Venkat A. et al., J. Sci. Res. Phar. 2013, 2(1), 1-7
J
ournal of
S
cientific
R
esearch in
P
harmacy
Research Article
Available online thr oug h
ISSN: 2277-9469
www.jsrponline.com
Protective role of Minocycline through zinc chelation in LPS induced Alzheimer’s model
Venkat A1*, Chaitanya P2, Ravinder P1, Dr.Shilendra3
*1Sri Krupa Institute Of Pharmaceutical Sciences, Medak Dist, India. 2Lalitha College of Pharmacy, RR Dist, India. 3Mediciti Institute of Medical Sciences, India.
Received on: 03-02-2013; Revised and Accepted on: 18-02-2013
ABSTRACT
Objective: The present study aims to investigate the neuroprotective effect of minocycline on the neuronal apoptosis by altering intrace llular zinc levels through its property of chelation.
Methods: Differentiated PC-12 cells were pretreated with varying concentration of minocycline (5μM,10 μM) for 2hr prior to treatment with 4μg/ml LPS (LD50) and incubated for 48hrs to assess PC-12 cell viability and oxidative stress, and apoptosis mediated PC-12 cell death mechanisms. Cell viability was analyzed by MTT assay. ROS/NO production was assessed by using Greiss reagent. Apoptosis mediated cell death mechanism was es timated by detecting 12-LOX, P38, Bad expression by western blot. Caspase-3 activity was measured by colorimetric assay. Flouzin-3 probe was used for quantification of zinc concentration.
Results: Our studies showed that the minocycline type of antibiotic can suppress the activation of 12-LOX by forming chelation with zinc. It also suppresses the activation of NO, P38, 12-LOX leads in a dose dependant manner, respectively. Cell apoptosis triggered by LPS and characterized with caspase -3 activity, pro-apoptoti c gene Bad also significantly inhibited by this drug. It also significantly decreased the number of apoptotic cells compared to control when assessed by immune fluorescence staining.
Conclusion: The results indicate that, minocycline showed a significant protective effect in a dose -dependent manner. Previous research works confirmed the therapeutic role of minocycline by improving cognitve performance in Alzheimer’s disease patents. This research work emphasize an additional protective role of minocycline, in protecting neuronal apoptosis in Alzheimer’s disease model.
Key words: Alzheimer’s disease, chelation, minocycline, zinc.
INTRODUCTION
The clinical picture of dementia as a deteriorate in cognition in the past years has been recognized for centuries but understood in its current form only from past 20 years. Alzheimer’s disease (AD) is the most common form of dementia which accounts to about 55-56% of all the cases. The prevalence of dementia is below 1% in individuals aged 60-65 years. But the prevalence is between 23-34% in the western world who aged 85 years or older. Several studies have demonstrated the involvement of reactive oxygen species (ROS) in beta amyloid induced neurotoxicity which has been shown to induce neuronal apoptosis [1].In
recent years research has clearly pointed the importance of oxidative imbalance in AD. There are some fundamental insights that have indicated the importance of oxidative stress and free radical damage in the pathogenesis of AD [2]. Previous studies conformed that oxidative
stress induced neuronal apoptosis is mediated by free Zinc ions activity in which the underlying mechanism was found that Zinc itself can induces the activation of 12 LOX, which suggests that a neuronal apoptosis pathway is triggered by Peroxynitrate [3].In this study we
aimed to block the 12 LOX activity by using a drug which can preferable to form complexation (chelation) with intracellular free Zinc ions by maintaining its homeostasis [4]. The whole phenomenon is assumed to
place by considering Peroxynitrate is one of the major causative for the generation of ROS in oxidative stress condition [5].In this research study
we tried to incorporate an additional mechanism of Minocycline to prevent neuronal apoptosis in oxidative stress condition.
MATERIALS AND METHODS
Bacterial lipo polysaccharide (LPS) E.coli serotype (055:B5), Catalog-H81565 (Cayman chemicals, USA), nerve growth factor (NGF) (Sigma-Aldrich, St. Louis, MI, USA). PC-12 rat pheocromacytoma cell line, F-12k culture media, HS, FBS (American Type Culture Collection, USA),
*Corresponding author:
Venkat ASri Krupa Institute Of Pharmaceutical Sciences, Medak Dist, India.
*E-Mail: [email protected]
ELISA kit for Caspase-3 (Invitrogen, USA), Fluozin-3 (Zinc florescent indicator, Sigma-Aldrich, St. Louis, MI, USA), 12-LOX western blot kit (assay designs, MI, USA), HRP conjugated goat anti rabbit antibodies (Cayman chemicals, USA), western blot kit for P38 activation (Biolegend,
USA) and all other reagents from Sigma (St. Louis, M I, USA).
Cell culture and treatment:
PC-12 cells were cultured in a prepared F-12k culture medium. The cells were seeded as 4X104 viable cells/ cm2 in 20 cm2
culture dishes in approximately 10-12 ml of media. The cells were grown in a constant supply of 95% air and 5% CO2 at 37o C and the medium
was changed for every 2 days. Sub culturing was followed for cryopreservation, when the cells became 80-90% confluent. The freezing medium in this cryopreservation was F-12k culture medium and DMSO.
Differentiation of PC-12 cells:
About 60-70% of confluent cells were incubated with low serum media (1% of HS and Penicillin) containing 100ng/ml of 2.5s nerve growth factor for about 12-16 days for complete neurite extension. Again culture medium was changed for every 2 alternative days. Further experiments were performed and evaluated using these differentiated PC-12 cells.
Toxin and drug treatment:
In order to determine the LD50 for LPS, the attached cells
were pre-treated with a concentration of 10µM Minocycline for 2 hrs before adding LPS at a concentration of 4µg/ml. The cells were incubated for 48 and 72 hrs respectively. The protective effect of minocycline was used on LPS induced differentiated PC-12 cells were examined by using the trypan blue exclusion test and the MTT assay.
Cell viability test (MTT assay):
for 3hrs showed a dose dependent increase in the cell viability of LPC treated cells.
NO assay:
NO plays a major role in neuronal cell death by inducing mitochondrial dysfunction and apoptosis. Hence we determine the protective effect of Minocycline on neuronal apoptosis induced by excessive free radical production [15]. Concentration of NO in the culture
supernatants were determined as nitrite which is a major stable oxidation product of NO, using the Griess reagent as descri bed previously. 50µl of conditioned medium was mixed with equal volume of Griess reagent and incubated at room temperature for 10min. The nitrite levels were determined colorimetrically at 550nm using microplate spectrophotometer [20].
Assay of 12-LOX activity:
12-Lipoxygenase activity was measured by quantifying the major metabolite product of 12-LOX, which is 12 hydroxy eicosatetraenoic acid (12 HETE) by using ELISA kit. The treated neurons were taken then the cultures were washed with HBSS containing 0.1mg/ml of BSA and 20µM of arachidonic acid was added to the culture. The medium was collected, extracted and assayed for 12-LOX activity.
The concentration of 12-HETE in each sample was normalized to the protein concentration of the sample. The same protein concentration was also estimated by western blot by using 12 HETE specific HRP conjugated secondary antibody.
Fluorescence assay for intracellular free Zinc ion:
Intracellular zinc ion concentration was determined by Fluozin-3 indicator. Briefly cells were detached and washed with DPBS without calcium and magnesium at 37OC, then 0.3µM of Fluozin-3
acetoxymethyl ester was added, then the cells were washed again to remove any residual fluorescent probe and incubated at 37oC.
Fluorescence was measured at 25oC at 492nm excitation and 517nm as
emission in a spectrofluorimeter [6, 21].
The concentration of zinc ion was calculated with following formula
[Zn+2]= Kd [F-Fmin]/ [Fmax-F]
F=Fluorescence intensity, Kd=Dissociation constant, Fmax= Fluorescence
obtained after addition of zinc selective ionophore, Fmin=Fluorescence
obtained after addition of selective zinc chelator [7].
Effect of minocycline on pro-apoptotic (P38) and anti-apoptotic
(BCL-2) protein expression:
Apoptotic research primarily based on pro-apoptotic and anti-apoptotic gene expression. The balance between those two family members is very important in regulation of apoptosis. To strengthen this, we investigated the effect of minocycline on the modulation of pro and anti-apoptotic genes in LPS induced PC-12 cells. To determine the cell viability and apoptosis after 48hrs of treatment with LPS and pre-treatment with drug. We carried out western blot analysis by using HRP conjugated goat anti-rabbit antibodies as secondary antibody. The expression of two proteins in this experiment were depicted in Fig. 5 & 7.
Effect of Minocycline on Caspase-3 activity:
Caspases are cysteine dependent enzymes that plays a crucial role in the induction, amplification and transduction of intracellular apoptotic pathway. Caspase-3 mainly acts on several substances like Poly ADP ribose polymerase (PARP) and induces apoptosis. To further confirm the protective effect of Minocycline in apoptosis, we quantified
the caspase-3 activity by colorimetric assay. A significant decrease in the activity of caspase-3 at absorbance 405 nm, indicated the reversal of apoptosis.
Morphological evaluation of apoptotic cells:
The evaluation of apoptosis was done by immune fluorescence assay in which the apoptotic cells were successfully labeled with fluorescent probe (PSVwav480) and observed under fluorescent microscope using FITC filter set.
RESULTS
The following results were obtained from the present research study.
1. Lipo polysaccharide induced apoptosis through intracellular Zinc accumulation:
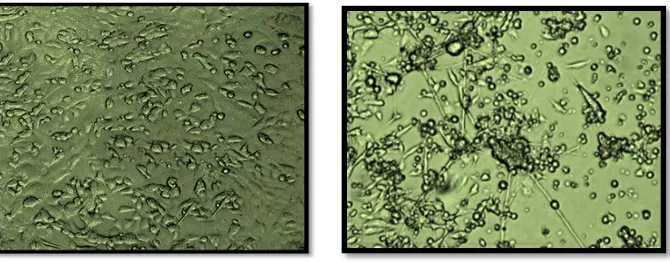
Previous studies had confirmed that some physiological conditions may accumulate the Zinc intracellular concentration and leads to apoptosis. By using fluorescent Zinc indicator, Fluozin-3, we have an evidence to confirm that physiological conditions like oxidative stress increases intracellular free Zinc concentrations leads to neuronal apoptosis. In our study the percentage of viable cells was confirmed by MTT assay in which the control cells had survivability of 100 +5%. The toxin (LPS) treated cells have average percentage viability for 72 hrs of toxin treated cells are 46+5% as depicted in the Fig. 1& 2.
2. Involvement of 12-LOX activation in LPS induced apoptosis:
Earlier research studies confirmed that 12-LOX activation in apoptosis through caspase pathway. Our study confirmed the relationship between high levels of Zinc accumulation and increased 12 -LOX activity, which leads to neuronal apoptosis. The western blot for 12-LOX activity was clearly confirming the high activity in apoptosis induced cells. Whereas decreased activity was achieved with concentration dependent of drug treatment. The statistical measures were depicted in the Fig. 4.
3. Zinc induced apoptosis mediated by activation of 12-LOX:
Present research confirmed that free intracellular Zinc induces activation of 12-LOX which further increases the activation of caspase-3, this caspase involves in caspase dependent apoptotic pathway.[8] Western blot for caspase-3 had shown that Zinc induced
apoptosis is mediated by activation of caspase-3, which is previously activated by 12-LOX. The statisti cal results were shown in the Fig. 4.
4. Relation between P38 and 12-LOX:
Our research work established an evidence that high levels of 12-LOX activation increases P38 (pro-apoptotic) activation which
involves in the caspase (casepase-3 activity can be seen in Fig. 5
dependent apoptosis. Results for western blot for P38 showed the
interdependent relation between P38 expression and 12-LOX activity [23].
5. Protective action of minocycline on LPS induced neurotoxicity:
In our study minocycline antibiotic which readily forms a complex (chelation) with free divalent Zinc atoms. We proved that formation of zinc complex with minocycline delays neuronal apoptosis in oxidative stress like Alzheimer’s disease.
The selective zinc fluorescent probes confirmed the increased survivability of drug treated cells [25]. The decreased fluorescence
intensity is directly proportional to the decreased levels of free zinc concentrations, which represents the protective action against apoptosis
[9]. Fig. 1
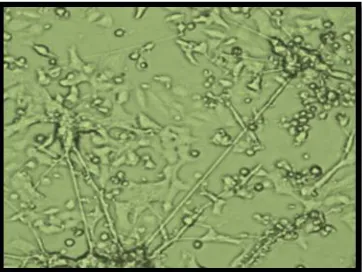
Fig. 1C: LPS(4µg/ml)treated(apoptosis)induced pc-12 cells Fig. 1D: Minocycline10 µg/ml pre-treated pc-12 cells
Fig. 1E: fluorescent probes denoting the free zinc levels Fig. 1F: decreased levels free zinc ions (LPS treated cells) (pre-treated cells with minocycline cells)
2) MTT assay:
Fig. 2: Effect of Minocyclin on cell viability: Cell viability was assessed by the MTT assay. Cells were treated with 0 (control), LPS (LPS 4µg/ml alone),or minocycline (only) for 24, 48 and 72 h. Pretreatment consisted of minocycline at different concentrations 5 µM,10 µM) for 2 h prior to stimulation with LPS for 24, 48 and 72 h. For statistical evaluations two -way ANOVA analysis, followed by Tukey’s range test was performed. (#p < 0.05) Compared with control; (*p < 0.05) compared with LPS treated cells only. Data are expressed as percentage viable cells compared with control cells.
3) NO levels:
Table No. 3
Control LPS (4μg/ml) Minocycline(5μg/ml) Minocycline(10μg/ml)
Fig. 3: Effect of Minocyclinon NO production in LPS induced PC 12 cells Effects: For statistical evaluations two-way ANOVA analysis, followed by tukey’s test was performed - statistically significant difference (#p<0.05) compared with control, statistically significant difference (*p<0.05) compared with 4 µg/ml LPS, and statistically significant difference (∆p<0.05) compared with individual drug treatment.
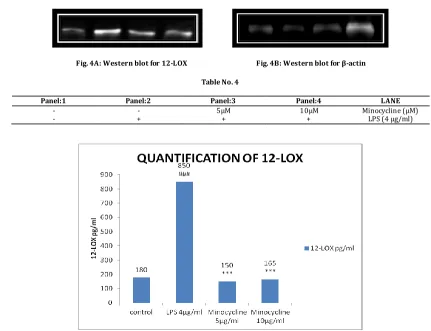
4) 12-LOX:
Fig. 4A: Western blot for 12-LOX Fig. 4B: Western blot for β-actin
Table No. 4
Panel:1 Panel:2 Panel:3 Panel:4 LANE
- - 5μM 10μM Minocycline (μM) - + + + LPS (4 μg/ml)
Fig. 4A: Western blot showing the expression of 12- LOX and β-actin in PC12 cells treated with Minocycline for 48 hours. [4B] Quantitative data for the expression of 12-LOX in Minocycline treated cells. For statistical evaluations two-way ANOVA analysis, followed by tukey’s test was performed - statistically significant difference (#p<0.05) compared with control, and statistically significant difference (*p<0.05) compared with 4 µg/ml LPS.
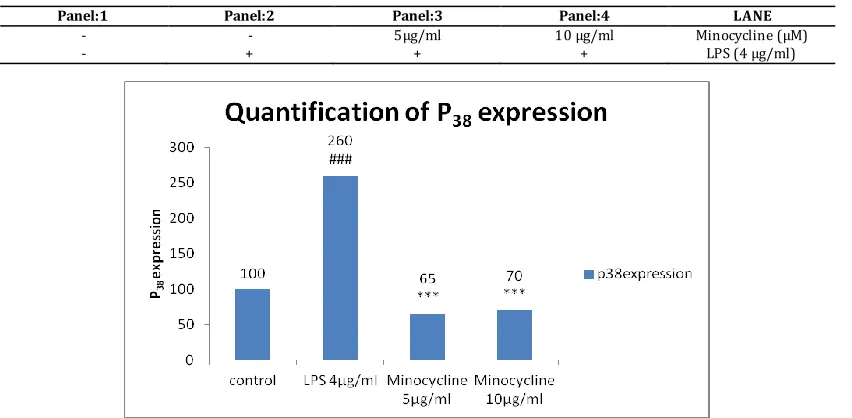
5) P38:
Table No. 5A
Panel:1 Panel:2 Panel:3 Panel:4 LANE
- - 5μg/ml 10 μg/ml Minocycline (μM) - + + + LPS (4 μg/ml)
Fig. 5A: Western blot showing the expression of P38 and β-actin in PC12 cells treated with Minocyclin for 48 hours. 5B) Quantitative data for
the expression of P38 in Minocyclin treated cells. For statisti cal evaluations two-way ANOVA analysis, followed by tukey’s test was performed -
statisti cally significant difference (#p<0.05) compared with control, and statistically significant difference (*p<0.05) compared with 4 µg/ml LPS.
6) Caspase-3:
Table No. 6A
Control LPS (4μg/ml) Minocycline(5μg/ml) Minocycline(10μg/ml)
0.1 1.35 0.1*** 0.1***
Fig. 6A: Effect of Minocyclin on caspase-3 activation in LPS induced PC 12 cells. For statistical evaluations two-way ANOVA analysis, followed by tukey’s test was performed - statistically significant difference (#p<0.05) compared with control, statistically significant difference (*p<0.05) compared with 4 µg/ml LPS, and , statistically significant difference (∆p<0.05) compared with Minocyclin treatment.
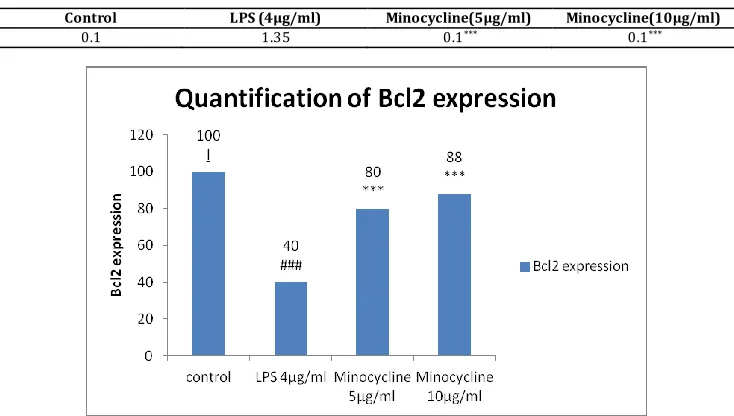
7) Bcl2:
Fig. 7A: Western blot for Bcl-2 Fig. 7B: Western blot for β-actin
Table No. 7A
Panel:1 Panel:2 Panel:3 Panel:4 LANE
Table No. 7B
Control LPS (4μg/ml) Minocycline(5μg/ml) Minocycline(10μg/ml)
0.1 1.35 0.1*** 0.1***
Fig. 7A: Western blot showing the expression of Bcl-2 and β-actin in PC12 cells treated with Minocyclin for 48 hours. [B] Quantitative data for the expression of Bcl-2 in Minocyclin treated cells. For statistical evaluations two-way ANOVA analysis, followed by tukey’s test was performed - statisti cally significant difference (#p<0.05) compared with control, and statistically significant difference (*p<0.05 ) compared with 4 µg/ml LPS.
DISCUSSION
Zinc translocati on from presynaptic nerve terminals to postsynaptic neurons has been considered the critical step leads to the accumulation of intracellular free zinc and subsequent neuronal death. Liberation of zinc from intracellular stores upon oxidative & nitrative stimulation contributes significantly to the toxicity of this metal to neurons [14].
The cell death signaling pathways are triggered by zinc. The below sections will discuss, How the activation of 12-LOX, Mitogen-Activated Protein Kinase (MAPK) contribute to the toxicity of liberated zinc to neurons [10].
Oxidative stress and intracellular zinc liberation:
Zinc is an important structural and functional component in many cellular proteins and enzymes. The concentrations of zinc are believed to be in picomolar to low nanomolar range. The increase in Zn+2
beyond these nanomolar levels is generally believed to contribute in rapid cellular demise [16].
Mechanism of zinc toxicity to neurons:
Previous research works had confirmed that, the intracellular zinc release is triggered by oxidative and nitrative stressors is the necessary first step to initiate cell death signaling pathways [17].
P38MAPK, DTDP, a cell permeant thiol oxidant has been found
to liberate zinc from intracellular stores and induce neuronal apoptosis via activation of P38 MAPK [21]. The activation of P38 MAPK is followed by
potassium efflux and the subsequent activation of caspases. The efflux of potassium ions is mediated by the direct phosphorylation of Kv2.1 encoded K+ channels by P38, leading to a SNARE-dependent insertion of
the channel and a dramatic increase in K+ currents [12].The toxicity is
mediated by the intracellular formation of peroxynitrate and subsequent release of zinc from intracellular stores. Thus, P38MAPK activation is
downstream of mitochondria dysfunction and Reactive Oxygen species (ROS) generation followed by zinc liberation [22].
Previous studies using peroxynitrite as an oxidizing agent also indicated that 12-lipoxygenase (12-LOX) activation contributes in mitochondria independent reactive oxygen species (ROS) generation, also occurs in upstream of P38MAPK activati on.
Several studies were confirmed a detrimental role for ERK1/2 signaling during oxidative neuronal injury. It has been formed that methyl-isothiazolinone (MIT), can trigger intracellular zinc release likely via the oxidation of cellular thiols.
Interestingly, Unlike the other oxidizing and nitrating agents mentioned above, neuronal cell death induced by MIT is mediated by activations of ERK1/2, and not P38 MAPK [11].Oxidative neuronal cell
death model in this involves downstream of zinc release and P38 MAPK
activation.
The Fig. 1 flow chart gives an idea of sequential events in oxidative stress pathway which leads to apoptosis.
Activation of 12- LOX:
Mitochondria are generally believed to be the major source of reactive oxygen species (ROS) generation induced by loss of cellular zinc homeostasis. However, recent studies also provide evidence of mitochondria independent reactive oxygen species (ROS) production, such as those generated from the activation of NADPH oxidase and 12 -LOX. 12-LOX is the primary LOX expressed in the brain and belongs to a family of Arachidonic acid metabolizing enzymes, which also include 5-LOX, cyclooxygenase and epoxygenase [19].
In previous studies it confirmed that glutamate induced Glutathione depletion causes 12-LOX activation, which leads to reactive oxygen species (ROS) generation and neuron cells apoptosis in primary culture of immature neurons. Activation of 12-LOX has also been found in MIT and peroxynitrate induced toxicity in neuron cells culture. Teracycline’s type of antibioti cs (Minocycline) works as a Zn+2chelator,
may blocks the membrane translocation and the activity of 12-LOX, strongly suggest that intracellular zinc release may causes activation of this enzyme [13].
Free radical generation and mitochondria dysfunction are thought to be the critical effectors of zinc mediated neuron toxicity. Zinc released from presynaptic terminals or from intracellular zinc binding proteins may impair the mitochondrial membrane potential and causes free radical generation and apoptic neural injury [24].
By these results we found that 12-LOX inhibition completely blocks reactive oxygen species (ROS) generation induced by zinc. These results were suggest that reactive oxygen species (ROS)primarly be derived from the activation of 12-LOX and, dysfunction of mitochondria might further facilitates the formation of free radicals.
It is therefore reasonable to speculate that rising levels of zinc during oxidative stress conditions contributes to 12-LOX activation and neuronal apoptosis. Significantly 12-Lox inhibitors and zinc chelating agent drugs like antibiotics (Teracycline’s & Quinoline) are may protective against the oxidative stress conditions and in Alzheimer’s disease.
CONCLUSION
Many research works were shown that, intracellular free zinc is toxic to motor neurons. It will be interesting to determine whether, blockade of 12-LOX activity might also have a therapeutic effect in improving the survival of motor neurons.
Our results suggest that, Zinc homeostasis and the 12-LOX metabolic pathway may play an important role in the pathogenesis of a variety of acute and chronic neurodegenerative diseases in which oxidative stress is involved.
REERENCES:
1. Li MH, Jang JH, Sun B, Surh YJ. Protective effects of oligomers of grape seed polyphenols against beta-amyloid-induced oxidative cell death. Ann. N. Y. Acad. Sci., 2004:1030; 317-29.
2. Smith MA, Rottkamp CA, Nunomura A, Raina AK, Perry G. Oxidative stress in Alzheimer's disease. Biochim. Biophys. Acta,
2000: 1502(1); 139-44.
3. Smith MA, Richey Harris PL, Sayre LM, Beckman JS, Perry G. Widespread peroxynitrite-mediated damage in Alzheimer's disease. J. Neurosci., 1997: 17(8); 2653-7.
4. Lovell MA, Xie C, Markesbery WR. Protection against amyloid beta peptide toxicity by zinc. Brain Res., 1999: 823(1-2); 88-95. 5. Giulian D, Li J, Li X, George J, Rutecki PA. The impact of
microglia-derived cytokines upon gliosis in the CNS. Dev. Neurosci., 1994: 16(3-4); 128-36.
6. Canzoniero LM, Sensi SL, Choi DW. Measurement of intracellular free zinc in living neurons. Neurobiol. Dis., 1997: 4(3-4); 275-9. 7. Fierke CA, Thompson RB. Fluorescence-based biosensing of zinc
using carbonic anhydrase. Biometals., 2001: 14(3-4); 205-22. 8. Frederickson CJ. Neurobiology of zinc and zinc-containing
neurons. Int. Rev. Neurobiol., 1989: 31; 145-238.
9. Gee KR, Zhou ZL, Qian WJ, Kennedy R. Detection and imaging of zinc secretion from pancreatic beta -cells using a new fluorescent zinc indicator. J. Am. Chem. Soc., 2002: 124(5); 776-8.
10. Krezel A, Maret W. Dual nanomolar and picomolar Zn(II) binding properties of metallothionein. J. Am. Chem. Soc., 2007: 129(35); 10911-21.
11. Wullner U, Loschmann PA, Schulz JB, Schmid A, Dringen R, Eblen F, et al. Glutathione depletion potentiates MPTP and MPP+ toxicity in nigral dopaminergic neurones. Neuroreport., 1996: 22;7(4); 921-3.
12. Li Y, Maher P, Schubert D. A role for 12-lipoxygenase in nerve cell death caused by glutathione depletion. Neuron., 1997: 19(2); 453-63.
13. Nagasawa K, Kakuda T, Higashi Y, Fujimoto S. Possible involvement of 12-lipoxygenase activation in glucose-deprivation/reload-treated neurons. Neurosci. Lett., 2007: 429(2-3); 120-5.
14. Aizenman E, Stout AK, Hartnett KA, Dineley KE, McLaughlin B, Reynolds IJ. Induction of neuronal apoptosis by thiol oxidation:
putative role of intracellular zinc release. J. Neurochem., 2000: 75(5); 1878-88.
15. Berendji D, Kolb-Bachofen V, Meyer KL, Grapenthin O, Weber H, Wahn V, et al. Nitric oxide mediates intracytoplasmic and intranuclear zinc release. FEBS Lett., 1997: 405(1); 37-41. 16. Frederickson CJ, Koh JY, Bush AI. The neurobiology of zinc in
health and disease. Nat. Rev. Neurosci., 2005: 6(6); 449-62. 17. Kim YH, Kim EY, Gwag BJ, Sohn S, Koh JY. Zinc-induced cortical
neuronal death with features of apoptosis and necrosis: mediation by free radicals. Neuroscience, 1999: 89(1); 175-82. 18. Du S, McLaughlin B, Pal S, Aizenman E. In vitro neurotoxicity of
methylisothiazolinone, a commonly used industrial and household biocide, proceeds via a zinc and extracellular signal-regulated kinase mitogen-activated protein kinase-dependent pathway. J. Neurosci., 2002: 22(17); 7408-16.
19. Kudo I, Murakami M. Phospholipase A2 enzymes. Prostaglandins Other Lipid Mediat; 2002: 68-69, 3-58.
20. Bossy-Wetzel E, Talantova MV, Lee WD, Scholzke MN, Harrop A, Mathews E, et al. Crosstalk between nitric oxide and zinc pathways to neuronal cell death involving mitochondrial dysfunction and p38-activated K+ channels. Neuron., 2004: 41(3); 351-65.
21. Bozym RA, Thompson RB, Stoddard AK, Fierke CA. Measuring picomolar intracellular exchangeable zinc in PC-12 cells using a ratiometric fluorescence biosensor. ACS Chem. Biol., 2006: 1(2); 103-11.
22. Sian J, Dexter DT, Lees AJ, Daniel S, Agid Y, Javoy -Agid F, et al. Alterations in glutathione levels in Parkinson's disease and other neurodegenerative disorders affecting basal ganglia. Ann. Neurol., 1994: 36(3); 348-55.
23. Lebeau A, Terro F, Rostene W, Pelaprat D. Blockade of 12-lipoxygenase expression protects cortical neurons from apoptosis induced by beta-amyloid peptide. Cell Death Differ.,
2004: 11(8); 875-84.
24. Li Y, Maret W. Transient fluctuations of intracellular zinc ions in cell proliferation. Exp. Cell Res., 2009: 315(14); 2463-70. 25. Mistri T, Dolai M, Chakraborty D, Khuda-Bukhsh AR, Das KK, Ali
M. A highly selective and sensitive in vivo fluorosensor for zinc (II) without cytotoxicity. Org. Biomol, Chem., 2012: 10(12); 2380-4.
Conflict of interest: The authors have declared that no conflict of interest exists.