The effect of bacteria, enzymes and inulin on fermentation and aerobic stability of corn silage.
Full text
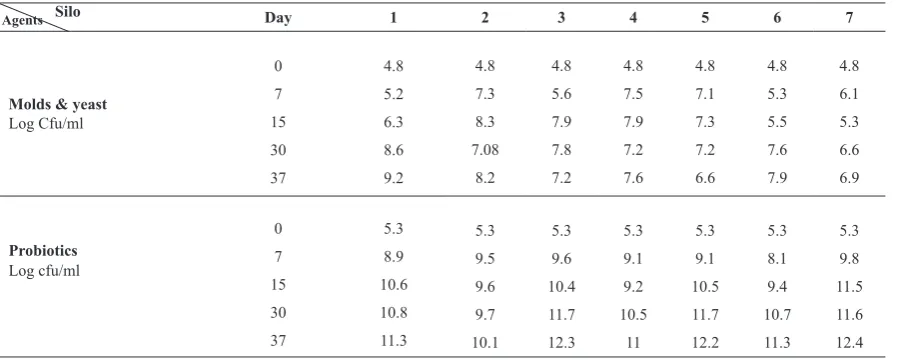
Figure

Related documents
Comparing the figures for the five largest banking groups headed by banche popolari with those for major and large banks of comparable size, the gap widens to
The Java SDK uses the VDFLoader , an abstract class, to load VDF data for a single video in to a VDF object. The VDFLoadParam , consisting of VDF- Loader parameters, specifies
Pyramid or pyramid representation is a type of multi-scale signal representation developed by the computer vision, image processing and signal processing communities, in which
Twenty-five percent of our respondents listed unilateral hearing loss as an indication for BAHA im- plantation, and only 17% routinely offered this treatment to children with
lustrates the time course in S-GOT in hepa- titis contacts with abnormal variability in S-GOT. It is worth emphasizing that, in spite of living in the same households with one or
Motivated by primality and integer factorization, this thesis introduces generalizations of standard binary multiplication to commutative n-ary operations based upon
o Generally, blanket approval has been granted by the State Authority the Land and Mines Department (PTG), subject to the Foreign Ownership Guideline in Johor
The purposes of this study were: (1) To investigate the leadership style of selected construction managers/project managers/superintendents in South Florida using Hersey
