuropharmacology, 58(3), 585-592. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/20005237. doi:10.1016/j.neuropharm.2009.11.018
Otte, N., Borelli, C., & Korting, H. C. (2005). Nicotinamide - biologic actions of an emerging cosmetic ingredient. Int J Cosmet Sci, 27(5), 255-261. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/18492206. doi:10.1111/j.1467-2494.2005.00266.x
Peterson, T. C., Anderson, G. D., Kantor, E. D., & Hoane, M. R. (2012). A comparison of the effects of nicotinamide and progesterone on functional recovery of cognitive behavior following cortical contusion injury in the rat. J Neurotrauma, 29(18), 2823-2830. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/23016598.
doi:10.1089/neu.2012.2471
Pomierny-Chamiolo, L., Moniczewski, A., Wydra, K., Suder, A., & Filip, M. (2013). Oxidative stress biomarkers in some rat brain structures and peripheral organs underwent cocaine. Neurotox Res, 23(1), 92-102. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/22791409. doi:10.1007/s12640-012-9335-6
Rennie, G., Chen, A. C., Dhillon, H., Vardy, J., & Damian, D. L. (2015). Nicotinamide and neurocognitive function. Nutr Neurosci, 18(5), 193-200. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/24559077. doi:10.1179/1476830514Y.0000000112
Requardt, R. P., Hirrlinger, P. G., Wilhelm, F., Winkler, U., Besser, S., & Hirrlinger, J. (2012). Ca(2)(+) signals of astrocytes are modulated by the NAD(+)/NADH redox state. J Neurochem, 120(6), 1014-1025. Retrieved from
Requardt, R. P., Wilhelm, F., Rillich, J., Winkler, U., & Hirrlinger, J. (2010). The biphasic NAD(P)H fluorescence response of astrocytes to dopamine reflects the metabolic actions of oxidative phosphorylation and glycolysis. J Neurochem, 115(2), 483-492. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/20698931. doi:10.1111/j.1471-4159.2010.06940.x
Riklis, E., Kol, R., & Marko, R. (1990). Trends and developments in radioprotection: the effect of nicotinamide on DNA repair. Int J Radiat Biol, 57(4), 699-708. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/1969902.
Sakakibara, Y., Mitha, A. P., Ogilvy, C. S., & Maynard, K. I. (2000). Post-treatment with nicotinamide (vitamin B(3)) reduces the infarct volume following permanent focal cerebral ischemia in female Sprague-Dawley and Wistar rats. Neurosci Lett, 281(2-3), 111-114. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/10704755.
Schieber, M., & Chandel, N. S. (2014). ROS function in redox signaling and oxidative stress. Curr Biol, 24(10), R453-462. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/24845678. doi:10.1016/j.cub.2014.03.034
Scobie, K. N., Damez-Werno, D., Sun, H., Shao, N., Gancarz, A., Panganiban, C. H., . . . Nestler, E. J. (2014). Essential role of poly(ADP-ribosyl)ation in cocaine action. Proc Natl Acad Sci U S A, 111(5), 2005-2010. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/24449909. doi:10.1073/pnas.1319703111
Scofield, M. D., Li, H., Siemsen, B. M., Healey, K. L., Tran, P. K., Woronoff, N., . . . Reissner, K. J. (2016). Cocaine Self-Administration and Extinction Leads to Reduced Glial Fibrillary Acidic Protein Expression and Morphometric Features of Astrocytes in the Nucleus Accumbens Core. Biol Psychiatry, 80(3), 207-215. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/26946381. doi:10.1016/j.biopsych.2015.12.022
See, R. E. (2005). Neural substrates of cocaine-cue associations that trigger relapse. Eur J Pharmacol, 526(1-3), 140-146. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/16253228. doi:10.1016/j.ejphar.2005.09.034
Siegel, C. S., & McCullough, L. D. (2013). NAD+ and nicotinamide: sex differences in cerebral ischemia. Neuroscience, 237, 223-231. Retrieved from
Song, S. B., Jang, S. Y., Kang, H. T., Wei, B., Jeoun, U. W., Yoon, G. S., & Hwang, E. S. (2017). Modulation of Mitochondrial Membrane Potential and ROS Generation by Nicotinamide in a Manner Independent of SIRT1 and Mitophagy. Mol Cells, 40(7), 503-514. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/28736426.
doi:10.14348/molcells.2017.0081
Steinmetz, A., Steffens, L., Moras, A. M., Prezzi, F., Braganhol, E., Saffi, J., . . . Moura, D. J. (2018). In vitro model to study cocaine and its contaminants. Chem Biol Interact, 285, 1-7. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/29475069.
doi:10.1016/j.cbi.2018.01.017
Stratford, M. R., & Dennis, M. F. (1994). Pharmacokinetics and biochemistry studies on nicotinamide in the mouse. Cancer Chemother Pharmacol, 34(5), 399-404. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/8070006.
Suh, S. W., Aoyama, K., Alano, C. C., Anderson, C. M., Hamby, A. M., & Swanson, R. A. (2007). Zinc inhibits astrocyte glutamate uptake by activation of poly(ADP-ribose) polymerase-1. Mol Med, 13(7-8), 344-349. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/17728843. doi:10.2119/2007-00043.Suh
Surjana, D., & Damian, D. L. (2011). Nicotinamide in dermatology and photoprotection. Skinmed, 9(6), 360-365. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/22256624.
Surjana, D., Halliday, G. M., Martin, A. J., Moloney, F. J., & Damian, D. L. (2012). Oral nicotinamide reduces actinic keratoses in phase II double-blinded randomized controlled trials. J Invest Dermatol, 132(5), 1497-1500. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/22297641. doi:10.1038/jid.2011.459
Tamsett, T. J., Picchione, K. E., & Bhattacharjee, A. (2009). NAD+ activates KNa channels in dorsal root ganglion neurons. J Neurosci, 29(16), 5127-5134. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/19386908. doi:10.1523/JNEUROSCI.0859-09.2009
Turunc Bayrakdar, E., Uyanikgil, Y., Kanit, L., Koylu, E., & Yalcin, A. (2014). Nicotinamide treatment reduces the levels of oxidative stress, apoptosis, and PARP-1 activity in Abeta(1-42)-induced rat model of Alzheimer's disease. Free Radic Res, 48(2), 146-158. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/24151909.
doi:10.3109/10715762.2013.857018
Virag, L., & Szabo, C. (2002). The therapeutic potential of poly(ADP-ribose) polymerase inhibitors. Pharmacol Rev, 54(3), 375-429. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/12223530.
Wang, C., Zhang, Y., Ding, J., Zhao, Z., Qian, C., Luan, Y., & Teng, G. J. (2017). Nicotinamide Administration Improves Remyelination after Stroke. Neural Plast, 2017, 7019803. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/28656112.
doi:10.1155/2017/7019803
Wang, P., Xu, T. Y., Guan, Y. F., Tian, W. W., Viollet, B., Rui, Y. C., . . . Miao, C. Y. (2011). Nicotinamide phosphoribosyltransferase protects against ischemic stroke through SIRT1-dependent adenosine monophosphate-activated kinase pathway. Ann Neurol, 69(2), 360-374. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/21246601.
doi:10.1002/ana.22236
Xu, J., Xu, S. Q., Liang, J., Lu, Y., Luo, J. H., & Jin, J. H. (2012). [Protective effect of
nicotinamide in a mouse Parkinson's disease model]. Zhejiang Da Xue Xue Bao Yi Xue Ban, 41(2), 146-152. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/22499510.
Yang, Y., Yao, H., Lu, Y., Wang, C., & Buch, S. (2010). Cocaine potentiates astrocyte toxicity mediated by human immunodeficiency virus (HIV-1) protein gp120. PLoS One, 5(10), e13427. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/20976166. doi:10.1371/journal.pone.0013427
Yao, H., Allen, J. E., Zhu, X., Callen, S., & Buch, S. (2009). Cocaine and human
immunodeficiency virus type 1 gp120 mediate neurotoxicity through overlapping signaling pathways. J Neurovirol, 15(2), 164-175. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/19319745. doi:10.1080/13550280902755375
Ying, W. (2007). NAD+ and NADH in brain functions, brain diseases and brain aging. Front Biosci, 12, 1863-1888. Retrieved from
Ying, W., Garnier, P., & Swanson, R. A. (2003). NAD+ repletion prevents PARP-1-induced glycolytic blockade and cell death in cultured mouse astrocytes. Biochem Biophys Res Commun, 308(4), 809-813. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/12927790.
Zenclussen, M. L., Casalis, P. A., Jensen, F., Woidacki, K., & Zenclussen, A. C. (2014). Hormonal Fluctuations during the Estrous Cycle Modulate Heme Oxygenase-1 Expression in the Uterus. Front Endocrinol (Lausanne), 5, 32. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/24659985. doi:10.3389/fendo.2014.00032
Zeng, J., Yang, G. Y., Ying, W., Kelly, M., Hirai, K., James, T. L., . . . Litt, L. (2007). Pyruvate improves recovery after PARP-1-associated energy failure induced by oxidative stress in neonatal rat cerebrocortical slices. J Cereb Blood Flow Metab, 27(2), 304-315. Retrieved from https://www.ncbi.nlm.nih.gov/pubmed/16736046.